










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.16 n.1 Mendoza ene./jun. 2009
ARTÍCULOS Y NOTAS
Aproximación cualitativa y aspectos funcionales en la ontogenia craneana de Puma concolor (Felidae)
Valentina Segura* y David Flores
Museo Argentino de Ciencias Naturales «Bernardino Rivadavia», Av. Ángel Gallardo 470, CP1405 Ciudad Autónoma de Buenos Aires, Argentina
*[Correspondencia: <vsegura@macn.gov.ar>]
RESUMEN: La variación ontogenética en félidos fue abordada principalmente desde la perspectiva de estimación de edades, en base a secuencia de desgaste y reemplazo dentario. Sin embargo, casi ninguna especie ha sido estudiada aún en un contexto morfológico-funcional en relación al cambio de dieta y comportamiento. En este trabajo analizamos la variación ontogenética de Puma concolor, tanto de las estructuras morfológicas como de los tipos y grado de fusión de las suturas, con el fin de relacionar cambios de forma con cambios de hábitos que ocurren durante el crecimiento. Nuestra muestra incluye especímenes de entre menos de 10 días a más de 10 años de edad. Si bien la mayor parte de las estructuras presentes en adultos están ya manifestadas en estadíos juveniles finales, casi todos los cambios tienden al crecimiento y reforzamiento de éstas, apuntando al fortalecimiento funcional del cráneo, vía el aumento del área de origen e inserción de musculatura temporal, masetérica y pterigoidea, así como el aumento en volumen de la musculatura cervical. La mayoría de las modificaciones suturales se evidencian antes de alcanzar la dentición definitiva y están asociadas al aumento de la resistencia al estrés mecánico, siendo los cambios más frecuentes la adquisición de la sutura de tipo aserrada y su fusión final. La comparación de los patrones evidenciados en P. concolor con félidos relacionados de menor tamaño (e.g. P. yagouaroundi) resultan pendientes, ya que las diferencias probablemente impliquen patrones de crecimiento distintos, producto de exigencias relacionadas con el tamaño de las presas.
ABSTRACT: Qualitative approach and function in the cranial ontogeny of Puma concolor (Felidae). The ontogenetic variation in felids was studied mainly from the perspective of age estimation based on the sequence of tooth replacement and wear. However, almost no species has been still analyzed in a morpho-functional frame, relating changes on diet and behavior. In this work, we analyzed the ontogenetic variation of the skull of Puma concolor, both in its morphological structures, as well as in its suture types and stage of fusion, in order to associate changes of shape with changes of habits that occur during the growth. Our sample includes specimens younger than 10 days, to specimens of approximately 10 years old. Although most of the structures present in adults are already observed in terminal young stages, almost all changes point to the functional strengthening of the skull, via the increased area for the origin and insertion of temporal, masseteric, and pterygoid musculature, as well as the enlargement of cervical muscles. Most of the sutural changes occur before reaching the permanent dentition, and are associated to the improvement of the resistance to mechanical stress, being the commonest change the acquisition of serrated suture and its final fusion. The comparison of the patterns observed in P. concolor with related species of lesser body size (e.g., P. yagouaroundi) are pending, since some differences can be associated to different growth pattern, product of different mechanical demands related to the size of preys.
Palabras clave. Anatomía; Carnivora; Desarrollo.
Key words. Anatomy; Carnivora; Development.
INTRODUCCIÓN
La familia Felidae constituye un diverso grupo de mamíferos con hábitos carnívoros estrictos, comportamiento predador bien desarrollado y tamaño corporal muy diverso, contando con especies de tamaño medio como Leopardus pardalis y otras mayores como Puma concolor o Panthera onca (Gomes de Oliveira, 1993; Lucherini et al., 2004). Durante el crecimiento, ocurren no solamente cambios en la composición alimenticia, sino también en los patrones de comportamiento relacionados con el aprendizaje de métodos de caza (Ewer, 1973; Binder y Van Valkenburgh, 2000). Es entonces esperable que las transformaciones morfológicas sufridas por el neurocráneo y el aparato trófico con relación a la edad, además del reemplazo dentario, tengan profundas implicancias funcionales que acompañen la adquisición de una dieta carnívora y nuevos patrones de comportamiento. Además, la uniformidad morfológica y variabilidad de tamaño que se observa en los félidos (Martin, 1996; Wozencraft, 1996) crean un ambiente especial para la comparación de los patrones de ontogenia craneana entre diferentes especies.
La morfología craneana en los félidos ha sido investigada desde varios enfoques, ya sea con un criterio puramente descriptivo (Kelson, 1946; Ximénez, 1974; Gilbert, 1997; García- Perea, 2002), filogenético (Collier y O'Brien, 1985; Salles, 1992; Johnson y O'Brien, 1997; Mattern y McLennan, 2000), o funcional (Emerson y Radinsky, 1980; Martin, 1980; Biknevicius, 1996; Antón et al., 2004). Sin embargo, las modificaciones estructurales del cráneo durante la ontogenia han sido escasamente estudiadas en un contexto morfológico. Solamente de algunos grupos de félidos, tales como Panthera onca (Stehlik, 1971; Slaughter et al., 1974), P. pardus (Stander, 1997), Leopardus wiedii (Volf, 1972; Fagen y Wiley, 1978; Petersen y Petersen, 1978), Lynx (Crowe, 1975; Johnson et al., 1981; Jackson et al., 1988; García-Perea, 1996), Acinonyx jubatus (Broom, 1949; Caro, 1994), Caracal caracal (Stuart y Stuart, 1985) y Puma concolor (Gay y Best, 1996a; Shaw et al., 2007) se han obtenido datos en un marco morfológico-ontogenético, aunque la mayor parte de ellos superficiales, no siempre considerando caracteres craneanos ni contexto funcional y apuntados a la estimación de edades. No obstante, los patrones de osificación de la mandíbula y su funcionalidad en relación al reemplazo dentario, recibieron cierta atención en P. concolor (ver Biknevicius, 1996; Biknevicius y Leigh, 1997).
