










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. v.17 n.1 Mendoza ene./jun. 2010
ARTÍCULOS Y NOTAS
Dieta de Lontra Longicaudis (Carnivora,Mustelidae) en el Parque Nacional El Rey (Salta, Argentina) y su comparación con otras poblaciones de la cuenca del Paraná
Silvina B. Chemes1,2, Alejandro R. Giraudo1,2 y Guillermo Gil3
1 Facultad de Humanidades y Ciencias, Universidad Nacional del Litoral, Ciudad Universitaria, Paraje El Pozo, S3000ZAA Santa Fe, Argentina [Correspondencia: Silvina B. Chemes <schemes@fhuc.unl.edu.ar>].
2 Instituto Nacional de Limnología (CONICET-UNL), Ciudad Universitaria, Paraje El Pozo, S3000ZAA Santa Fe, Argentina.
3 Administración de Parques Nacionales, Delegación Regional Nordeste, Av. Tres Fronteras 183, N3370AWB Puerto Iguazú, Misiones, Argentina
RESUMEN: Se analizó la dieta de Lontra longicaudis en el Parque Nacional El Rey, en la eco-región de las Yungas, noroeste argentino. Se compararon estos datos con los de poblaciones previamente estudiadas en la laguna Iberá (Esteros del Iberá) y en el río Iguazú (Selva paranaense) en el nordeste argentino. Se analizaron 130 heces colectadas en 37 sitios evaluando porcentaje de ocurrencia, frecuencia relativa, importancia relativa porcentual y diversidad de Levin. Se comparó con otras poblaciones mediante el índice de similitud porcentual, re-muestreo por bootstrap y análisis no paramétricos. De un total de 623 presas, peces, insectos y crustáceos fueron más frecuentes que anfibios, moluscos, reptiles y mamíferos. La mayoría de las presas fueron organismos bentónicos. Nuestros resultados fueron más similares a uno de los estudios del Iberá. Otros dos estudios, del Iberá y del Iguazú, publicados por el mismo autor, fueron más semejantes entre sí. La adaptabilidad trófica de L. longicaudis a diferencias en disponibilidad y variación temporal de las presas o los sesgos metodológicos entre autores, podrían ser causas de las diferencias en las dietas de las poblaciones estudiadas en la cuenca del Paraná.
ABSTRACT: Diet of Lontra longicaudis (Carnivora, Mustelidae) in El Rey National Park (Salta, Argentina) and its comparison with other populations from Paraná basin. It was analyzed the diet of the Lontra longicaudis in El Rey National Park, within the Yungas forest eco-region, in the Argentine Northwest. Our data was compared with that from other populations previously studied in the Iberá Lagoon (Iberá marshes) and the Iguazú River (Paranaense forest) in the Argentine Northeast. We analyzed 130 feces from 37 latrines. Percentage of occurrence, relative frequency, percentage of relative importance and Levin's diversity were analyzed. We compared our results with those from other populations by means of the percentage similarity index, the re-sampling by bootstrapping and non-parametric analysis. We recorded 623 prey items. Fish, insects and crustaceans were more frequently consumed than amphibians, mollusks, reptiles and mammals. The majority of preys were benthonic organisms. Our results were more similar to the ones obtained in the Iberá Lagoon. Other studies in the Iberá and the Iguazú, published by the same author, were more similar between themselves. Differences in trophic adaptability, availability, temporal variation of preys or methodological biases among authors could be possible reasons for the diet differences of the populations studied in Argentina.
Palabras clave. Argentina; Dieta; Lontra longicaudis; Yungas.
Key words. Argentina; Diet; Lontra longicaudis; Yungas.
INTRODUCCIÓN
El nicho ecológico de un organismo está definido por la suma total de las variables ambientales de su entorno, tales como su posición trófica, fuentes de energía y repartición de recursos (Odum y Warrett, 2006). La dieta del animal emerge de la interrelación entre su conducta de forrajeo, distribución y abundancia del alimento y de sus necesidades fisiológicas. Las especies generalistas se alimentan de un amplio espectro de alimentos en términos de números de especies y del microhábitat en el cual viven y no poseen una marcada preferencia por una fuente de alimento. En cambio, los especialistas presentan su dieta restringida a un número relativamente pequeño de especies (Gerking, 1994). La ocurrencia de especialistas o generalistas en un determinado hábitat está influenciada por la dinámica de los recursos alimentarios. Se ha reconocido un predominio de generalistas en ríos y especialistas en lagos; y que la especialización no es típica de hábitats efímeros (Lowe-McConnell, 1987; Araujo Lima et al., 1995). Un gran número de especies tiene la capacidad de cambiar sus hábitos alimentarios para responder a las variaciones estacionales y/o espaciales en la disponibilidad de alimento. Esta flexibilidad trófica implica la habilidad para sacar provecho de la fuente de alimento más ventajosa en un determinado tiempo o lugar (Gerking, 1994). Son pocos los estudios sobre respuesta funcional de los predadores, selección y vulnerabilidad de las presas de mamíferos carnívoros en la región Neotropical (Bueno y Motta-Junior, 2006).
Los carnívoros cumplen funciones importantes en los ecosistemas por su rol en las redes tróficas, tales como controlar la calidad de las poblaciones de presas, participar en la dispersión de semillas, ser impulsores de procesos co-evolutivos y comportarse como superpredadores (García-Perea et al., 1996). Lontra longicaudis Olfers, 1818 es un mustélido semiacuático, encontrado en la región Neotropical desde México hasta el centro de Argentina y Uruguay, donde habita en ríos, lagunas y arroyos (Cabrera y Yepes, 1940; Wilson y Reeder, 1993; Larivière, 1999). Es considerada una especie insuficientemente conocida por la UICN (IUCN, 2006) y en peligro en Argentina (Díaz y Ojeda, 2000). Al igual que otros carnívoros, esta especie ha estado seriamente amenazada por la cacería. La destrucción de su hábitat, la contaminación acuática y su captura en redes de pesca son las causas de su baja densidad poblacional actual (García-Perea et al., 1996; Larivière, 1999). En el extremo sur de su distribución, los antecedentes sobre su ecología trófica son aún escasos (Parera, 1992, 1993a, 1993b; Gori et al., 2003). Estos lutrinos son registrados como predadores oportunistas por la alta plasticidad dietaria entre hábitats; se considera que su dieta varía en respuesta a la disponibilidad estacional de las presas y que tienden a seleccionar peces de tamaño moderado y poco móviles (Pardini, 1998; Cote et al., 2008).
