










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkMastozoología neotropical
Print version ISSN 0327-9383On-line version ISSN 1666-0536
Mastozool. neotrop. vol.17 no.1 Mendoza Jan./June 2010
ARTÍCULOS Y NOTAS
Mamíferos del pedemonte de Yungas de la Alta Cuenca del Río Bermejo en Argentina: una línea de base de diversidad
J. Pablo Jayat1 y P. E. Ortiz2
1 Laboratorio de Investigaciones Ecológicas de las Yungas (LIEY), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, Casilla de Correo 34, 4107 Yerba Buena, Tucumán, Argentina [Correspondencia: <pjayat@proyungas.org.ar>].
2 Cátedra de Paleontología, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, San Miguel de Tucumán, Tucumán, Argentina.
RESUMEN: Las Yungas albergan gran parte de la biodiversidad presente en Argentina, siendo la Alta Cuenca del Río Bermejo (ACRB) un área prioritaria de conservación. Numerosos mamíferos de Yungas han sufrido retracciones de su distribución por lo que el conocimiento sobre diversidad y estatus de sus poblaciones es imprescindible. Se reportan resultados de inventarios para 22 localidades de pedemonte de la ACRB. Se describe la composición de especies para todos los grupos y, para micromamíferos, la riqueza, dominancia y abundancia relativa. Se registraron 74 especies de mamíferos para la ACRB. La riqueza y dominancia fueron 27.5 (estimador Chao 1, S*) y 0.47 (índice de Simpson, D) para quirópteros, y 17 (S*) y 0.24 (D) para micromamíferos terrestres. Sturnira lilium, Akodon simulator y Euryoryzomys legatus fueron notablemente más abundantes que el resto de las especies. Diez especies se capturaron sólo en la temporada seca y ocho durante la húmeda. La riqueza y dominancia para ambos grupos fue prácticamente la misma al comparar entre estaciones. Para quirópteros se observó una especie (Lasiurus blossevillii) con una mayor abundancia relativa durante la temporada húmeda pero no se registraron diferencias para micromamíferos terrestres.
ABSTRACT: Mammals of the piedmont of Yungas in the Alta Cuenca del Río Bermejo in Argentina: A diversity base-line. Yungas forests harbor a great part of the biodiversity in Argentina, being the Alta Cuenca del Río Bermejo (ACRB) a high-priority area for conservation. Because many of Yungas mammals have suffered retractions of their distribution, studies about their diversity and population status are essential. We report results of rapid biodiversity assessments for 22 localities of Yungas premontane forest of the ACRB. We describe the community composition for all taxonomic groups of mammals and we estimated, for micromammals, species richness, dominance and relative abundance. We documented 74 species of mammals for the ACRB. The species richness and dominance were 27.5 (Chao 1 estimator, S*) and 0.47 (Simpson index, D) for bats, and 17 (S*) and 0.24 (D) for non volant mammals. Sturnira lilium, Akodon simulator and Euryoryzomys legatus were notably more abundant than other species. Ten species were caught exclusively in the dry season and eight during the wet season. Species richness and dominance for both species groups were very similar among seasons. For bats we observed one species (Lasiurus blossevillii) with a higher relative abundance during the humid season but we do not register differences for non volant micromammals.
Key words. Environmental baseline; Northwestern Argentina; Small mammals; Species evenness; Species richness; Yungas.
Palabras clave. Línea de base ambiental; Noroeste argentino; Pequeños mamíferos; Riqueza de especies; Equitatividad de especies; Yungas.
INTRODUCCIÓN
Los bosques húmedos subtropicales en Argentina, representados por las ecorregiones Paranaense y Yungas, constituyen menos del 2% de la superficie de Argentina continental pero contienen más del 50% de la biodiversidad presente en este país (Brown et al., 2001). Las Yungas argentinas, distribuidas desde la frontera con Bolivia hasta el sudeste de la provincia de Catamarca, ocupan una superficie estimada en 40 000 km2, con una extensión latitudinal de 600 km y menos de 100 km de ancho, y un rango altitudinal entre 400 y 3000 m (Brown et al., 2001). Latitudinalmente, como consecuencia de la disposición de los cordones montañosos sobre los que se desarrollan, las Yungas presentan una distribución discontinua e insular. Esta configuración genera condiciones para la coexistencia de una gran diversidad biológica, organizándola en pisos altitudinales (Selva Pedemontana, Selva Montana y Bosque Montano) y sectores latitudinales (Norte, Centro y Sur) de características claramente distinguibles. Los sectores pedemontanos situados al norte de la región son los que presentan una mayor diversidad biológica y cultural (Ojeda y Mares, 1989; Brown et al., 2001).
Diversos trabajos realizados en los últimos años han señalado a la Alta Cuenca del Río Bermejo (ACRB) como un área prioritaria de conservación en el noroeste de Argentina (e. g., Brown, 1995a; Brown et al., 2001). Esta región presenta la mayor superficie continua de selvas de montaña de nuestro país, posee importantes recursos forestales, numerosos humedales de biodiversidad sobresaliente y la concentración más importante de comunidades aborígenes y campesinas. Por todo ello, la ACRB ha sido considerada la única área capaz de garantizar a largo plazo la persistencia de la biodiversidad completa de la ecorregión.
Los mamíferos de las Yungas constituyen un grupo diverso con aproximadamente 130 especies registradas hasta el presente. Muchas de las especies de este grupo (en particular aquellas de mayor porte) han sufrido retracciones notables de su distribución debidas, principalmente, a la pérdida de su hábitat natural. Esta situación ha derivado en un estatus de conservación de riesgo para muchas de las especies de este ambiente (Díaz y Ojeda, 2000). Las especies pequeñas (mayormente roedores, quirópteros y marsupiales), aunque potencialmente menos afectadas por las actividades humanas, constituyen la mayor parte de la diversidad del grupo y son ampliamente desconocidas en aspectos elementales de su biología tales como taxonomía y distribución. Las adiciones de especies de pequeños mamíferos a la fauna local y los debates sobre su estatus taxonómico son frecuentes (e. g., Barquez et al., 1999a, 2006a y b; Díaz et al., 1999, 2002; Jayat y Miotti, 2005; Jayat et al., 2006, 2007, 2008a). En este contexto, los estudios sobre diversidad y el estatus de sus poblaciones en la región son prioritarios ya que constituyen una herramienta primordial para su manejo y conservación.