Los recientes estudios ontogenéticos cualitativos realizados en mamíferos en sentido funcional y comparativo general del cráneo, están enfocados especialmente en marsupiales didélfidos (Abdala et al., 2001; Flores et al., 2003), microbiotéridos (Giannini et al., 2004) y dasyúridos australianos (Flores et al., 2006). Estos estudios mostraron transformaciones morfológicas-morfométricas del cráneo, relacionadas con cambios en la dieta, durante etapas tempranas del desarrollo postdestete. Adicionalmente, demostraron que el neurocráneo y esplacnocráneo se modifican de diferentes modos y que estas modificaciones no son independientes, sino que tienen profundas influencias recíprocas en el crecimiento de ambas regiones, lo cual también es esperable en mamíferos placentarios con hábitos carnívoros estrictos.
Las crías de P. concolor tienen una masa promedio de 400 g en el momento del nacimiento. Los incisivos deciduos emergen a la edad de 10 a 20 días, seguido por los caninos (20 a 30 días) y premolares (30 a 50 días) (Eaton y Verlander, 1977); los incisivos permanentes empiezan a reemplazar los elementos deciduos a la edad de cinco meses y medio y los caninos definitivos emergen a los ocho meses (Currier, 1983). Los jóvenes permanecen con la madre hasta la edad de un año y medio a dos, y la edad de madurez sexual en las hembras sucede a los 2 ó 3 años de edad (Young y Goldman, 1946; Eaton y Verlander, 1977; Shaw et al., 2007).
Los resultados alcanzados en este trabajo son novedosos y podrían obtenerse patrones comunes de modificaciones ontogenéticas, mediante la comparación con otros grupos de carnívoros y mamíferos en general previamente estudiados (e.g. Saunders, 1961; Andersen y Wiig, 1984; Kritzman y Dudley, 1986; García- Perea, 1991, 1996; Beltrán y Delibes, 1993; Abdala et al., 2001; Hattori et al., 2003; Flores et al., 2003, 2006; Giannini et al., 2004).
En este estudio, realizamos una descripción de los cambios morfológicos craneanos ocurridos en P. concolor y sus posibles consecuencias funcionales durante el crecimiento ontogenético. Finalmente, ya que todos los félidos adquieren una alimentación estrictamente carnívora en jóvenes y adultos, los resultados obtenidos de los cambios morfofuncionales a través de la ontogenia, son interesantes en un contexto comparativo con félidos de menor tamaño (e.g. Lynx, García-Perea, 1996).
MATERIALES Y MÉTODOS
En este trabajo elegimos a P. concolor para el estudio de la variación ontogenética, por ser una especie con buena disponibilidad de especímenes de diferentes edades en colecciones sistemáticas. Aunque algunos aspectos morfológicos y reproductivos han sido relativamente bien estudiados en ésta (e.g. Pocock, 1916, 1917; Young y Goldman, 1946; Eaton y Verlander, 1977; Currier, 1979, 1983; Gay y Best, 1996a, b; Shaw et al., 2007), el conocimiento de su ontogenia está más enfocado hacia cambios en la masa corporal y recambio dentario que a las modificaciones estructurales craneanas y sus consecuencias funcionales.
En este estudio, analizamos una serie ontogenética de 98 cráneos de P. concolor depositados en la Colección Mamíferos Lillo de la Universidad Nacional de Tucumán (CML) y el Museo Argentino de Ciencias Naturales «Bernardino Rivadavia » (MACN): CML: 234; 235; 443; 650; 787; 788; 3726; 4623; 5483; 5484; 5485; 6197; 6285; 6338; 6354; 6355; 6356; 6357; 6358; 6359; 6360; 6362; 6363; 7252; 7354; 7369. MACN: 13045; 13046; 13047; 13048; 13059; 13074; 13328; 13330; 13339; 13340; 13341; 13342; 13343; 13344; 13345; 13346; 13457; 13458; 13459; 13460; 13461; 13.462; 13463; 14024; 15307; 19244; 20629; 20630; 20.64; 214; 24.19; 25.152; 25.208; 25.61; 26.9; 28.168; 29.240; 29.841; 29.891; 30.19; 30.196; 30.214; 30.215; 30.25; 30.250; 30.251; 3.15; 31.57; 32.80; 34.325; 36.614; 36.615; 36.689; 36.939; 37.27; 37.28; 38.44; 39.208; 41.222; 4.422; 47.2; 47.405; 48.344; 48.345; 49.295; 49.296; 49.470; 50.46; 53.58; 53.59; 53.97; 7.39.
En nuestra muestra, 18 especímenes no tienen aún la dentición definitiva completamente erupcionada, mientras que los restantes ejemplares son adultos ubicados en diferentes estadíos de edad. En base al conocimiento previo de la edad absoluta de los estadíos de reemplazo y desgaste dentario en P. concolor (Eaton y Verlander, 1977; Currier, 1979, 1983; Shaw et al., 2007), puede establecerse que los ejemplares más jóvenes de nuestra serie (CML 787, 788) tienen entre 10 y 20 días de edad, mientras que los más adultos (CML 6354, MACN 20.64) tienen aproximadamente 10 años. Los datos precisos conocidos sobre la correlación entre cronología de erupción y desgaste dentario con la edad absoluta (e.g. Laundré et al., 2000; Shaw et al., 2007), representan una herramienta importante en la elaboración de las consecuencias funcionales del cambio morfológico craneano en la historia de vida de P. concolor.
Para el análisis de cambios de tipo de suturas, determinamos series de edades basadas en la secuencia de erupción dentaria y desgaste. Si bien existen series de edades para P. concolor definidas en trabajos previos (e.g. Gay y Best, 1996a; Laundré et al., 2000; Shaw et al., 2007), la propuesta en este trabajo apunta a maximizar la información sobre los cambios de tipos suturales en los estadíos juveniles, ya que es en este período cuando se observan las mayores modificaciones:
Clase B: Maxila con dI1 y dI2 erupcionados. Cúspides de dI3 y dC apenas visibles. Mandíbula con dc asomando, alvéolos de dp3 y dp4 cubiertos por una fina capa de hueso.
Clase J1: Maxila con dI1, dI2 y dI3 erupcionados; dC, dP2, dP3 y dP4 erupcionados. Mandíbula con di1, di2 y di3 erupcionados; dc, dp3 y dp4 erupcionados.