En este trabajo se estudia la alimentación de una población de L. longicaudis que habita en arroyos y ríos pequeños de la eco-región de las Yungas y Chaco Serrano, en el Parque Nacional El Rey (Salta, Argentina) (Burkart et al., 1999). Esta área protegida abarca las cabeceras de la subcuenca del río del Valle, la temperatura promedio en primavera-verano es de 20.2 ºC y en otoño-invierno de 12.9 ºC, las precipitaciones anuales promedio son de 837 mm, concentradas en el período estival (Servicio Meteorológico Nacional, 2008). La región ictiológica es la Paranoplatense del Oeste, que constituye el límite oeste de distribución de la fauna paranoplatense, con especies de Characidae, Pimelodidae y Loricariidae. Se destaca la ocurrencia de especies asociadas con el rango altitudinal de esta región, apareciendo endemismos principalmente en la familia Trichomycteridae (López et al., 2002). Dentro del área se han detectado 23 especies de peces (Burkart et al., 1999; López et al., 2002; APN-SIB, 2008). Existen sólo otros dos sitios de la misma cuenca hídrica del Paraná donde se realizaron estudios previos de dieta de la especie: laguna Iberá y río Iguazú superior (Parera, 1993b; Gori et al., 2003).
En Laguna Iberá la temperatura promedio en primavera-verano es de 23.8 ºC y en otoño-invierno de 15.7 ºC, las precipitaciones anuales promedio son de 1611 mm, mientras que las estivales son ligeramente superiores (Servicio Meteorológico Nacional, 2008). Esta laguna con costas sin pendientes posee una superficie de 52.1 km2 y una profundidad promedio de 3.2 m con una variación estacional máxima de 0.4 m (Canziani et al., 2003; Cozar et al., 2003). Se encuentra en la eco-región de Esteros del Iberá y su perímetro está cubierto de vegetación palustre arraigada y flotante con algunos sectores con leñosas de porte arbóreo. La región ictiológica es el eje potámico subtropical, caracterizada por su alta diversidad y el dominio de familias de ostariofíseos, los lepidosirénidos y grupos de origen marino (Clupeidae, Engraulidae, Atherinopsidae, Belonidae, Sciaenidae, Achiridae y Potamotrygoninae) (Burkart et al., 1999; López et al., 2002). Para la laguna Iberá se registraron 87 especies de peces, de los órdenes Myliobatiformes, Clupeiformes, Characiformes, Siluriformes, Gymnotiformes, Atheriniformes, Beloniformes, Cyprinodontiformes, Symbranchiformes, Perciformes y Lepidosireniformes (Casciotta et al., 2005).
En la zona del río Iguazú superior, la temperatura promedio en primavera-verano es de 23.8 ºC y en otoño-invierno de 17.5 ºC, las precipitaciones anuales promedio son de 1919 mm, sin una época lluviosa distinguible (Servicio Meteorológico Nacional, 2008). Este tramo del río posee un ancho de entre 500 y 1350 m, costas con barrancas de hasta cuatro metros, profundidades máximas de 20 m y afloramientos del lecho rocoso en las "correderas". Su caudal medio anual de 1824 m3/s (908.4-3291 m3/s) con pequeños picos en mayo, junio y octubre (hasta 2535 m3/s) y leves bajas en diciembre, enero y marzo (hasta 1330 m3/s) (Drago y Paira, 2001; Subsecretaría de Recursos Hídricos, 2008). Este sector del río se encuentra en la eco-región de selva paranaense y sus márgenes están cubiertas de selvas ribereñas. Está incluida en la zona ictiológica misionera, caracterizada por la ausencia de ciertos grupos que son comunes en el resto de la cuenca (por ejemplo: Myliobatiformes, Clupeiformes, Pleuronectiformes, y algunos Characiformes y Siluriformes). Para el sector argentino de este río se conocen 45 especies de peces, ocho de ellas endémicas de esta subcuenca (Burkart et al., 1999; López et al., 2002).
Las tres eco-regiones son distintas, incluso ictiológicamente y sus cuerpos de agua presentan dinámicas hídricas muy diferentes. Esto generaría una disponibilidad de presas dispar que afectaría a la dieta de una especie generalista como L. longicaudis. Este trabajo tuvo como finalidad determinar la composición de la dieta de la población de L. longicaudis en las Yungas y analizar si existen variaciones entre esta población y las otras estudiadas en la misma cuenca.