Inventarios rápidos de diversidad de mamíferos realizados en los últimos años en numerosas localidades de la ACRB han permitido generar una línea de base sobre riqueza y abundancia relativa de las especies de este grupo. Este conocimiento puede constituir un punto de partida para el desarrollo de sistemas de monitoreo de los cambios producidos en las comunidades de mamíferos y un insumo más de los procesos de planificación territorial en la región. En este trabajo se presentan los resultados obtenidos hasta el momento en estos inventarios.
ANTECEDENTES
Aunque se han llevado a cabo numerosos estudios mastozoológicos en el noroeste argentino, en la actualidad no existen aproximaciones detalladas que permitan caracterizar la comunidad de mamíferos de las Yungas. Así, aunque algunos trabajos se han centrado en grupos taxonómicos particulares (e. g., Barquez y Díaz, 2001; Perovic, 2002; Jayat et al., 2008b) o han caracterizado algunos sectores de la ecorregión (e. g., Ojeda y Mares, 1989; Jayat et al., 2008b, 2009), los registros que documentan la riqueza e identidad taxonómica de su mastofauna corresponden, en general, a menciones ocasionales. De este modo, mucha de la información disponible en la actualidad es antigua (e. g., Allen, 1901; Thomas, 1918, 1921; Yepes, 1935), está contenida en revisiones taxonómicas a nivel continental (e. g., Pearson, 1958; Hershkovitz, 1962; Olds, 1988; Myers, 1989; Myers et al., 1990), en trabajos que documentan de un modo general la mastofauna de amplias regiones o áreas protegidas del país (e. g., Olrog, 1979; Ojeda y Mares, 1989; Heinonen y Bosso, 1994; Mares et al., 1997; Barquez et al., 1999b; Contreras, 1999; Jayat et al., 1999, 2006; Díaz et al., 2000; Perovic, 2002; Gil y Heinonen Fortabat, 2003; Díaz y Barquez, 2007; Flores et al., 2007) y en publicaciones referidas a otros tópicos (e. g., Bianchi et al., 1971; Brown et al., 1986; Varela y Brown, 1995; Autino et al., 1999; Giannini, 1999; Ortiz y Pardiñas, 2001). En consecuencia, gran parte de la información se encuentra dispersa, es incompleta, y no ha sido aún críticamente evaluada.
Las áreas con mayor información dentro de las Yungas son los sectores pedemontanos. Sin embargo, no existe aún un consenso acerca de las especies de mamíferos incluidas en este piso altitudinal. Jayat et al. (2009) refieren una lista de 137 especies con al menos una mención para este sector. Aunque este número representa algo más del 35% de las especies de mamíferos de Argentina, siete taxones fueron considerados sólo de presencia histórica en el área y 24 tratados como menciones que deben ser adecuadamente documentadas. De las 106 especies consideradas de presencia actual comprobada, el 80% estuvo representado por quirópteros (35%), roedores (29%) y carnívoros (15%). De acuerdo a Jayat et al. (2009), las áreas de selva pedemontana están entre las mejor relevadas del noroeste argentino, con algo más de 340 localidades referidas. Sin embargo, la mayoría de las localidades se encuentran agrupadas en el sector norte de la región y un 76% presentan cinco o menos especies registradas. Además, la mayor parte de las especies cuenta con escasos registros, con un 70% de aquellas con presencia comprobada conocidas sólo para diez o menos localidades (véase también Ojeda et al., 2003). El sector con mayor diversidad de mamíferos en el pedemonte de Argentina es el ubicado en la ACRB (Brown et al., 2001; Ojeda et al., 2008). Jayat et al. (2009) mencionan 97 especies para esta área, 14 de ellas exclusivas. A pesar de estos aportes, no se han realizado hasta el momento estudios que enfoquen la diversidad de mamíferos del pedemonte de Yungas desde un punto de vista cuantitativo y, por lo tanto, no existen estimaciones confiables y comparativamente útiles sobre riqueza, abundancia relativa y patrones de dominancia de las distintas especies.
MATERIALES Y MÉTODOS
Área de estudio y sitios de muestreo
El área de estudio de esta contribución se centra en el pedemonte de la ACRB. Debido a la dominancia de Calycophyllum multiflorum y Phyllostilon rhamnoides en este sector, este piso altitudinal es conocido también como "selva de palo blanco y palo amarillo". La selva pedemontana en la ACRB se distribuye altitudinalmente entre 400 y 900 m y presenta una superficie superior a 5000 km2. Algunas especies de árboles también dominantes en elárea incluyen el lapacho rosado (Tabebuia impetiginosa), cedro rosado (Cedrela balansae), roble (Amburana cearensis), cebil colorado (Anadenanthera colubrina), quina (Myroxylon peruiferum), afata (Cordia tricótoma), palo lanza (Patagonula americana) y urundel (Astronium urundeuva) (Brown, 1995b).