Clase J2: Similar a J1 pero con M1 y m1 en proceso de erupción.
Clase J3: Maxila con I1 y I2 erupcionados; I3 esbozados, dI3 aún persistentes; C esbozados; dC, dP3 y dP4 aún persistentes; M1 erupcionados. Mandíbula con i1 e i2 erupcionados; di3, dc, dp3 y dp4 aún persistentes.
Clase J4: Maxila con I3 y C en proceso de erupción. Mandíbula con i3 y c en proceso de erupción; dc, dp3 y dp4 aún persistentes.
Clase J5: Maxila con I3, C y P4 en proceso de erupción; dC, dP2 y dP3 aún persistentes. Mandíbula con i1, i2 e i3 erupcionados.
Clase J6: Maxila con I1, I2 e I3 erupcionados; C, P2, P3 y P4 en proceso de erupción. Mandíbula como J5.
Clase A1: Maxila con I1, I2 e I3 sin desgaste, I3 más alto que los restantes; C, P2, P3 y P4 sin desgaste; proceso preglenoideo desarrollado que no cubre anteriormente el cóndilo articular. Mandíbula con i1, i2 e i3 sin desgaste; c, p3 y p4 sin desgaste; m1 sin desgaste, protocónido más alto que paracónido.
Clase A2: Maxila con I1, I2 e I3 desgastados; porción lingual del metastilo del P3 desgastado, casi plano; protocono del P4 romo; proceso preglenoideo bien desarrollado que cubre anteriormente el cóndilo articular. Mandíbula con i1, i2 e i3 desgastados; paracónidos y primera cúspide accesoria del p3 desgastados; protocónido del p4 desgastados; m1 desgastados, protocónido y paracónido a la misma altura.
Clase A3: Maxila con I1, I2 e I3 notablemente desgastados, con el ápice romo e I3 al mismo nivel que los restantes; C, P2, P3 y P4 romos, muy desgastados (se observa dentina). Mandíbula con i1, i2 e i3 muy desgastados; c, p3, p4 y m1 romos (se observa dentina).
Para la nomenclatura de huesos, forámenes, procesos, suturas y sincondrosis se siguió a Kelson (1946), Evans (1993) y Gilbert (1997).
RESULTADOS Y DISCUSIÓN
En este estudio, identificamos 32 caracteres que muestran cambios en el desarrollo ontogenético de P. concolor, comparando especímenes jóvenes (aproximadamente 2 meses) con ejemplares adultos (más de 2 años). Estos cambios (Tabla 1) suceden en la mayor parte de las regiones craneanas: rostrum (4 caracteres), paladar (1), región orbitotemporal (10), occipital (5), basicráneo y bulla timpánica (5) y mandíbula (7). De los cambios listados en la Tabla 1, casi la mitad corresponden a reordenamientos de estructuras ya presentes en especímenes jóvenes (caracteres 2, 3, 6, 11, 16, 20, 21, 22, 24, 25, 28 y 31); un número importante de caracteres corresponden a terminación o crecimiento de estructuras ocurridos durante la ontogenia (5, 7, 8, 9, 10, 14, 17, 26, 27, 29 y 30), por último, la aparición de estructuras (12, 13, 15, 18, 19, 23 y 32), así como el decremento o desaparición de éstas en los adultos (1 y 4), son las modificaciones menos frecuentes en la ontogenia craneana de P. concolor. La frecuencia de los tipos de cambios observados, difieren de lo evidenciado en otros mamíferos con fuertes tendencias carnívoras, como el marsupial Dasyurus albopunctatus (Flores et al., 2006). Siete caracteres están presentes únicamente en los adultos y ausentes en jóvenes de P. concolor, mientras que en Dasyurus diez caracteres (casi un tercio de los cambios observados) representan apariciones en la estructura craneana de los adultos. Estas observaciones indican que la mayor parte de las estructuras están ya presentes o esbozadas en los jóvenes de P. concolor y que la mayor parte de los cambios ocurridos durante el crecimiento apuntan a reordenamientos de forma, posición y aumento de tamaño general de las estructuras. Es probable entonces que los cambios tendientes a la adquisición de caracteres relacionados con la carnivoría observados en P. concolor, representen patrones ontogenéticos propios de félidos.
Tabla 1. Lista de cambios morfológicos observados en especímenes jóvenes y adultos de Puma concolor, indicando la figura en la que el carácter es observado.

Muchas de las modificaciones observadas (Tabla 1) tienden al fortalecimiento y reordenamiento del aparato masticatorio y musculatura cervical, como fuera observado también en Lynx (García-Perea, 1996) y en otros carnívoros como el lutrino Enhydra (Hattori et al., 2003) y el hiénido Crocuta (Binder y Van Valkenburgh, 2000). Estas transformaciones incluyen no solo alteraciones consecuencia del reemplazo dentario (Biknevicius, 1996), sino también modificaciones de los músculos masticatorios, cervicales y de la articulación temporomandibular.
Los cambios osteológicos más importantes apuntan principalmente al aumento de tamaño de las áreas de origen e inserción de los músculos temporales, maseteros y pterigoides. La presencia en los adultos del proceso supraorbital, cresta sagital y nucal altamente desarrollados (caracteres 8, 12 y 19 respectivamente; Figs. 1, 2), así como el aumento en área y profundidad de la fosa temporal (carácter 10; Figs. 1, 3) están en relación directa con el aumento del área de origen y volumen de la musculatura temporal. La cresta nucal muestra un notable crecimiento (Fig. 4), característica que probablemente enfatiza en los adultos la importancia de la mordida a nivel de los caninos, ya que las fibras posteriores del músculo temporal (originadas en la cresta nucal) son más importantes que las fibras temporales anteriores (originadas en la cresta sagital) para generar fuerzas transmitidas a los caninos (Hiiemae y Jenkins, 1969). Este patrón ha sido observado en la ontogenia de otros grupos de mamíferos con fuertes tendencias carnívoras, tales como en los marsupiales Lutreolina (Flores et al., 2003) y Dasyurus (Flores et al., 2006) y es característico de los adultos de grandes felinos (Duckler, 1998; Antón y Galobart, 1999; Antón et al., 2004). Adicionalmente, la ausencia de constricción postorbital (carácter 15) y la caja cerebral redondeada y comparativamente grande en jóvenes (Fig. 1A), brindan un espacio limitado para la musculatura temporal. En cambio, la constricción postorbital está presente en adultos y la caja cerebral se muestra relativamente menor y con crestas evidentes, lo que sumado a la notable expansión lateral de los arcos zigomáticos brinda un espacio considerable para el aumento en volumen de la musculatura temporal (Fig. 1B).