MATERIALES Y MÉTODOS
Área de estudio y métodos
Se trabajó en el Parque Nacional El Rey, en la Provincia de Salta (24º 38' S, 64º 37' W), incluyendo ambientes del chaco serrano, selvas de transición y selvas montanas (Cabrera, 1976), entre 690 y 1149 m snm. En agosto de 1996 y entre agosto y noviembre de 1997, se muestreó el río Popayán y los arroyos Los Puestos, La Sala, Soco Hondo y Los Noques (Fig. 1), pertenecientes a la subcuenca del río del Valle, caracterizados como cursos de agua mesosaprobios en estiaje y oligosaprobios en épocas de aguas altas, con un nivel de contaminación bajo a moderado (Gonzo, 2003). Estos arroyos, muestreados en época de estiaje, comprendieron cursos de agua con 1 a 45º de pendiente de la costa, de corriente rápida y limpidez absoluta, poseían un ancho de uno a cinco metros aunque con lechos secos de rocas, cantos rodados y arena de hasta 60 m limitados por barrancas de unos tres metros de altura; las profundidades medias eran menores a 0.5 m con afloramientos del lecho rocoso y pozos de hasta 4 m. El caudal medio anual del río del Valle es de 4.64 m3/s, con un máximo de 8.62 m3/s y un mínimo de 1.78 m3/s, aunque la estacionalidad de las lluvias marca un pico de 13.2 m3/s en febrero y la menor lectura para septiembre con 1.65 m3/s (Gil, obs. pers.; Subsecretaría de Recursos Hídricos, 2008). La región estudiada no fue cercana a viviendas, pero se observó ganado doméstico y presencia de visitantes del parque (Gil, obs. pers.).
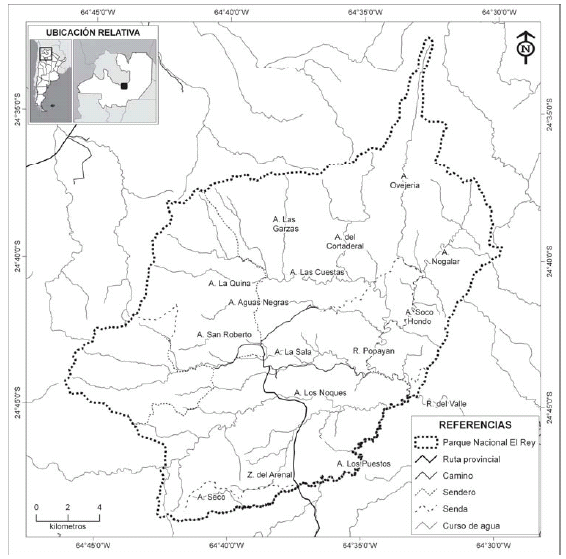
Fig. 1. Ubicación geográfica del área de estudio Parque Nacional El Rey (Salta, Argentina) (APN-SIB 2008).
Se analizaron 130 heces, incluyendo ocho heces individuales y el resto componiendo 29 letrinas que se colectaron en 11 de 20 estaciones de muestreo. Estas consistieron en transectas sobre una de las márgenes del río, de 600 m de longitud y 20 m de ancho. Cada estación insumió un día entre acceso y recorrida. Las fecas estuvieron depositadas sobre rocas, troncos o raigones descalzados en la costa. Las unidades de muestreo fueron las fecas individuales (una deposición). En las letrinas, según el caso, se estimó el número de heces que esta incluía por encontrarse físicamente separadas, con distinto color y/o tenor de humedad. Si las unidades de las fecas se hallaban disgregadas, se aproximó la cantidad por comparación de volúmenes promedios con unidades enteras (Gil, 1998).
Las fecas secas se procesaron siguiendo los métodos propuestos por Reynolds y Aebischer (1991), Carss y Parkinson (1996) y Jacobsen y Hansen (1996) con modificaciones: inmersión en agua caliente y detergente, filtrado (500 μm), enjuague con agua corriente hasta lograr transparencia en el agua residual, secado en estufa (60º C, 48 horas) y secado final mediante exposición a luz solar o calefactor. La determinación de las presas se realizó al menor nivel taxonómico posible mediante el examen de los restos y comparación con material de museos, consultas a especialistas y bibliografía (e.g., Gonzo, 2003). Se contabilizó el número mínimo de individuos de cada categoría de presa en cada feca, usando criterios de repetición de piezas pares y variaciones en la talla (Ruiz Olmo et al., 1989).
Análisis de datos
Se utilizaron los índices de porcentaje de ocurrencia (Ruiz Olmo et al., 1989), frecuencia relativa, importancia relativa porcentual (Laroche, 1982), y diversidad B de Levin (Ruiz Olmo et al., 1989). Aunque la frecuencia relativa no pondera el peso de los restos de presas ni la biomasa ingerida, constituye un método apropiado para determinar la composición y los aportes relativos de cada presa porque es simple de aplicar y sus resultados pueden ser comparados directamente con la mayoría de estudios de la misma o diferentes especies de carnívoros (Corbett, 1989). Con los datos de frecuencia relativa, la dieta fue comparada con las de otras poblaciones de la cuenca del Paraná (Parera, 1992, 1993b; Gori et al., 2003). Se utilizó: (1) el índice porcentaje de similitud (cuali-cuantitativo), (2) los test no paramétricos W de Kendall y rS de Spearman, en los que se ordenaron en rangos las frecuencias relativas de las presas en cada sitio para luego comparar estos entre sí y (3) la técnica de re-muestreo de "bootstrap" (Efron, 1979), por la que se calcularon los intervalos de confianza de cada clase de presa de los cuatro estudios comparados a partir de 1000 muestras del mismo n que cada muestra original. Se utilizaron los programas MVSP 3.1, SPSS 11.5 e InfoStat 2005.1 con un 95% de confianza (Siegel, 1983).
RESULTADOS
Dieta
Las 130 fecas contuvieron 623 presas (Tabla 1). La población estudiada en las Yungas se alimenta principalmente de peces, insectos y crustáceos, en concordancia parcial con los antecedentes bibliográficos (Rosenzvaig, 1974; Parera, 1992, 1993a, 1993b; Helder, 1997; Pardini, 1998; Colares y Waldemarin, 2000; Quadros y Monteiro Filho, 2001; Gori et al., 2003; Louzada-Silva et al., 2003; Kasper et al., 2004).