En el presente estudio se analiza la información sobre diversidad de mamíferos para 22 localidades inventariadas entre mayo del año 2003 y marzo de 2008 (Fig. 1 y Apéndice). Diez de las localidades relevadas se encuentran situadas al noreste de la ACRB, en los alrededores de las ciudades de Tartagal y General Ballivián. Este sector de la alta cuenca se encuentra aislado de la Cordillera Oriental por los ríos Itaú, Grande de Tarija y Bermejo y está vinculado más estrechamente a zonas chaqueñas que a otros pisos altitudinales de Yungas. Al norte limita con áreas pedemontanas transicionales de Yungas y Chaco de Bolivia. El pedemonte alcanza en esta región 900 m de altitud y muestra características particulares, con algunas especies de árboles (e. g., Caesalpinea floribunda, Capparis sp., Tartagalia roseorum) no registradas en otros sectores. Las áreas de selva montana están reducidas aquí a pequeños parches aislados, ubicados en quebradas húmedas por encima de 900 m en áreas cumbrales de la Sierra de Tartagal. Los inventarios realizados en esta zona incluyen 50 noches de muestreo, ocho de ellas en otoño, 17 en invierno, 12 en primavera y 13 en verano. Cuatro localidades están situadas en la zona central del área de estudio, en los alrededores de la ciudad de Orán. El área se encuentra estrechamente vinculada a la Cordillera Oriental y la selva pedemontana llega sólo a los 700 m, limitando ampliamente con la selva montana de Argentina y Bolivia. Hacia el Este limita con áreas de pedemonte altamente perturbado y ambientes de chaco serrano. Las localidades estudiadas en esta región se encuentran separadas de aquellas del sector noreste por el curso superior del río Bermejo. En esta área se realizaron 19 noches de muestreo, cinco en primavera y 14 en verano. Las restantes ocho localidades de estudio se encuentran al Sur, cinco en los alrededores de la ciudad de Libertador General San Martín y tres en las cercanías de El Piquete, al pie de la Sierra de Santa Bárbara. Este sector está relacionado estrechamente con la Cordillera Oriental y el sistema de Sierras Subandinas. Al igual que en la región anterior el pedemonte en esta área no supera los 700 m y se dispone en amplio ecotono con la selva montana hacia el Oeste y con bosques pedemontanos perturbados y de chaco serrano del este de Jujuy y centro-oeste de Salta. Se encuentra, además, separado de los restantes sectores por los ríos Colorado y San Francisco, entre otros. El río San Francisco y áreas de chaco serrano separan a las localidades situadas en los alrededores de Libertador General San Martín de aquellas asociadas a la Sierra de Santa Bárbara. Los inventarios realizados en esta área incluyen 33 noches de muestreos, 16 de ellas en otoño, cuatro en invierno, cuatro en primavera y nueve en verano.

Fig. 1. Mapa del área de estudio y localidades de muestreo. Los números de las localidades corresponden a los del Apéndice.
Técnicas de muestreo y taxidermia
Las técnicas de muestreo se dividieron en tres categorías, relacionadas a cada grupo de mamíferos estudiado: 1) micromamíferos voladores, 2) micromamíferos no voladores y 3) mesomacromamíferos.
1) Micromamíferos voladores: para la captura de quirópteros se utilizaron redes de niebla dispuestas en transectas sobre cuerpos de agua, caminos y el interior del bosque. El esfuerzo de muestreo diario y total para este grupo se expresó en metros red/hora (m r/h).
2) Micromamíferos no voladores: los pequeños marsupiales y roedores se capturaron mediante transectas de trampas de captura viva tipo Sherman cebadas con avena. El esfuerzo de muestreo para este grupo se expresó como trampas noche (t/n). El diseño de transectas empleado en ambos casos maximiza la superficie relevada y aumenta así las probabilidades de registro de la riqueza de especies de una localidad. Este diseño optimiza, además, el inventario de la heterogeneidad ambiental y magnifica la probabilidad de detección de la riqueza total de especies presentes en un área determinada. Estas técnicas son las más apropiadas para estudios rápidos de diversidad de estos grupos, ya que permiten el examen detallado, la colecta o liberación de los especímenes, su identificación y la cuantificación de la abundancia relativa en períodos cortos de tiempo.
3) Meso-macromamíferos: los mamíferos de mediano y gran porte (especies de 500 g o más) fueron registrados mediante observaciones directas, encuestas a pobladores locales y la observación de huellas y otros signos de actividad. En cada localidad se realizaron un mínimo de dos encuestas a pobladores de la zona. Sólo se realizaron identificaciones a nivel de especie a partir de esta última técnica cuando los pobladores pudieron aportar un conjunto de caracteres "diagnósticos" o que inequívocamente pudieron ser asignados a las mismas.
La identificación de los especímenes se realizó con la ayuda de fuentes bibliográficas y mediante comparación con material de referencia depositado en las siguientes colecciones: Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (MACN), Buenos Aires; Museo Municipal de Ciencias Naturales "Lorenzo Scaglia" (MMPMa), Mar del Plata, Buenos Aires; Colección de Mamíferos Lillo (CML), San Miguel de Tucumán, Tucumán; Museo de Ciencias Naturales de Salta (MCNS), Salta, y la Colección Mastozoológica del IADIZA (CMI), Mendoza.
Debido a las dificultades para identificar en el campo algunos especímenes de pequeños mamíferos a partir de sus caracteres externos, se colectaron ejemplares de referencia para su estudio posterior en gabinete. Las técnicas de taxidermia siguieron los lineamientos básicos de Mares et al. (1989) y Díaz et al. (1998). Todos los especímenes colectados están registrados en el catálogo personal de uno de los autores (JPJ) y serán depositados en el MACN.
Análisis de datos
A partir de la revisión de literatura y registros de colecciones se realizó una descripción de la composición de especies potencialmente presentes en el área de estudio. Los fundamentos para la inclusión o no de las distintas especies siguen los criterios de Jayat et al. (2009).
Se realizó también una descripción de la composición de especies registrada durante los inventarios para toda el área de estudio y para las temporadas seca y húmeda. El noroeste argentino presenta un régimen de precipitaciones cuasimonzónica (Minetti et al., 2005), con la mayor parte de las lluvias concentradas entre fines de la primavera y principios de otoño. Por ello, consideramos como estación húmeda al período comprendido entre los meses de noviembre y abril, y seca al comprendido entre mayo y octubre. Para cada especie registrada se detallan las localidades y los tipos de registro obtenidos durante los trabajos de campo (Tabla 1).