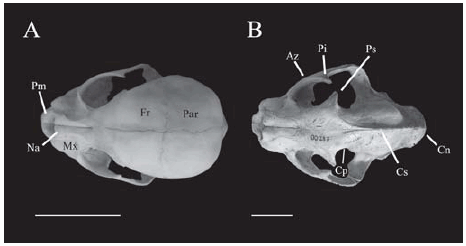
Fig. 1. Vista dorsal del cráneo de Puma concolor. A, joven de aproximadamente dos meses (CML 7369); B, adulto de aproximadamente cinco años (CML 6285). Az, Arco zigomático; Cn, Cresta nucal o lambdoidea; Cp, Constricción postorbital; Cs, Cresta sagital; Fr, Frontal; Mx, Maxilar; Na, Nasales; Par, Parietal; Pi, Proceso infraorbital; Pm, Premaxilar; Ps, Proceso supraorbital. Escala: 5 cm.

Fig. 2. Vista lateral del cráneo de Puma concolor. A, joven (CML 7369), B, adulto (CML 6285). Edades aproximadas como en Figura 1. Al, Aliesfenoides; b, borde de origen del masetero; Bu, Bulla timpánica; C, Canino definitivo; Cn, Cresta nucal; Co, Cóndilo occipital; Cs, Cresta sagital; dC, Canino deciduo; dP3 Premolar deciduo 3; dP4, Premolar deciduo 4; Es, Escamosal; fi, Foramen infraorbital; Fr, Frontal; ha, Hamulus del pterigoides; Ju, Yugal; La, Lacrimal; lm, Línea masetérica del yugal; Ma, Mastoideo; Mae, Meato auditivo externo; Mx, Maxilar; P2, Premolar definitivo 2; P3, Premolar definitivo 3; P4 Premolar definitivo 4; Pal, Palatino; Par, Parietal; Pi, Proceso infraorbital; Pm, Premaxilar; Ppa, Proceso paracondilar; Ppr, Proceso preglenoideo; Ps, Proceso supraorbital; Pz, Proceso zigomático del escamosal; So, Supraoccipital. Escala: 5 cm.

Fig. 3. Vista ventral del craneo de Puma concolor. A, joven (CML 7369), B, adulto (CML 6285). Edades aproximadas como en Figura 1. Al, Aliesfenoides; aM1, Alveolo molar definitivo 1; Az, Arco zigomático; Bf, Basiesfenoides; Bo, Basioccipital; Bu, Bulla timpánica; C, Canino definitivo; Co, Cóndilo occipital; dC, Canino deciduo; dI1, Incisivo deciduo 1; dI3, Incisivo deciduo 3; dP2, Premolar deciduo 2; dP3, Premolar deciduo 3; dP4, Premolar deciduo 4; Fg, Fosa glenoidea; Fj, Foramen yugular; Fm, Foramen magnum; Fo, Foramen oval; Ju, Yugal; Mx, Maxilar; P2, Premolar definitivo 2; P3 Premolar definitivo 3; P4, Premolar definitivo 4; Pal, Palatino; Pf, Presfenoides; Pm, Premaxilar; Pp, Proceso postglenoideo; Ppr, Proceso preglenoideo; Pt, Pterigoides; Te, Canal para el Tubo de Eustaquio; Vo, Vómer. Escala: 5 cm.

Fig. 4. Vista occipital del cráneo de Puma concolor. A, joven (CML 7369); B, adulto (CML 6285). Edades aproximadas como en Figura 1. Bu, Bulla Timpánica; Cn, Cresta nucal; Co, Cóndilo occipital; Eo, Exoccipitales; Fm, Foramen magnum; Es, Escamosal; Ip, Interparietal; Ma, Mastoideo; Par, Parietal; Pma, Proceso mastoideo; Pp, Proceso Postglenoideo; Ppa, Proceso paracondilar; So, Supraoccipital. Escala: 3 cm.
En relación con el músculo masetero, la robustez del proceso zigomático del escamosal y la línea masetérica del yugal bien desarrollada (caracteres 9 y 13 respectivamente; Fig. 2) determinan áreas de origen particularmente fuertes en los adultos. Su inserción en la mandíbula se ve particularmente reforzada en éstos, ya que una serie de estructuras y reordenamientos ocurridos, tienden a aumentar la resistencia y el área de inserción de este músculo (proceso coronoides muy amplio [carácter 28]; fosa masetérica muy desarrollada y profunda [29]; cresta coronoidea muy desarrollada [30]; y línea masetérica bien marcada y formando un borde [32]; Fig. 5). Además, el proceso angular más desarrollado en los adultos (carácter 26; Fig. 5), incrementa la resistencia y el área de inserción del masetero superficial y la inserción interna del pterigoides medio. De acuerdo a Biknevicius y Leigh (1997), la osificación postnatal de la mandíbula consta de dos fases que apuntan al reforzamiento del cuerpo mandibular, aumentando su resistencia mecánica. Esta característica es claramente visible en el borde ventral del cuerpo (carácter 31; Fig. 5). También relacionado con la función masticatoria, el mayor desarrollo de los procesos paracondilares (carácter 17; Fig. 4) indican un incremento en el área de origen de la fascia posterior del m. digastricus en los adultos.

Fig. 5. Vista lateral y oclusal de la mandíbula de Puma concolor. A-B, joven (CML 7369), CD, adulto (CML 6285). Edades aproximadas como en Figura 1. am1, Alveolo molar definitivo 1; c, Canino definitivo; cc, Cresta coronoidea; Con, Proceso condiloideo; dc, Canino deciduo; dp3, Premolar deciduo 3; dp4, Premolar deciduo 4; Fma, Fosa masetérica; m1, Molar definitivo 1; p3, Premolar definitivo 3; p4, Premolar definitivo 4; Pa, Proceso angular; Pc, Proceso coronoides; ra, Rama ascendente. Escala: 3 cm.