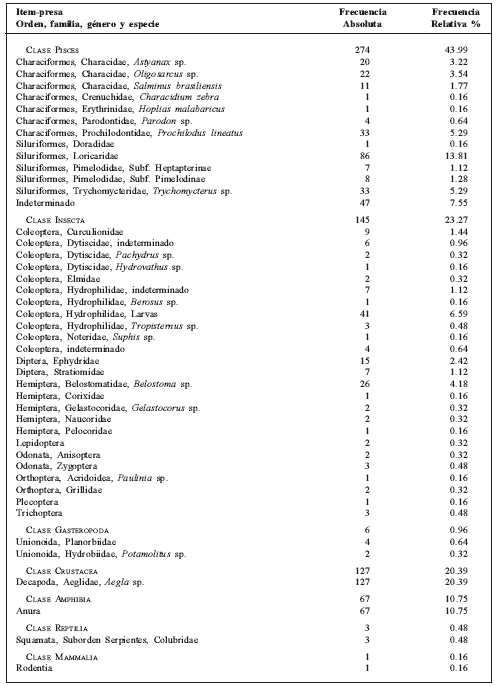
Tabla 1 Espectro trófico de Lontra longicaudis en el Parque Nacional El Rey (Salta, Argentina) sobre la base de 130 heces (623 presas).
A nivel de órdenes mostraron mayor frecuencia relativa los siluriformes y decápodos, seguidos por characiformes y coleópteros (Fig. 2).
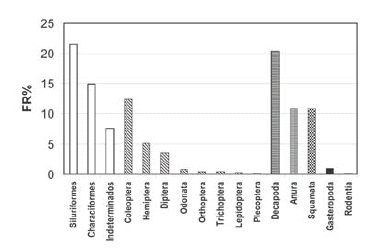
Fig. 2. Espectro trófico de Lontra longicaudis en el Parque Nacional El Rey (Salta, Argentina), en categoría de órdenes (invierno 1996, invierno y primavera 1997).
Entre los peces, a nivel de familias, los ítems con mayor frecuencia absoluta fueron Loricaridae (86 individuos), Characidae (53) y Prochilodontidae (33). Entre los insectos, los coleópteros (77) fueron los más consumidos, seguidos por dípteros (32) y hemípteros (22). Los crustáceos estuvieron representados por ejemplares de Aegla sp. (127).
Considerando la Importancia Relativa Porcentual, los peces fueron las presas más frecuentes (53%), seguidos por los insectos (24%), crustáceos (16%) y anfibios (7%). Reptiles, moluscos y mamíferos mostraron muy bajos valores, menores al 0.1%.
El Indice de Diversidad de Levin aplicado a nivel de clases, alcanzó un valor de 0.584, en un rango de 0 a 1.
Comparación con otros estudios
El índice de diversidad se aplicó también para los espectros tróficos de los otros estudios realizados en la cuenca del Paraná. Para la Laguna Iberá, Corrientes (Gori et al., 2003), resultó una diversidad aproximada (B = 0.596); y valores muy bajos para la misma laguna en un trabajo anterior (B = 0.259) (Parera, 1992) y para el río Iguazú, Misiones (B = 0.085) (Parera, 1993b).
Al comparar las proporciones a nivel de clases, mediante el porcentaje de similitud, se observó que el grupo de mayor afinidad (88%) reúne a los estudios de la laguna Iberá y el río Iguazú realizados por Parera (1992, 1993b). El segundo grupo de mayor similitud (69%) está constituido por el estudio de la laguna Iberá de Gori et al. (2003) con el presente trabajo. Similares resultados se obtuvieron al comparar estadísticamente las dietas. En conjunto, no estuvieron correlacionadas significativamente (W = 0.109; p = 0.352), pero al compararlas de a pares presentaron correlación significativa entre sí los trabajos de laguna Iberá y río Iguazú (Parera, 1992, 1993b) (rS = 0.679; p = 0.031) y el de Iberá (Gori et al., 2003) junto con el presente estudio (rS = 0.681; p = 0.03).
En la Fig. 3 se observan los intervalos de confianza calculados mediante el análisis de bootstrap, donde se destaca que la proporción de peces, el principal alimento ingerido por los lobitos en todos los estudios, fue similar entre los estudios de Iguazú e Iberá (Parera, 1992, 1993b) con proporciones significativamente mayores a las observadas en este estudio y el de Iberá de Gori et al. (2003), que mostraron proporciones similares entre sí.
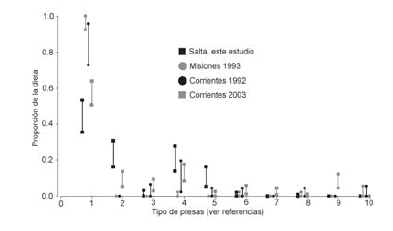
Fig. 3. Intervalos de confianza, obtenidos por bootstrap, de la proporción de ingesta de cada tipo de presa en cuatro estudios. Referencias: 1. Peces, 2. Insectos, 3. Gasterópodos, 4. Crustáceos, 5. Anfibios, 6. Reptiles, 7. Aves, 8. Mamíferos; 9. Plantas, 10. Indeterminadas.
Los peces de la familia Loricaridae fueron las presas más frecuentes en la población del Parque Nacional El Rey, coincidiendo con estudios realizados en el río Iguazú (Parera, 1993b) y en ríos de Brasil (Pardini, 1998; Kasper et al., 2004). La alta frecuencia de presas de peces de la familia Characidae en Salta es similar a lo observado por Gori et al. (2003), pero contrasta con lo registrado en las otras localidades donde están en muy baja proporción e incluso con selección negativa (Pardini, 1998; Quadros y Monteiro-Filho, 2001; Kasper et al., 2004).
La población de Yungas consumió mayor proporción de insectos respecto a las demás. Uno de los estudios de Iberá (Gori et al., 2003) contiene mayor proporción de insectos que el otro trabajo para la misma localidad y que para Iguazú (Parera 1992, 1993b). Aún así, los invertebrados (45% del total de presas), principalmente crustáceos (20%) e insectos (23%), fueron un componente relevante.