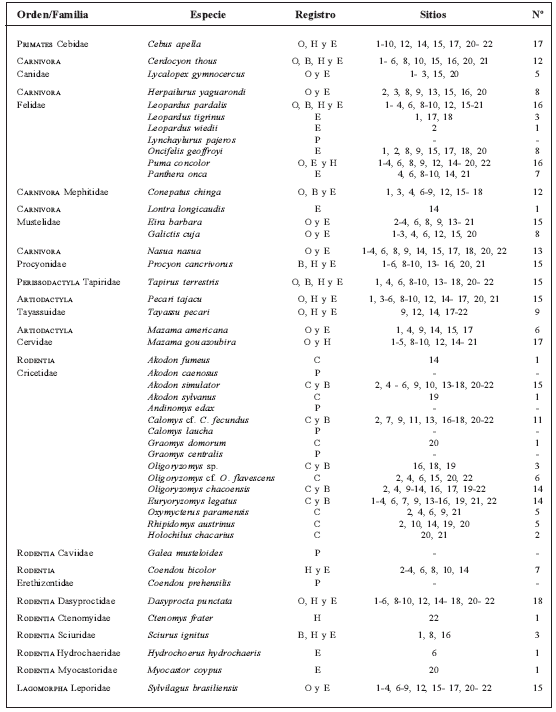
Tabla 1 Listado taxonómico de especies de mamíferos probables y registradas para el pedemonte de Yungas de la ACRB. El ordenamiento taxonómico sigue los criterios de Barquez et al. (2006a) con modificaciones. B = bibliografía; C = captura; E = encuesta; P = probable; O = observada; H = huellas, signos de actividad o restos; Nº = número total de localidades.

Para pequeños mamíferos se llevó a cabo, además, un análisis cuantitativo de la riqueza, dominancia y abundancia relativa de las especies de la comunidad. La riqueza se expresó mediante el número de especies registradas o riqueza observada (S) y el estimador Chao 1 (S*; Chao, 1987; Colwell y Coddington, 1994). Esta última técnica se basa en el número de especies raras y constituye un estimador simple de la riqueza de un ensamble de especies. Sus cualidades no paramétricas lo hacen especialmente apropiado para el tipo de datos obtenidos en inventarios rápidos y su frecuente utilización en estudios de diversidad de mamíferos en los últimos años facilitan su comparación (e. g., Simmon y Voss, 1998; Voss et al., 2001; Sánchez et al., 2008). El componente de equitatividad se expresó mediante el índice de Simpson (D) y mediante gráficas de la distribución de abundancias de las especies (proporción de individuos de cada especie respecto del total de individuos). El índice de Simpson ha sido propuesto como una medida de dominancia más que de diversidad debido al peso que otorga a las abundancias de las especies más comunes en la muestra. Este índice presenta, además, una baja sensibilidad a diferentes tamaños de muestra y ha sido ampliamente utilizado en estudios de biodiversidad (Magurran, 1998). La abundancia relativa para pequeños roedores y marsupiales se calculó como el número de individuos capturados cada 100 trampas noche (N° de individuos/100 t/n) mientras que para quirópteros este valor se expresa como el número de individuos capturados por cada 100 metros red por hora de muestreo (Nº de individuos/100 m r/h). La significancia de las diferencias en abundancia relativa para cada especie entre temporadas se evaluó mediante el test no paramétrico de Kruskal-Wallis para un p < 0.05.
RESULTADOS
ACRB
Durante los inventarios realizados se documentaron 74 de las 97 especies consideradas de presencia actual comprobada por Jayat et al. (2009) para el pedemonte de Yungas de la ACRB (Tabla 1). Estas especies están agrupadas en diez órdenes y 25 familias. Se registraron además dos formas aún no determinadas del género Myotis y una de Oligoryzomys. Quirópteros, roedores y carnívoros fueron los grupos taxonómicos mejor representados durante los estudios de campo, acumulando en conjunto el 78% de las especies registradas, con 25, 18 y 15 especies respectivamente. El 22% restante estuvo representado por seis marsupiales, seis xenartros, cuatro artiodáctilos, un primate, un perisodáctilo y un lagomorfo (Tabla 1).
Una especie de quiróptero (Sturnira lilium) fue la más frecuentemente registrada en nuestros relevamientos. Esta especie fue documentada en 20 de las 22 localidades estudiadas. Otras especies frecuentes en nuestros inventarios incluyeron Dasyprocta punctata, (18 localidades), Cebus apella, Mazama gouazoubira (17 localidades), Leopardus pardalis, Puma concolor (16 localidades), Eira barbara, Procyon cancrivorus, Tapirus terrestris, Pecari tajacu, Akodon simulator, Sylvilagus brasiliensis (15 localidades), Oligoryzomys chacoensis, Euryoryzomys legatus (14 localidades), Nasua nasua (13 localidades), Artibeus planirostris, Cerdocyon thous, Conepatus chinga (12 localidades) Euphractus sexcinctus, Eptesicus furinalis y Calomys cf. C. fecundus (11 localidades). En contraposición, se han registrado 36 especies de mamíferos para menos de cinco localidades (Tabla 1). Entre éstas hay dos especies que hasta el momento parecen estar restringidas a un sector geográfico particular de la ACRB. Monodelphis kunsi es una especie registrada en sólo cuatro localidades (y por apenas cinco individuos) en el pedemonte del sector noreste, en los alrededores de Tartagal, mientras que Akodon sylvanus cuenta con escasas localidades de registro en el extremo sur de la ACRB (sólo uno de ellos en áreas pedemontanas), en los alrededores de la Sierra de Santa Bárbara.
Se capturaron un total de 1484 especímenes de micromamíferos, 955 ejemplares pertenecieron a quirópteros y 529 a pequeños roedores y marsupiales. Estos ejemplares se agruparon en 41 especies, 25 de quirópteros y 16 de micromamíferos no voladores. Las curvas acumulativas de especies obtenidas para ambos grupos muestran una tasa de adición de nuevas especies decreciente con el aumento de localidades relevadas pero no muestran evidencias de haber alcanzado un valor asintótico (Fig. 2). Los resultados obtenidos en este estudio indican, además, un inventario más completo en el caso de los micromamíferos no voladores. Para este grupo se observó un mismo valor de riqueza observada (S) y esperada (S*) (Tabla 2) y las curvas acumulativas de especies presentaron una tasa de adición de nuevas especies algo menor (Fig. 2).