Los cambios en la región temporomandibular son notables. La fosa glenoidea se hace profunda, con los procesos pre y postglenoideo bien desarrollados (carácter 14; Figs. 2, 3), contribuyendo a la firmeza de esta articulación, la cual está sometida a un fuerte estrés mecánico en los adultos. Adicionalmente, el crecimiento de estos procesos, así como la expansión lateral de los procesos condiloideos de la mandíbula (carácter 27; Fig. 5), limitan el desplazamiento rostro-caudal y los movimientos transversales, restringiendo la movilidad de la mandíbula a un plano vertical (Biknevicius y Van Valkenburgh, 1996). Estas tendencias fueron también evidenciadas, aunque en menor grado, en marsupiales carnívoros (Flores et al., 2003, 2006).
En la porción occipital del cráneo (Figs. 2, 4), la parte posterior de la cresta sagital y la parte dorsal de la cresta nucal, determinan la zona de inserción de importantes músculos cervicales (Wickland et al., 1991; Richmond et al., 1992; Evans, 1993; Duckler, 1998; Antón et al., 2004). En los grandes félidos, la morfología de la musculatura cervical no ha sido aún bien estudiada, con excepción de los aportes de Antón y Galobart (1999) y Antón et al. (2004) y la detección de anomalías morfológicas producto de la hiperactividad de músculos cervicales en Panthera tigris (e.g. Duckler, 1998). La porción mastoidea altamente desarrollada en ejemplares más adultos, así como la proyección caudal de la cresta nucal (Fig. 4B), reflejan la importancia del m. obliquus capitis cranialis, profundamente implicado en los movimientos latero-dorsales de la cabeza al nivel de la articulación atlantooccipital (Argot, 2003; Antón et al., 2004). Las notables improntas de inserción muscular en la parte superior y lateral de la placa occipital de los adultos (Fig. 4B) revelan el alto grado de desarrollo de la musculatura originada en los procesos espinosos de las vértebras cervicales y torácicas más anteriores, tales como los Mm. rectus capitis dorsalis, spinalis capitis y splenius. Esta musculatura asociada a algunos movimientos de la cabeza, está también relacionada con la apertura mandibular, ya que eleva la línea dentaria superior mediante la fuerza ejercida en la zona occipito-dorsal del cráneo, acompañando la contracción del m. digastricus, el cual deprime ventralmente la mandíbula (Biknevicius y Van Valkenburgh, 1996; Antón et al., 2004). El cambio hacia una orientación caudal del foramen magnum en los adultos (carácter 16; Fig. 4), más alineado con el plano horizontal, sugiere la existencia de reordenamientos en la posición del cráneo con relación a las vértebras cervicales, así como en su musculatura asociada. El hecho de que los músculos insertados en la región occipital realizan acciones similares (i.e. elevación del cráneo), indica que el incremento de algunos movimientos enfatiza el desarrollo de determinadas áreas del cráneo (Duckler, 1998). El cambio de comportamiento hacia la adquisición de hábitos predadores incluye la reiteración de acciones que someten ciertas áreas específicas (cresta sagital, región occipital, rostro) a una fuerte demanda mecánica. En estas áreas se producen modificaciones estructurales que apuntan al fortalecimiento general del cráneo. Mientras que los cambios en las crestas sagital y nucal resultan de una elevada actividad muscular de la mandíbula, las modificaciones en el borde posterior de la cresta nucal y placa occipital, son condicionadas por los movimientos cervicales.
Adicionalmente, detectamos cambios ontogenéticos de la mayor parte de las suturas y sincondrosis craneanas en los estadíos de edad definidos (Tabla 2). Al igual que las alteraciones de forma, la tendencia general de las transformaciones suturales es incrementar el fortalecimiento y la rigidez del cráneo en los adultos. Además, existe un aumento en la absorción del estrés mecánico producto de la masticación activa en estas zonas, en comparación con la menor resistencia que muestra el tejido óseo continuo (Rafferty y Herring, 1999; Mao et al., 2003; Byron et al., 2004; Wang et al., 2006; Moazen et al., 2009). La mayor parte de las suturas sufren cambios en los diferentes estadíos de edad antes de alcanzar la dentición definitiva (Tabla 2), llegando a su fusión total en los especímenes de edad más avanzada (aproximadamente 10 años; Tabla 2), reflejando en este último caso el final del crecimiento craneal, ya que las suturas son unidades temporarias que representan zonas de crecimiento (Wagemans et al., 1988; Herring y Teng, 2000; Wang et al., 2006; Popowics y Herring, 2007; Moazen et al., 2009). Sin embargo, algunas suturas se muestran invariantes a través del crecimiento (numeradas como 3, 10, 13, 15, 21, 22, 23, 24, 25, 27, 34, 35, 39, 40, 41, 42, 43, 44, 45 en Tabla 2). De los tipos de uniones identificados, el tipo aserrada resulta altamente resistente a los requerimientos mecánicos, debido a las interdigitaciones que refuerzan la unión entre dos o más elementos, incrementando la superficie para el anclaje de fibras de colágeno resistentes a la compresión (Jaslow, 1990; Evans, 1993; Rafferty y Herring, 1999; Byron et al., 2004). Muchas suturas, entre las que se incluyen la mayor parte de las uniones del rostro, pasan del tipo plana a aserrada (numeradas como 4, 5, 7, 8, 16, 30, 31, 32, 33 en Tabla 2). Asimismo, la mayoría de las suturas localizadas en la conexión neurocráneoesplacnocráneo, pasan del tipo escamosa (donde se observa un creciente solapamiento óseo) a aserrada (numeradas como 12, 14, 28, 29, 37, 38 en Tabla 2), sumando resistencia a esta importante región del cráneo.
Tabla 2. Cambios ontogenéticos en la forma de suturas y sincondrosis craneanas de Puma concolor. NF= No Fusionada; NE= No existe; CC= Contacto cercano; ?= no observada. Las celdas rellenas indican fusión parcial (más claro) y total (más oscuro). Estadíos de edad definidos en Materiales y Métodos.