En diversos estudios de dieta, los artrópodos suelen estar ausentes (Louzada-Silva et al., 2003; Muanis y Waldemarin 2003), o con frecuencias menores al 10% (Quadros y Monteiro-Filho, 2001; Kasper et al., 2004; Uchôa et al., 2004). Sólo se destaca un caso donde superó el 21% (Pardini, 1998). En este y otro caso (Colares y Waldemarin, 2000) hubo diferencias estacionales aunque con tendencias contrarias. En el presente estudio, los insectos estuvieron representados principalmente por larvas hidrofílidas y adultos de Belostoma, mientras que los crustáceos, por Aegla. El consumo de insectos belostomátidos ya fue descrito para Rio Grande do Sul (Brasil) e Iberá (Gori et al., 2003; Kasper et al., 2004).
La cantidad de crustáceos indicados para Iguazú (Parera, 1993b) es inferior a la de las demás localidades. En poblaciones de Brasil, los crustáceos también se ubican entre los componentes más consumidos, aunque se observaron diferencias entre localidades de una misma población y disminución de consumo en verano, esto último al igual que en Iberá (Parera, 1992; Pardini, 1998; Colares y Waldemarin, 2000; Quadros y Monteiro-Filho, 2001). En un caso, camarones y cangrejos superaron el 50% (Uchôa et al., 2004).
Los anfibios fueron relativamente frecuentes en la población de Yungas (14%), mientras que no alcanzaron el 1% en los registros de los otros autores (Parera 1992, 1993b; Gori et al. 2003). En poblaciones de Brasil, fueron poco frecuentes (Pardini, 1998; Kasper et al., 2004) o estuvieron ausentes (Colares y Waldemarin, 2000; Quadros y Monteiro-Filho, 2001; Louzada-Silva et al., 2003; Muanis y Waldemarin, 2003).
Uno de los estudios de Iberá (Gori et al., 2003) fue el único que registró plantas en las fecas, aspecto que difiere significativamente de los demás estudios. Quadros y Monteiro Filho (2000) mencionan frutos entre sus alimentos en Brasil.
DISCUSIÓN
En base al amplio espectro de presas consumidas por L. longicaudis, se concluye que la especie se comporta como un predador de hábitos generalistas. Tiene un nicho trófico amplio, ya que a pesar de consumir mayormente algunos ítems (peces y artrópodos), se alimenta ocasionalmente de anfibios, reptiles, gasterópodos y pequeños mamíferos.
La oferta alimentaria disponible se encuentra condicionada por las características de hábitat y, por ende, del tipo de organismos que conforman sus comunidades (Gerking, 1994). En las Yungas, cursos lóticos con pendiente, sustratos duros y elevada transparencia que alterna con turbidez total, O2 en exceso y baja temperatura del agua con estratificación en los pozos, se asocian a la presencia de organismos con respuestas reotácticas desarrolladas, baja densidad poblacional y con dispositivos de sujeción, como loricáridos y dorádidos (Cole, 1988; Merritt y Cummins, 1996). Los peces consumidos en la subcuenca del río del Valle fueron mayormente frecuentadores del fondo (Ringuelet, 1975). En ríos de Brasil (Pardini, 1998; Kasper et al., 2004) se comprobó una selección de este tipo de presas respecto a su disponibilidad y se expuso como posible explicación la facilidad de captura por ser peces bentónicos. En el presente estudio, podrían relacionarse con una mayor oferta de loricáridos en ríos correntosos y con piedras. Otras especies que tienen el cuerpo hidrodinámico y nadan contra la corriente, como peces de las familias Characidae, Crenuchidae, Erythrinidae y Parodontidae también fueron componentes de la dieta (Ringuelet, 1975; Galvis et al., 1997).
Las comunidades ícticas se encuentran determinadas por dos componentes, uno temporal-cíclico ligado a los ciclos hidrológico y estacional, y otro espacial que incluye disponibilidad de alimento y fenómenos denso dependientes (migraciones, natalidad, mortalidad) (Quirós, 1987). Las migraciones reproductivas se han asociado a las diferencias entre caudales dentro de la cuenca, como ocurre con Prochilodus lineatus, que cuando el caudal es máximo migra para desovar en la zona andina del río (Bayley, 1973). Durante nuestro estudio, los cursos de agua se encontraban en época de estiaje y no se daban las condiciones para altas concentraciones de peces. Esto podría explicar que aun siendo presas predominantes, los peces compartieran su importancia relativa con insectos y crustáceos al momento de hacer el estudio.
Por su parte, la importancia relativa de los cangrejos Aegla en el espectro trófico de L. longicaudis se puede asociar a una respuesta funcional de tipo 2, en la que la alta tasa de consumo se relaciona con un aumento temporal de densidad de la presa. Durante el periodo de muestreo existió mayor abundancia de Aegla (Gil, obs. pers.), tanto por sus preferencias espaciales como por su estadio reproductivo. Estas poblaciones presentan disposición espacial contagiosa, prefiriendo lugares sombreados y protegidos de la fuerte corriente, como pozos profundos, troncos caídos, raíces o rocas sueltas (Swiech-Ayoub y Masunari, 2001a; Giri y Williner, 2006). Temporalmente, los muestreos coincidieron con el periodo reproductivo de Aegla, entre otoño y primavera. En esta etapa, las hembras adultas no se alimentan y pasan la mayor parte del tiempo enterradas de modo críptico entre las piedras, en la tentativa de buscar hábitats protegidos, registrándose densidades entre 8.7 a 19 individuos/m2, con máximos en invierno (Bueno y Bond-Buckup, 2000; Swiech-Ayoub y Masunari, 2001b), siendo presas fáciles para L. longicaudis.
La importancia relativa de los invertebrados constituye la principal diferencia en la dieta de la población del noroeste respecto a las otras conocidas del nordeste argentino. Sólo en un estudio en Brasil los crustáceos fueron más frecuentes que los peces (Uchôa et al., 2004).