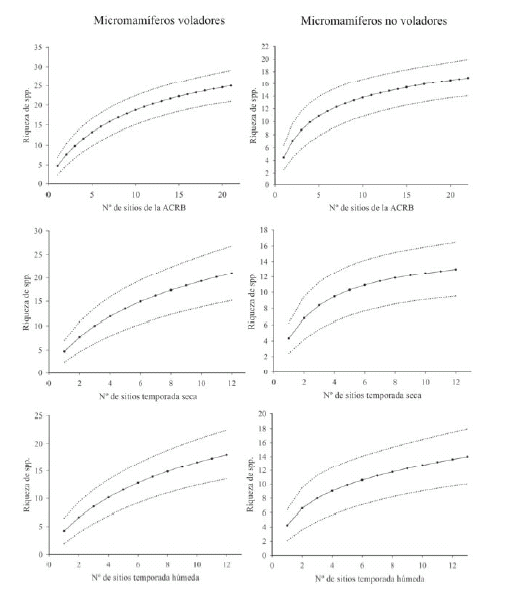
Fig. 2. Curva acumulativa de especies de quirópteros y micromamíferos no voladores por el número de localidades relevadas para toda la ACRB y las estaciones seca y húmeda. Los valores medios de riqueza y los intervalos de confianza (95%) se obtuvieron mediante la fórmula analítica (Sobs.) desarrollada en Colwell et al. (2004).
Tabla 2 Esfuerzo de muestreo, número de individuos, valores de riqueza observada (S) y esperada por el estimador Chao 1 (± desvío estándar de la media) y dominancia estimada por el índice de Simpson (D) de las especies de micromamíferos registradas en la ACRB y para las estaciones seca y húmeda.

Las gráficas de distribución de abundancias para toda la ACRB muestran una especie de quiróptero (Sturnira lilium) y 2 especies de roedores sigmodontinos (Akodon simulator y Euryoryzomys legatus) ampliamente dominantes sobre el resto de los micromamíferos (Fig. 3). Casi el 70% de los ejemplares de quirópteros capturados correspondieron a S. lilium y una proporción similar estuvo representada por los individuos de A. simulator y E. legatus entre los micromamíferos no voladores. La dominancia por una sola especie en el caso de los quirópteros, en lugar de dos, como ocurrió en los micromamíferos no voladores, se refleja en un mayor índice de Simpson (Tabla 2).

Fig. 3. Gráfica de abundancia por especies de quirópteros y micromamíferos no voladores registradas para toda la ACRB y las estaciones seca y húmeda. La abundancia (eje y) se expresa como el porcentaje de individuos para cada una de las especies (eje x) respecto del total de individuos registrados. Los números corresponden a los de las especies de las Tablas 3 (quirópteros) y 4 (micromamíferos no voladores).
De acuerdo a los datos de abundancia relativa, sólo dos especies de quirópteros (S. lilium y S. erythromos) y tres sigmodontinos (A. simulator, E. legatus y Oligoryzomys sp.) presentan, en promedio, más de un individuo cada 100 mr/h y 100 t/n respectivamente. Otras especies relativamente abundantes incluyeron a Molossus molossus, Oligoryzomys chacoensis y Calomys cf. C. fecundus, todas con un promedio mayor a 0.7 individuos y un límite superior de rango mayor a un individuo cada 100 mr/h y 100 t/n (Tablas 3 y 4). Artibeus planirostris, Eptesicus furinalis, Myotis nigricans y Akodon fumeus son otras especies que fueron abundantes en algunas localidades, con límites superiores de rango iguales o mayores a un individuo, pero con promedios relativamente bajos para el área de la ACRB en general.
Tabla 3 Valores de abundancia relativa para las especies de quirópteros registradas en el pedemonte de Yungas de la ACRB. Se ofrecen promedio, rango y tamaño de la muestra. La abundacia relativa fue medida como el número de individuos cada 100 metros red por hora de muestreo. El asterisco indica la única especie con abundancia relativa significativamente diferente entre las estaciones seca y húmeda de acuerdo al test de Kruskal-Wallis para un p< 0.05.

Tabla 4 Valores de abundancia relativa para las especies de micromamíferos no voladores registradas en el pedemonte de Yungas de la ACRB. Se ofrecen promedio, rango y tamaño de la muestra. La abundancia relativa fue medida como número de individuos capturados cada 100 trampas noche.

Estaciones seca y húmeda
Durante los inventarios se registraron diez especies de micromamíferos exclusivamente en la temporada seca y ocho sólo durante la húmeda. La mayoría de las diferencias encontradas entre estaciones se debe a cambios en la composición de quirópteros, con siete especies sólo registradas en la estación seca (Noctilio leporinus, Pygoderma bilabiatum, Sturnira oporaphilum, Histiotus macrotus, Myotis albescens y Myotis sp. 1 y sp. 2) y cuatro sólo en la húmeda (Glossophaga soricina, Histiotus laephotis, Molossops teminckii y Tadarida brasiliensis). Además, se registraron tres sigmodontinos exclusivos para la estación seca (Akodon fumeus, A. sylvanus, Oligoryzomys sp.) y dos sigmodontinos y un marsupial para la estación húmeda (Holochilus chacarius, Graomys domorum, Thylamys cinderella).
La diversidad de especies documentada para ambos grupos de micromamíferos fue prácticamente la misma entre estaciones, aunque levemente mayor para la húmeda de acuerdo al estimador S*. En el caso de los quirópteros la riqueza observada de especies (S) no coincidió con la esperada (S*) y fue apenas menor en la estación húmeda (Tabla 2).
Tampoco se observaron diferencias notorias en la dominancia de ambos grupos de micromamíferos entre temporadas, aunque en ambos casos la equitatividad fue levemente mayor en la estación seca (Tabla 2).
Sólo una de las 24 especies de micromamíferos documentadas en ambas temporadas (Lasiurus blossevillii) fue significativamente más abundante durante la estación húmeda (Tabla 3). Para quirópteros, una vez más, Sturnira lilium fue la especie más abundante en ambas estaciones del año. Sturnira erythromos y Molossus molossus fueron otras dos formas con más de un individuo por cada 100 mr/h en las temporadas seca y húmeda respectivamente. Entre los micromamíferos no voladores Akodon fumeus, A. simulator, Euryoryzomys legatus y Calomys cf. C. fecundus presentaron más de un individuo por cada 100 t/n en la estación seca y A.simulator y E. legatus durante la húmeda.