El análisis de la secuencia de cambios de las suturas, indica que la mayor parte de éstas empiezan su fusión después de haber alcanzado la dentición definitiva (aproximadamente a partir del año de edad; A1-A3 en Tabla 2), agregando resistencia frente al creciente estrés mecánico sufrido en los especímenes más adultos. No obstante, las suturas occipitales e interparietales sufren una fusión notablemente temprana en comparación con el resto (Tabla 2). En un sentido general, las suturas del neurocráneo se fusionan antes que las del esplacnocráneo (Tabla 2), otorgando rigidez a la caja cerebral y a las áreas de inserción muscular temporal y cervical. La fusión temprana del neurocráneo es también un indicador de la temprana finalización del crecimiento de las cápsulas sensoriales, lo cual está en relación con la alometría negativa de las estructuras neurocraneanas en la mayor parte de los mamíferos (Emerson y Bramble, 1993). La secuencia de osificación de la sincondrosis interesfenoidalis en la base del cráneo sigue un patrón similar al descripto para Lynx (García-Perea, 1996). La osificación de ésta comienza luego de haber alcanzado la dentición definitiva y finaliza en la fusión total en los estadíos de edad terminales (Tabla 2). En algunos especímenes asignados a estadíos adultos intermedios (A2 en Tabla 2), la sincondrosis se reduce a un orificio que marca el límite entre ambos elementos óseos (basiesfenoides y presfenoides), lo cual también es observado en Lynx lynx (García-Perea, 1996), aunque la osificación total observada en especímenes adultos terminales de P. concolor refleja el patrón observado en L. pardinus (García-Perea, 1996).
CONCLUSIONES
Los cambios ocurridos en la ontogenia craneana de P. concolor se enfocan hacia el fortalecimiento de áreas específicas asociadas al origen e inserción de musculatura temporal, masetérica y cervical. Si bien la mayor parte de las estructuras craneanas están precozmente presentes en los especímenes jóvenes, el crecimiento general y reforzamiento de éstas crea el marco adecuado para el desarrollo de un cráneo altamente eficaz en la resistencia a las exigencias mecánicas que plantean los hábitos alimenticios de los adultos. Similarmente, las suturas muestran cambios ontogenéticos que apuntan hacia la misma tendencia de reforzamiento, siendo los tipos de cambios más comunes desde plana o escamosa a aserrada. Efectivamente, la mayoría de las suturas que sirven de conexión entre neurocráneo y esplacnocráneo se tornan aserradas en los estadíos más terminales de juveniles y adultos. La fusión de las suturas y sincondrosis es la condición más común en los estadíos finales de los adultos, aunque algunas (principalmente relacionadas con los nasales) no muestran fusión a través del crecimiento y otras (relacionadas con la placa occipital e interparietal) se fusionan precozmente (Tabla 2). Por último, las tendencias ontogenéticas cualitativas de P. concolor mostraron características propias, diferentes de las observadas en otros mamíferos con hábitos carnívoros (i.e. presencia de la mayor parte de las estructuras en especímenes jóvenes, fusión previa de las suturas asociadas al neurocráneo), las que probablemente representan tendencias propias de la familia, dada su similitud con algunas observadas en otros felinos (e.g. Lynx en García- Perea, 1996). Análisis cuantitativos futuros (tanto de alometría bivariada como multivariada) son importantes para contrastar las observaciones morfológicas detectadas en este estudio y su funcionalidad. La comparación de los resultados obtenidos con los patrones ontogenéticos en otras especies de félidos más estrechamente relacionadas con P. concolor, en un contexto cualitativo y cuantitativo, son temas pendientes fundamentales ya que se espera que formas de menor tamaño corporal, como P. yagouaroundi, muestren variaciones a partir de lo expuesto en P. concolor, como consecuencia de distintos patrones de crecimiento y demandas funcionales asociadas al diferente tamaño de sus presas.
AGRADECIMIENTOS
Por permitir el acceso al material bajo estudio agradecemos a Rubén Barquez (CML) y Esperanza Varela (MACN). Julio Rojo colaboró significativamente con el apoyo en la obtención del material fotográfico en el Museo de Buenos Aires. Agradecemos a Norberto Giannini y Julio Rojo por las discusiones. Los aportes de Sergio Vizcaíno, Richard Fariña y Francisco Prevosti fueron muy importantes para mejorar la calidad del manuscrito. Parte de los fondos de esta investigación fueron provistos por el CONICET.