Nuestros datos fueron similares a los documentados por Gori et al. (2003). En ambas localidades de la cuenca del Paraná existe similitud, ya que L. longicaudis consume en mayor cantidad peces, insectos y crustáceos. Sin embargo, difieren de los documentados por Parera (1992) para la laguna Iberá. Los datos de este último estudio muestran mayor similitud con los reportados para el río Iguazú (Parera, 1993b), localidad donde se registró una alta proporción de peces, seguidos por crustáceos, reptiles y mamíferos.
El patrón de similitud detectado no es explicado totalmente por las diferencias ambientales y ecológicas de las localidades comparadas (e. g. tipos de ambientes acuáticos, variaciones hidrológicas, disponibilidad de presas). De hecho, las poblaciones con mayor similitud se encuentran a más de 800 km, en condiciones ambientales y ecológicas muy diferentes (e.g. pluviosidad, cobertura vegetal, composición de especies, etc.). Por el contrario, estudios realizados en la misma localidad (Parera, 1992; Gori et al., 2003), y por tanto en condiciones ambientales y ecológicas similares, presentaron baja similitud.
El patrón observado puede ser explicado por dos aspectos, uno teórico y otro metodológico. La explicación teórica estaría dada por la adaptabilidad trófica de L. longicaudis, lo cual le permitiría explotar el recurso alimentario, cambiando sus hábitos de alimentación en respuesta a variaciones temporales y espaciales en la oferta de los recursos (Parera, 1992, 1993b; Gerking, 1994; Pardini, 1998; Colares y Waldemarin, 2000; Gori et al., 2003; Uchôa et al., 2004; Begon et al., 2005). Las razones metodológicas serían fundamentalmente los sesgos en las determinaciones o estimaciones de la representatividad de presas, que provocaron que los trabajos de un mismo autor (Parera, 1992, 1993b) sean más similares entre sí respecto a los realizados por otros (Gori et al., 2003 y este estudio). El nivel de precisión en las determinaciones taxonómicas puede haber influenciado en los resultados obtenidos por los distintos autores, presumiblemente a partir de los listados de ítems presas y sus frecuencias relativas. Por la técnica utilizada, también pueden haber existido otros sesgos, tales como el registro de moluscos que podría haberse subestimado, debido a que los lobitos de río suelen abandonar las caparazones, por lo que resulta poco probable hallarlos en las heces, excepto ingesta ocasional o de ejemplares muy pequeños (Pardini, 1998; Gori et al., 2003). Por último, el análisis de los datos aplicado en cada estudio podría constituir un factor de distorsión considerable.
En resumen, L. longicaudis es una especie generalista, presenta un amplio nicho trófico y responde funcionalmente a la fluctuación de sus presas. Probablemente manifieste una respuesta numérica del consumo a la disponibilidad de las presas, por lo que serían necesarios estudios sobre oferta alimentaria y variaciones estacionales de las mismas.
AGRADECIMIENTOS
Al personal del Instituto Nacional de Limnología (CONICET-UNL) y del Parque Nacional El Rey. Al Nodo NOA del SIB (Administración de Parques Nacionales) por proveer el mapa. A los especialistas que colaboraron en la identificación de algunas presas: G.M. de Gonzo, M. Rodriguez, L. Rossi, C. Noriega, G.P. de Amsler, P. Collins y V. Williner. A N. Pla por su asistencia en los análisis estadísticos.
LITERATURA CITADA
1. APN-SIB. 2008. Administración de Parques Nacionales. Sistema de Información de Biodiversidad. Proyecto de Conservación de la Biodiversidad - Donación GEF-BIRF TF 028372-AR. http://www.sib.gov.ar [ Links ]
2. ARAUJO LIMA CAM, AA AGOSTINHO y NN FABRÉ. 1995. Trophic aspects of fish communities in Brazilian Rivers and Reservoirs. Pp. 105-281, en: JG Tundisi, CE Bicudo y T Matsumura Tundisi (eds.) Limnology in Brazil, ABC/SBL, Brasil. [ Links ]
3. BAYLEY PB. 1973. Studies on the migratory characin, Prochilodus platensis Holmberg 1889, (Pisces, Characoidei) in the River Pilcomayo, South America. Journal of Fish Biology 5:25-40. [ Links ]
4. BEGON M, JL HARPER y CR TOWNSEND. 2005. Ecología. Individuos, poblaciones y comunidades. Ediciones Omega, Barcelona. [ Links ]
5. BUENO AA y G BOND-BUCKUP. 2000. Dinâmica populacional de Aegla platensis Schmitt (Crustacea, Decapoda, Aeglidae). Revista Brasileira de Zoologia 17:43-49. [ Links ]
6. BUENO AA y JC MOTTA-JUNIOR. 2006. Small mammal selection and functional response in the diet of the maned wolf, Chrysocyon brachyurus (Mammalia: Canidae), in Southeast Brazil. Mastozoología Neotropical 13:11-19. [ Links ]
7. BURKART R, N BÁRBARO, R SANCHEZ y D GÓMEZ. 1999. Eco-regiones de la Argentina. APNPRODIA. [ Links ]
8. CABRERA A y J YEPES. 1940. Mamíferos sudamericanos. Vida, costumbres y descripción. Compañía Argentina de Editores, Argentina. [ Links ]