DISCUSIÓN Y CONCLUSIONES
La urgente necesidad de estudios cuantitativos sobre la diversidad de especies de mamíferos (y otros grupos taxonómicos) en remanentes de bosques de Yungas en el noroeste argentino es hoy una realidad irrefutable. Aunque la alta diversidad biológica de esta ecorregión es un hecho conocido y valorado, son escasas las investigaciones detalladas para la mayoría de los grupos taxonómicos allí presentes, por lo que nuestro conocimiento de su riqueza de especies es aún limitado. Estudios realizados en los últimos años han resultado en numerosos aportes al conocimiento de las especies de diversos grupos taxonómicos (e. g., Barquez et al., 1999a; Molineri, 2001; Lavilla et al., 2003; Mirande et al., 2004a,b; Molineri y Zúñiga, 2004; Aguilera y Mirande, 2005, 2006; Jayat y Miotti, 2005; Barquez et al., 2006b; Jayat et al., 2006; 2006), poniendo de manifiesto nuestra falta de información acerca de la misma. En este contexto, contribuciones recientes (Barquez et al., 1999a; Barquez y Díaz, 2001; Jayat y Miotti, 2005; Jayat et al., 2009) y este trabajo han permitido documentar la presencia actual de por lo menos 74 de las 97 especies consideradas como probablemente presentes para el pedemonte de Yungas del extremo norte de Argentina, destacándose el registro de especies posiblemente nuevas para la ciencia (e. g., representantes de los géneros Micronycteris, Myotis y Oligoryzomys), especies de difícil observación y/o con escasos registros para la región (e. g., Monodelphis kunsi, Noctilio leporinus, Glossophaga soricina, Pygoderma bilabiatum, Sturnira oporaphylum, Leopardus tigrinus, L. wiedii, Akodon fumeus, A. sylvanus, Coendou bicolor), especies probablemente endémicas de la ACRB en Argentina (e. g., Akodon sylvanus) y de especies de mediano y gran porte restringidas en Argentina al sector norte de Yungas o que presentan problemas de conservación (e. g., Dasypus yepesi, Myrmecophaga tridactyla, Panthera onca, Nasua nasua, Tapirus terrestris, Tayassu pecari).
La mayor parte de las especies potencialmente presentes en el área de estudio y que no se registraron en nuestros relevamientos correspondieron a quirópteros, el grupo más diverso dentro de la comunidad de mamíferos de áreas pedemontanas. Para este grupo sólo pudimos registrar el 65% de las especies consideradas presentes en el área. Este bajo éxito de captura puede explicarse por diversos factores, incluyendo la poca eficacia de las redes para capturar algunas de las especies insectívoras de vuelo alto (e. g., representantes de Eumops) y la baja actividad que muestran los individuos de este grupo en condiciones de baja temperatura ambiente y durante ciertos momentos del ciclo lunar. Indudablemente, el muestreo apropiado de los quirópteros demanda la utilización de redes de niebla colocadas en otros estratos de la vegetación y de métodos complementarios como la búsqueda activa de ejemplares en madrigueras o el de grabaciones ultrasónicas de llamadas de ecolocalización como las realizadas en otros estudios. Por ejemplo, en los inventarios llevados a cabo por Simmons y Voss (1998) sobre la fauna de quirópteros de Paracú, en la Guayana Francesa, los relevamientos de quirópteros mediante redes de niebla colocadas al nivel del suelo sólo permitieron registrar el 83% del total de especies presentes en el área.
En general, en la actualidad se acepta que los acelerados procesos de modificación del paisaje natural producidos desde la llegada de los conquistadores españoles han generado, en gran medida, retracciones en la distribución de numerosas especies de mamíferos y una disminución de sus tamaños poblacionales (Roig, 1991; Chebez, 1999; Barquez y Díaz, 2001; Ojeda et al., 2008). Sin embargo, salvo en casos emblemáticos como los del yaguareté y el aguará guazú (e. g., Arra, 1974; Roig, 1991; Perovic y Herrán, 1998; Chebez, 1999 y referencia allí; Perovic, 2002; Prevosti et al., 2004 y referencias allí; Miatello y Cobos, 2008), nuestro conocimiento detallado, cuantitativo y sistemático de estos procesos y sus consecuencias es prácticamente nulo (Brown et al., 2001). Para grupos que no gozan del estatus de "estandartes de la conservación", como los micromamíferos, el conocimiento acerca de la influencia antrópica, e incluso de los procesos naturales, sobre parámetros poblacionales y comunitarios no sólo es escaso sino también difícil de alcanzar. Es así que este trabajo es de particular relevancia para el conocimiento cuantitativo de la diversidad del grupo en la región, aportando estimaciones de riqueza, dominancia y abundancia relativa para un conjunto de 42 especies de pequeños mamíferos. A pesar de esto, la totalidad de los sitios relevados aquí se encuentran bajo algún grado de perturbación antrópica (emprendimientos forestales y agropecuarios, construcción de gasoductos, oleoductos o electroductos). En consecuencia, los valores obtenidos sólo representan una situación pauperizada, típica de ambientes de Yungas caracterizados por crecimiento secundario, y no la de ambientes en buen estado de conservación. La medición de parámetros de biodiversidad de ambientes de Yungas bien conservados, aún presentes en ciertos sectores de la ACRB, constituye una tarea pendiente fundamental para establecer una línea de base adecuada y un parámetro de comparación. Datos preliminares aún no publicados indican diferencias sutiles en cuanto a riqueza de especies de micromamíferos no voladores en sectores pedemontanos relativamente bien conservados del Parque Nacional Calilegua respecto de sectores muy modificados en sus alrededores. Sin embargo, se encontraron diferencias importantes en cuanto a la equitatividad del ensamble y a la abundancia relativa para todas las especies analizadas, con una distribución más equitativa de los individuos entre especies y valores de abundancia relativa notablemente altos para formas típicas de ambientes boscosos (e. g., Thylamys sponsorius, Akodon simulator, Euryoryzomys legatus, Oxymycterus paramensis) y extremadamente bajos para aquellas que son dominantes en ambientes modificados (e. g., Holochilus chacarius; Calomys cf. C. fecundus).