LITERATURA CITADA
1. ABDALA F, DA FLORES y NP GIANNINI. 2001. Postweaning ontogeny in the skull in Didelphis albiventris. Journal of Mammalogy 82:190-200. [ Links ]
2. ANDERSEN T y O WIIG. 1984. Growth of the skull of Norwegian lynx. Acta Theriologica 29:89-100. [ Links ]
3. ANTÓN M y G GALOBART. 1999. Neck function and predatory behavior in the Scimitar toothed cat Homotherium latidens (Owen). Journal of Vertebrate Paleontology 19:771-784. [ Links ]
4. ANTÓN M, MJ SALESA, JF PASTOR, IM SÁNCHEZ, S FRAILE y J MORALES. 2004. Implications of the mastoid anatomy of larger extant felids for the evolution and predatory behaviour of Sabretoothed cats (Mammalia, Carnivora, Felidae). Zoological Journal of the Linnean Society 140:207-221. [ Links ]
5. ARGOT C. 2003. Functional-adaptative anatomy of the axial skeleton of some extant marsupials and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. Journal of Morphology 255:279-300. [ Links ]
6. BELTRÁN JF y M DELIBES. 1993. Physical characteristics of Iberian lynxes (Lynx pardinus) from Doñana, southwestern Spain. Journal of Mammalogy 74:852- 862. [ Links ]
7. BIKNEVICIUS AR. 1996. Functional discrimination in the masticatory apparatus of juvenile and adult cougars (Puma concolor) and spotted hyenas (Crocuta crocuta). Canadian Journal of Zoology 74:1934- 1942. [ Links ]
8. BIKNEVICIUS AR y B VAN VALKENBURGH. 1996. Design for killing: Craniodental adaptations of predators. Pp. 393-428, en: Carnivore behavior, ecology, and evolution, Vol. 2. (JL Gittleman, ed.). Cornell University Press, New York. [ Links ]
9. BIKNEVICIUS AR y SR LEIGH. 1997. Patterns of growth of the mandibular corpus in spotted hyenas (Crocuta crocuta) and cougars (Puma concolor). Zoological Journal of the Linnean Society 120:139- 161. [ Links ]
10. BINDER WJ y B VAN VALKENBURGH. 2000. Development of bite strength and feeding behavior in juvenile spotted hyenas (Crocuta crocuta). Journal of Zoology 252:273-283. [ Links ]
11. BROOM R. 1949. Notes on the milk dentition of the lion, leopard and cheetah. Annals Transvaal Museum 21:183-185. [ Links ]
12. BYRON CD, J BORKE, J YU, D PASHLEY, CJ WINGARD y M HAMRICK. 2004. Effects of increased muscle mass on mouse sagittal suture morphology and mechanics. The Anatomical Record 279A:676-684. [ Links ]
13. CARO TM. 1994. Cheetahs of the Serengeti Plains: group living in an asocial species. University of Chicago Press, Illinois. [ Links ]
14. COLLIER GE y SJ O'BRIEN. 1985. A molecular phylogeny of the Felidae: immunological distance. Evolution 39:473-487. [ Links ]
15. CROWE DM. 1975. Aspects of ageing, growth, and reproduction of bobcats from Wyoming. Journal of Mammalogy 56:177-198. [ Links ]
16. CURRIER MJP. 1979. An age estimation technique and some normal blood values for mountain lions (Felis concolor). Unpublished Ph.D. Thesis, Colorado State University, Fort Collins. [ Links ]
17. CURRIER MJP. 1983. Felis concolor. Mammalian species 200:1-7. [ Links ]
18. DUCKLER GL. 1998. An unusual osteological formation in the posterior skulls of captive tigers (Panthera tigris). Zoo Biology 17:135-142. [ Links ]
19. EATON RL y KA VERLANDER. 1977. Reproduction in the puma: biology, behavior and ontogeny. Pp. 45-70, en: The world's cats. vol.3, nº3. (RL Eaton, ed.). Carnivore Research Institute, Burke Museum, Univ. Washington, Seattle. [ Links ]
20. EMERSON SB y L RADINSKY, 1980. Functional analysis of sabertooth cranial morphology. Paleobiology 6:295-312. [ Links ]
21. EMERSON SB y DM BRAMBLE. 1993. Scaling, allometry and skull design. Pp. 384-416, en: The skull (J Hanken y BK Hall, eds.). The University of Chicago Press, Chicago, Illinois. [ Links ]
22. EVANS HE. 1993. Miller's Anatomy of the dog, 3rd edition. W.B. Saunders Company. [ Links ]
23. EWER R. 1973. The Carnivores. Cornell University Press, Ithaca. [ Links ]
24. FAGEN RM y KS WILEY.1978. Felid paedomorphosis, with special reference to Leopardus. Carnivore 1:72- 81. [ Links ]
25. FLORES DA, NP GIANNINI y F ABDALA. 2003. Cranial ontogeny on Lutreolina crassicaudata (Didelphidae): a comparison with Didelphis albiventris. Acta Theriologica 48:1-9. [ Links ]
26. FLORES DA, N GIANNINI y F ABDALA. 2006. Comparative Postnatal Ontogeny of the skull in an Australidelphian Metatherian, Dasyurus albopunctatus (Marsupialia: Dasyuromorpha: Dasyuridae). Journal of Morphology 267:426-440. [ Links ]
27. GARCÍA-PEREA R. 1991. Variabilidad morfológica del género Lynx Kerr, 1792 (Carnivora: Felidae). Ph. D. Thesis. Universidad Complutense, Madrid, España. [ Links ]
28. GARCÍA-PEREA R. 1996. Patterns of Postnatal Development in skulls of Lynxes, Genus Lynx (Mammalia: Carnivora). Journal of Morphology 229:241-254. [ Links ]
29. GARCÍA-PEREA R. 2002. Andean mountain cat, Oreailurus jacobita: Morphological description and Comparison with other felines from the altiplano. Journal of Mammalogy 83:110-124. [ Links ]
30. GAY SW y TL BEST. 1996a. Age-related variation in skulls of the puma (Puma concolor). Journal of Mammalogy 77:191-198. [ Links ]
31. GAY SW y TL BEST. 1996b. Relationships between abiotic variables and geographic variation in skulls of pumas (Puma concolor: Mammalia, Felidae) in North and South America. Zoological Journal of the Linnean Society 117:259-282. [ Links ]
32. GIANNINI N, F ABDALA y D FLORES. 2004. Comparative Postnatal Ontogeny of the skull in Dromiciops gliroides (Marsupialia: Microbiotheriidae). American Museum Novitates 3460:1-17 [ Links ]
33. GILBERT SG. 1997. Pictorial Anatomy of the cat. University of Washington Press. [ Links ]
34. GOMES DE OLIVEIRA T. 1993. Neotropical Cats, Ecology and Conservation. Edufma, Sao Luís. [ Links ]