9. CABRERA A. 1976. Regiones fitogeográficas argentinas. Pp. 1-85, en: Enciclopedia Argentina de Agricultura y Jardinería (WF Kugler, ed.). II, ACME, Argentina. [ Links ]
10. CANZIANI G, C ROSSI, S LOISELLE y R FERRATI (eds.) 2003. Los Esteros del Iberá. Informe del Proyecto "El manejo sustentable de humedales en el MERCOSUR". Fundación Vida Silvestre Argentina, Buenos Aires, Argentina. [ Links ]
11. CARSS D y S PARKINSON. 1996. Errors associated with otter Lutra lutra faecal analysis. I. Assesing general diet from spraints. Journal of Zoological London (Reino Unido) 238:301-317. [ Links ]
12. CASCIOTTA J, A ALMIRON y J BECHARA. 2005. Peces del Iberá. Hábitat y diversidad. Graficar, La Plata, Argentina. [ Links ]
13. COLARES EP y HF WALDEMARIN. 2000. Feeding of the Neotropical river otter (Lontra longicaudis) in the coastal region of the Rio Grande do Sul State, Southern Brazil. IUCN Otter Specialist Group Bulletin 17(1):6-13. [ Links ]
14. COLE GA. 1988. Manual de Limnología. Editorial Hemisferio Sur SA, Argentina. [ Links ]
15. CORBETT LK. 1989. Assessing the diet of dingoes from feces: a comparison of three methods. Journal of Wildlife Management 53:343-346. [ Links ]
16. COTE D, HMJ STEWART, RS GREGORY, J GOSSE, JJ REYNOLDS, GB STENSON y EH MILLER. 2008. Prey selection by marine-coastal river otters (Lontra canadensis) in Newfoundland, Canadá. Journal of Mammalogy 89:1001-1011. [ Links ]
17. COZAR A, CM GARCÍA y JA GÁLVEZ. 2003. Limnología de las lagunas Iberá y Galarza. Pp. 117-142, en: Los Esteros del Iberá. Informe del Proyecto "El manejo sustentable de humedales en el MERCOSUR" (G Canziani, C Rossi, S Loiselle y R Ferrati (eds.). Fundación Vida Silvestre Argentina, Buenos Aires, Argentina. [ Links ]
18. DÍAZ GB y RA OJEDA (eds.). 2000. Libro rojo de los mamíferos amenazados de la Argentina. Sociedad Argentina para el Estudio de los Mamíferos, Argentina. [ Links ]
19. DRAGO E y A PAIRA. 2001. Limnología física. Pp. 3-11, en: Evaluación de impacto en el río Iguazú en relación al derrame de petróleo en territorio brasilero. Tercer informe (muestreo 6 al 10 de agosto 2001) y Consideraciones finales (MJ Parma de Croux, coord.). Informe Inédito INALI, Santa Fe, Argentina. [ Links ]
20. EFRON B. 1979. Bootstrap methods: Another look at the jackknife. Annals of Statistics 7:1-26. [ Links ]
21. GALVIS G, J MOJICA y M CAMARGO. 1997. Peces del Catatumbo. Asociación Cravo Norte, Colombia. [ Links ]
22. GARCÍA-PEREA R, R PEREA, A BAQUERO y R FERNÁNDEZ-SALVADOR. 1996. Carnívoros: evolución, ecología y conservación. CSIC Museo de Ciencias Naturales, Sociedad Española para la conservación y estudio de los mamíferos, España. [ Links ]
23. GERKING SD. 1994. Feeding ecology of Fish. Academic Press, Estados Unidos. [ Links ]
24. GIL G. 1998. Distribution and monitoring of the Neotropical Otter in the El Rey National Park, Salta Province, Argentina. IUCN Otter Specialist Group Bulletin 15(1):56. [ Links ]
25. GIRI F y V WILLINER. 2006. Arreglo espacial del cangrejo dulceacuícola Aegla affinis (Crustacea: Decapoda: Anomura) en un ambiente léntico de altura. 22 Reunión Argentina de Ecología. Córdoba, Argentina. Libro de Resúmenes:307. [ Links ]
26. GONZO GM. 2003. Peces de los ríos Bermejo, Juramento y Cuencas Endorreicas. Museo de Ciencias Naturales y Consejo de Investigaciones de la Universidad Nacional de Salta, Argentina. [ Links ]
27. GORI M, GM CARPANETO y P OTTINO. 2003. Spatial distribution and diet of the Neotropical otter Lontra longicaudis in the Iberá Lake (northern Argentina). Acta Theriologica 48:495-504. [ Links ]
28. HELDER DA. 1997. Food and feeding habits of the Neotropical river otter Lontra longicaudis (Carnivora, Mustelidae). Mammalia 61:193-203. [ Links ]
29. IUCN. 2006. 2006 IUCN Red List of Threatened Species. http://www.iucnredlist.org [ Links ]
30. JACOBSEN L y HM HANSEN. 1996. Analysis of otter (Lutra lutra) spraints: Part 1: Comparison of methods to estimate prey proportions; Part 2: Estimation of the size of prey fish. Journal of Zoological London (Reino Unido) 238:167-180. [ Links ]
31. KASPER CB, MJ FELDENS, J SALVI y C ZANARDI GRILLO. 2004. Estudo preliminar sobre a ecologia de Lontra longicaudis (Olfers) (Carnivora, Mustelidae) no Vale do Taquari, Sul do Brasil. Revista Brasileira de Zoologia 21:65-72. [ Links ]
32. LARIVIERE S. 1999. Lontra longicaudis. Mammalian Species 609:1-5. [ Links ]
33. LAROCHE JL. 1982. Trophic patters among larvae of fish species of sculpins (Family: Cottidae) in marine stuary. Fishery Bulletin, Seattle (Estados Unidos) 80:827-840. [ Links ]
34. LÓPEZ HL, CC MORGAN y MJ MONTENEGRO. 2002. Ichthyological Ecoregions of Argentina. ProBiota (FCNyM, UNLP), Serie Documentos nº 1. On line version, http://aplicaciones.medioambiente.gov.ar/archivos/web/GTRA/File/ichthyological%20ecoregions.pdf [ Links ]