Aunque el esfuerzo de captura de este estudio fue, en líneas generales, considerable (con casi 30 000 m r/h y 10 000 t/n en 102 noches de muestreo), los resultados obtenidos para algunos análisis deben ser tomados con cautela y complementados con estudios adicionales. En este sentido, los resultados obtenidos para quirópteros fueron, en general, menos satisfactorios (Tabla 2). Para este grupo las diferencias entre la riqueza observada y esperada y los desvíos respecto del valor promedio de S* fueron siempre mayores. Además, las curvas acumulativas de la riqueza de especies siempre estuvieron más alejadas de la asíntota (Fig. 2).
Los valores de abundancia relativa indican densidades extremadamente bajas para algunas de las especies de quirópteros registradas. Este es el caso de Chrotopterus auritus, Glossophaga soricina, Molossops teminckii y Pygoderma bilabiatum, con menos de 0.08 individuos por cada 100 mr/h. Tres de estas especies fueron, además, registradas en sólo una de las 22 localidades relevadas, por lo que el estatus de sus poblacionales debería ser estudiado más en detalle. Otras especies no documentadas en este estudio, probablemente por encontrarse también en densidades muy bajas, incluyen a Anoura caudifer, Diaemus youngi, Micronycteris sp., Tonatia bidens y Nyctinomops laticaudatus.
Los resultados obtenidos en este estudio indican escasas diferencias en composición, diversidad y abundancia de micromamíferos entre las estaciones seca y húmeda. Las diferencias más notorias, relacionadas con cambios en la composición de especies, pueden explicarse principalmente por la poca eficacia de los muestreos. Los quirópteros, el grupo para el que los resultados de los inventarios fueron menos satisfactorios, incluyen la mayoría de las diferencias registradas. Además, muchas de las especies documentadas exclusivamente en una sola temporada son formas consideradas raras o poco abundantes en el área de estudio, algunas de ellas registradas para sólo una o dos localidades de las 22 analizadas (e. g., Noctilio leporinus, Pygoderma bilabiatum, Sturnira oporaphilum).
Debido a la notable complejidad de la problemática ambiental en el pedemonte de Yungas en Argentina, la preservación de su biodiversidad sólo será posible en el marco de una estrategia de generación de conocimiento cuantitativo que involucre, además, una estrecha colaboración entre los distintos actores sociales de la región. Por muchas de sus características, la ACRB parece ser el sector ideal para la obtención de este objetivo. Es en este
contexto que la elaboración de una línea de base ambiental y la organización de sistemas de monitoreos de biodiversidad a mediano y largo plazo constituyen una tarea fundamental.
Sector Noreste
Sitio 1: Macueta Norte, km 72 de la ruta que se dirige al Pozo 1001 b, 1148 m (Gral. José de San Martín, Salta). 22º 01' 13.8" S, 63° 54' 23.2" W.
Sitio 2: Campo Largo, km 74.5 de la Ruta entre Acambuco y Campo Largo, 684 m (Gral. José de San Martín, Salta). 22º 01' 43.2" S, 63º 55' 35.9" W.
Sitio 3: Bajo Macueta, aprox. 10 km al NE de Acambuco, sobre Quebrada San Francisco, 736 m (Gral. José de San Martín, Salta). 22° 05' 57.5" S, 63° 53' 39.3" W.
Sitio 4: Finca Falcón, aprox. 3 km al NW del Pozo de agua N° 5, cercanías del Río Seco, 704 m (Gral. José de San Martín, Salta). 22° 18' 41.4" S; 63° 58' 7.1" W.
Sitio 5: Escuela Histórica San Pedrito, 1100 m (Gral. José de San Martín, Salta). 22° 22' 57.6" S; 64° 00' 4.9" W.
Sitio 6: aprox. 20 km (por ruta) al WNW de Vespucio, sobre el río Seco (Gral. José de San Martín, Salta). 22º 32' 16.3" S; 64º 00' 2.05" W.
Sitio 7: camino a la antena repetidora Yacuy, altura del km 12.5 de la ruta provincial N° 54, aprox. 13 km (por ruta) al W de Piquirenda (Gral. José de San Martín, Salta). 22º 19' 48.5" S; 63º 49' 48.6" W.
Sitio 8: aprox. 1 km al ENE del puesto Las Vertientes, 1053 m (Gral. José de San Martín, Salta) 22º 00' 21.7" S, 63º 44' 36.8" W.
Sitio 9: Finca Río Seco, 40 km al N de Embarcación, 524 m (Gral. José de San Martín, Salta) 22º 51' 6.33" S, 64º 04' 45.5" W.
Sitio 10: aprox. 36 km al NNW de Embarcación, Finca Río Seco, 611 m (Gral. José de San Martín, Salta) 22º 53' 35.4' S, 64º 08' 07" W.
Sector Central
Sitio 11: Finca El Carmen, cultivo de Eucaliptus en la entrada de la propiedad, 362 m (Orán, Salta). 23° 07' 05" S, 64° 16' 39.5" W.
Sitio 12: Finca El Carmen, bosque nativo al fondo de la propiedad, 396 m (Orán, Salta). 23° 06' 51.1" S, 64° 15' 03.3" W.
Sitio 13: Finca El Carmen, bosque inundable al fondo de la propiedad, 315 m (Orán, Salta). 23° 05' 10.5" S, 64° 16' 08" W.
Sitio 14: Finca Alto Verde, 670 m (Orán, Salta) 23º 13' 38.4" S; 64º 32' 35.1" W.
Sector Sur
Sitio 15: Finca FAMAT, 409 m (Ledesma, Jujuy) 23º 37' 26.8" S, 64º 30' 58.2" W.
Sitio 16: Finca Sauzalito, aprox. 1 km al E del cruce entre la ruta nacional N° 34 y el río Yuto, 413 m (Ledesma, Jujuy) 23º 39' 42.6" S, 64º 32' 2.8" W.
Sitio 17: Finca El Piquete, aprox. a 3 km del cruce del Río Tamango con la senda Maderera, 820 m (Santa Bárbara, Jujuy). 24º 10' 30.6" S, 64º 34' 20.1" W.