35. HATTORI K, AM BURDIN, M SUZUKI y N OHTAISHI. 2003. Age-related change and allometry of skull and canine of Sea Otters, Enhydra lutris. Journal of Veterinary Medical Science 65:439-447. [ Links ]
36. HERRING SW y S TENG. 2000. Strain in the braincase and its sutures during function. American Journal of Physical Anthropology 112:575-593. [ Links ]
37. HIIEMAE K y FA JENKINS JR. 1969. The anatomy and internal architecture of the muscles of mastication in Didelphis marsupialis. Postilla 140:1-49. [ Links ]
38. JACKSON DL, EA GLUESING y HA JACOBSON. 1988. Dental eruption in bobcats. The Journal of Wildlife Management 52:515-517. [ Links ]
39. JASLOW CR. 1990. Mechanical properties of cranial sutures. Journal of Biomechanics 23:313-321. [ Links ]
40. JOHNSON NF, BA BROWN y JC BOSOMWORTH. 1981. Age and Sex characteristics of bobcat canines and their use in population assessment. Wildlife Society Bulletin 9:202-206. [ Links ]
41. JOHNSON WE y SJ O'BRIEN. 1997. Phylogenetic Reconstruction of the Felidae Using 16S rRNA and NADH-5 Mitochondrial Genes. Journal of Molecular Evolution 44:98-116. [ Links ]
42. KELSON KR. 1946. Notes on the comparative osteology of the bobcat and house cat. Journal of Mammalogy 27:255-264. [ Links ]
43. KRITZMAN EB y JC DUDLEY. 1986. Skull morphometry of Western Washington Bobcats (Felis rufus fasciatus). Actas of 66th Annual Meeting of American Society of Mammalogists. University of Wisconsin. [ Links ]
44. LAUNDRÉ JW, L HERNANDEZ, D STREUBEL, K ALTENDORF y CL GONZALEZ. 2000. Aging Mountain Lions using Gum-Line Recession. Wildlife Society Bulletin 28:963-966. [ Links ]
45. LUCHERINI M, L SOLER y E LUENGOS VIDAL. 2004. A preliminary revision of knowledge status of felids in Argentina. Mastozoología Neotropical 11:7- 17. [ Links ]
46. MAO JJ, X WANG y RA KOPHER. 2003. Biomechanics of Craniofacial Sutures: Orthopedic Implications. Angle Orthodontist 73:128-135. [ Links ]
47. MARTIN LD. 1980. Functional Morphology and the evolution of cats. Transactions of the Nebraska Academy of Sciences 8:141-154. [ Links ]
48. MARTIN LD. 1996. Fossil History of the Terrestrial Carnivora. Pp.536-568, en: Carnivore behavior, ecology, and evolution. Vol 1. (JL Gittleman, ed.). Cornell University press. [ Links ]
49. MATTERN MY y DA MC LENNAN. 2000. Phylogeny and Speciation of Felids. Cladistics 16:232-253. [ Links ]
50. MOAZEN M, N CURTIS, P O'HIGGINS, MEH JONES, SE EVANS y MJ FAGAN. 2009. Assessment of the role of sutures in a lizard skull: a computer modelling study. Proceedings of the Royal Society 276:39- 46. [ Links ]
51. PETERSEN MK y MK PETERSEN. 1978. Growth rate and other postnatal developmental changes in margays. Carnivore 1:87-92. [ Links ]
52. POCOCK RI. 1916. The structure of the auditory bulla in existing species of Felidae. Annals and Magazine of Natural History 18:326-334. [ Links ]
53. POCOCK RI. 1917. On the external characters of the Felidae. Annals and Magazine of Natural History 19:113-136. [ Links ]
54. POPOWICS TE y SW HERRING. 2007. Load transmission in the nasofrontal suture of the pig, Sus scrofa. Journal of Biomechanics 40:837-844. [ Links ]
55. RAFFERTY KL y SW HERRING. 1999. Craniofacial Sutures: Morphology, Growth, and in Vivo masticatory Strains. Journal of Morphology 242:167-179. [ Links ]
56. RICHMOND FJR, DB THOMSON y GE LOEB. 1992. Electromyographic studies of neck muscles in the intact cat. Experimental Brain Research 88:41-58. [ Links ]
57. SALLES LO. 1992. Felid phylogenetics: Extant taxa and skull morphology (Felidae, Aeluroidea). American Museum Novitates 3047:1-67. [ Links ]
58. SAUNDERS JK. 1961. The biology of the Newfoundland lynx. Ph.D. Thesis. Cornell University. [ Links ]
59. SHAW HG, P BEIER, M CULVER y M GRIGIONE. 2007. Puma Field Guide. The Cougar Network. [ Links ]
60. SLAUGHTER BH, RH PINE y NE PINE. 1974. Eruption of cheek teeth in Insectivora and Carnivora. Journal of Mammalogy 55:115-125. [ Links ]
61. STANDER PE. 1997. Field age determination of Leopards by tooth wear. African Journal of Ecology 35:156-161. [ Links ]
62. STEHLIK J. 1971. Breeding jaguars Panthera onca at Ostrava Zoo. International Zoo Yearbook 11:116- 118. [ Links ]
63. STUART CHT y TD STUART. 1985. Age determination and development of foetal and juvenile Felis caracal Schreber, 1776. Säugertierkunde Mitteilungen 32:217-229. [ Links ]
64. VOLF J. 1972. Exigences alimentaires et dentition des jeunes de trois especes de felides. Mammalia 36:683- 686. [ Links ]
65. WAGEMANS PAHM, JP VAN DE VELDE y AM KUIJPERS-JAGTMAN. 1988. Sutures and forces: A review. American Journal of Orthodontics and Dentofacial Orthopedics 94:129-141. [ Links ]
66. WANG Q, DS STRAIT y PC DECHOW. 2006. Fusion Patterns of Craniofacial Sutures in Rhesus Monkey Skulls of Known Age and Sex from Cayo Santiago. American Journal of Physical Anthropology 131:469- 485. [ Links ]
67. WICKLAND CR, JF BAKER, BW PATERSON. 1991. Torque vectors of neck muscles in the cat. Experimental Brain Research 84:64-659. [ Links ]
68. WOZENCRAFT WC. 1996. The Phylogeny of the Recent Carnivora. Pp. 569-593, en: Carnivore behavior, ecology, and evolution. Vol. 1. (JL Gittleman, ed.). Cornell University Press. [ Links ]
69. XIMÉNEZ A. 1974. Notas sobre Félidos Neotropicales, VI. Contribución a la elucidación de las variaciones individuales de Felis pardalis Linne, 1758 (Mammalia, Felidae). Comunicaciones del Museo Argentino de Ciencias Naturales «Bernardino Rivadavia» e Instituto Nacional de Investigación de las Ciencias Naturales 4:41-55. [ Links ]
70. YOUNG SP y EA GOLDMAN. 1946. The Puma, mysterious American cat. The American Wildlife Institute., Washington D.C. [ Links ]
Recibido 6 agosto 2007.
Aceptado 3 diciembre 2007.
Editor asociado: SF Vizcaíno