35. LOUZADA-SILVA D, T MARTINS VIEIRA, J PINHO DE CARVALHO, A PUCCI HERCOS y B MERGULHÃO DE SOUZA. 2003. Uso de espaço e de alimento por Lontra longicaudis no Lago Paranoá, Brasília, DF. Universitas Ciências da Saúde (Brasil) 1(2):305-316. [ Links ]
36. LOWE-MC CONNELL RH. 1987. Ecological Studies in Tropical Fish Communities. Cambridge University Press, Cambridge. [ Links ]
37. MERRITT RW y KW CUMMINS. 1996. An introduction to the aquatic insects of North America. Kendall/Hunt Pub. Dubuque, Iowa, Estados Unidos. [ Links ]
38. MUANIS MC y HF WALDEMARIN. 2003. Dieta da lontra (Lontra longicaudis) e da ariranha (Pteronura brasiliensis) no Rio Negro, Pantanal, MS. Pp. 55-57, en: Anais de Trabalhos Completos do VI Congresso de Ecologia do Brasil, Fortaleza, Brasil (V de Claudino-Sales, IM Tonini y EW Correia-Dantas, eds.). Editora da Universidade Federal do Ceará, Brasil. [ Links ]
39. ODUM EP y GW WARRETT. 2006. Fundamentos de Ecología, 5º Edición. Thomson, México. [ Links ]
40. PARDINI R. 1998. Feeding ecology of the Neotropical river otter Lontra longicaudis in an Atlantic Forest stream, south-eastern. Journal of Zoology 245:385-391. [ Links ]
41. PARERA A. 1992. Análisis de la dieta de Lutra longicaudis en Laguna Iberá, Provincia de Corrientes, Argentina. 5 Reunión de Especialistas en Mamíferos Acuáticos de América del Sur. Libro de Resúmenes. Buenos Aires, Argentina: 49. [ Links ]
42. PARERA A. 1993a. Alimentación del lobito de río Lontra longicaudis (Olfers, 1818) en cautiverio. Aspectos inherentes a los hábitos de forrajeo y estimación de dieta por análisis de fecas. Seminario de Licenciatura, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina. [ Links ]
43. PARERA A. 1993b. Dieta de Lutra longicaudis en el Río Iguazú, Misiones, Argentina. Póster presentado en XVI Reunión Argentina en Ecología. Puerto Madryn. Argentina: 8. [ Links ]
44. QUADROS J y ELA MONTEIRO FILHO. 2000. Fruit ocurrence in the diet of the Neotropical otter Lontra longicaudis, in Southern Brazilian Atlantic Forest and its implication for seed dispersion. Mastozoología Neotropical 7:33-36. [ Links ]
45. QUADROS J y ELA MONTEIRO FILHO. 2001. Diet of the Neotropical Otter, Lontra longicaudis, in an Atlantic Forest Area, Santa Catarina State, Southern Brazil. Studies on Neotropical Fauna & Environment 36:15-21. [ Links ]
46. QUIRÓS R. 1987. Abundancia de peces en relación con la materia orgánica, en la cuenca del Plata (Sudamérica). Ecología 8:91-109. [ Links ]
47. REYNOLDS JC y NJ AEBISCHER. 1991. Comparison and quantification of carnivore diet by faecal analysis: a critique, with recommendations, based on a study of the Fox Vulpes vulpes. Mammalian Review 21:97-122. [ Links ]
48. RINGUELET RA. 1975. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2(3):1-122. [ Links ]
49. ROSENZVAIG AL. 1974. Contribución al conocimiento de la biología del lobito de río (Lutra platensis Waterhouse, 1839). Instituto para la Investigación de los problemas del mar. Mar del Plata, Argentina. [ Links ]
50. RUIZ OLMO J, G JORDÁN y J GOSALBEZ. 1989. Alimentación de la nutria (Lutra lutra L., 1758) en el Nordeste de la Península Ibérica. Doñana Acta Vertebrata 16:227-237. [ Links ]
51. SERVICIO METEOROLÓGICO NACIONAL. 2008. Climatología. Datos Estadísticos. http://www.smn.gov.ar/?mod=clima%26id=30%26provincia=Misiones%26ciudad=Iguazú [ Links ]
52. SIEGEL S. 1983. Estadística no paramétrica aplicada a las ciencias de la conducta. Editorial Trillas, México. [ Links ]
53. SUBSECRETARÍA DE RECURSOS HÍDRICOS. 2008. Sistema Nacional de Información Hídrica / Red Básica. Descripción de Cuencas Hídricas. Red hidrológica nacional. Estadística hidrológica 2004. Estaciones El Piquete (Cód. 0618) y Puerto Andresito (Cód. 3448). http://www.hidricosargentina.gov.ar/estad2004/sus-0618.htm. http://www.hidricosargentina.gov.ar/estad2004/act-3448.htm [ Links ]
54. SWIECH-AYOUB BP y S MASUNARI. 2001a. Flutuações temporal e espacial de abundância e composição de tamanho de Aegla castro Schmitt (Crustacea, Anomura, Aeglidae) no Buraco do Padre, Ponta Grossa, Paraná, Brasil. Revista Brasileira de Zoologia 18:1003-1017. [ Links ]
55. SWIECH-AYOUB BP y S MASUNARI. 2001b. Biologia reproductiva de Aegla castro Schmitt (Crustacea, Anomura, Aeglidae) no Buraco do Padre, Ponta Grossa, Paraná, Brasil. Revista Brasileira de Zoologia 18:1019-1030. [ Links ]
56. UCHÔA T, GP VIDOLIN, TM FERNANDES, GO VELASTIN y PR MANGINI. 2004. Aspectos ecológicos e sanitários da lontra (Lontra longicaudis Olfers, 1818) na Reserva Natural Salto Morato, Guaraqueçaba, Paraná, Brasil. Cadernos da Biodiversidade 4:19-28. [ Links ]
57. WILSON DE y DM REEDER (eds.). 1993. Mammals Species of the World. Smithsonian Institution Press, Estados Unidos. [ Links ]
Recibido 3 mayo 2007.
Aceptado 11 marzo 2009.
Editor asociado: J Polop