Sitio 18: Finca El Piquete, márgenes del Río Tamango, aprox. a 1.5 km del cruce del río con la senda maderera, 820 m (Santa Bárbara, Jujuy). 24º 10' 10.5" S, 64º 34' 13.5" W.
Sitio 19: Finca El Piquete, márgenes del Río Volcán, aprox. a 5 km del cruce del Río Tamango y la senda maderera, 973 m (Santa Bárbara, Jujuy). 24º 11' 8.3" S, 64º 33' 34.2" W.
Sitio 20: Finca Las Moras, 460 m (Santa Bárbara, Jujuy). 23º 49' 38.8" S, 64º 31' 33.5" W.
Sitio 21: Planta Caimancito, Batería, 535 m (Ledesma, Jujuy). 23º 38' 40.7" S, 64º 36' 11.9" W.
Sitio 22: aprox. 2 km al NW de la intersección entre la ruta nacional 34 y el camino a la batería de la planta Caimancito, 461 m (Ledesma, Jujuy) 23º 39' 45.9" S, 64º 34' 52.9" W.
AGRADECIMIENTOS
Agradecemos la gentil colaboración de C. Alonso Herrera, D. Delgado, C. Aguirre, D. Miotti, G. Námen, U. Pardiñas y W. Villafañe (Almita) en las tareas de campo. Agradecemos también a los curadores de las colecciones estudiadas que gentilmente permitieron el acceso a los materiales depositados en las mismas. El mapa de distribución fue realizado por S. Pacheco del sistema de información geográfico ambiental (SIGA) de la Fundación Proyungas. Cecilia Blundo y P. Blendinger colaboraron en la descripción de los distintos sectores de Yungas considerados en este estudio. Los comentarios y sugerencias de M. Di Bitetti y C. Galliari mejoraron substancialmente la calidad del manuscrito original. Este trabajo fue principalmente solventado mediante fondos aportados por la Fundación ProYungas y se llevó a cabo con el apoyo institucional del Laboratorio de Investigaciones Ecológicas de las Yungas del Instituto de Ecología Regional (IER-LIEY) y la Cátedra de Paleontología de la Facultad de Ciencias Naturales e Inst. M. Lillo, UNT.
LITERATURA CITADA
1. AGUILERA G y JM MIRANDE. 2005. A new species of Jenynsia (Cyprinodontiformes: Anablepidae) from northwestern Argentina and its phylogenetic relationships. Zootaxa 1096:29-39. [ Links ]
2. AGUILERA G y JM MIRANDE. 2006. First record of Cetopsis starnesi (Siluriformes: Cetopsidae) from Argentina. Zeitschrift für Fischkunde 8:101-102. [ Links ]
3. ALLEN JA.1901. New South American Muridae and a new Metachirus. Bulletin of the American Museum of Natural History 14:405-412 [ Links ]
4. ARRA MA. 1974. Distribución de Leo onca (L) en Argentina. Neotropica 20:156-158. [ Links ]
5. AUTINO AG, GL CLAPS y RM BARQUEZ. 1999. Insectos ectoparásitos de murciélagos de las yungas de la Argentina. Acta Zoológica Mexicana, nueva serie 78:119-169. [ Links ]
6. BARQUEZ RM, JK BRAUN, MA MARES, JP JAYAT y D FLORES. 1999a. First record for Argentina for a bat in the genus Micronycteris. Mammalia 63:368-372. [ Links ]
7. BARQUEZ RM, MA MARES y JK BRAUN. 1999 b. The bats of Argentina. Special Publications, Museum of Texas Tech University 42. [ Links ]
8. BARQUEZ RM y MM DÍAZ. 2001. Bats of the Argentine Yungas: a systematic and distributional analysis. Acta Zoológica Mexicana, nueva serie 82:29-81. [ Links ]
9. BARQUEZ RM, MM DÍAZ y RA OJEDA (eds.). 2006a. Mamíferos de Argentina, sistemática y distribución. Sociedad Argentina para el Estudio de los Mamíferos, Tucumán. [ Links ]
10. BARQUEZ RM, LI FERRO y MS SÁNCHEZ. 2006 b. Tapecomys primus (Rodentia: Cricetidae), nuevo género y especie para la Argentina. Mastozoología Neotropical 13:117-121. [ Links ]
11. BIANCHI NO, OA REIG, OJ MOLINA y FN DULOUT. 1971. Cytogenetics of the South American akodont rodents (Cricetidae). I. A progress report of Argentinian and Venezuelan forms. Evolution 25:724-736. [ Links ]
12. BROWN AD. 1995 a. Introducción y conclusiones de trabajo en talleres de la Primera Reunión Regional sobre Selvas Subtropicales de Montaña. Pp. 1-8, en: Investigación, conservación y desarrollo en las selvas subtropicales de montaña (AD Brown y HR Grau, eds.). Laboratorio de Investigaciones Ecológicas de las Yungas, UNT. [ Links ]
13. BROWN AD. 1995 b. Las selvas de montaña del noroeste de Argentina: problemas ambientales e importancia de su conservación. Pp. 9-18, en: Investigación, conservación y desarrollo en las selvas subtropicales de montaña (AD Brown y HR Grau, eds.). Laboratorio de Investigaciones Ecológicas de las Yungas, UNT. [ Links ]
14. BROWN AD, SC CHALUKIAN, L MALMIERCA y OJ COLILLAS. 1986. Habitat structure and feeding behavior of Cebus apella in El Rey National Park, Argentine. Pp. 137-151, en: Current perspectives in primate social dynamics (DM Taub y Eking, eds.). Van Nostrand Reinhold Company. [ Links ]
15. BROWN AD, HR GRAU, LR MALIZIA y A GRAU. 2001. Argentina. Pp. 623-659, en: Bosques Nublados del Neotrópico (M Kappelle y AD Brown, eds.). Instituto Nacional de Biodiversidad (INBio). Costa Rica. [ Links ]
16. CHAO A. 1987. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 43:783-791. [ Links ]
17. CHEBEZ JC. 1999. Los que se van. Especies argentinas en peligro. Editorial Albatros, SACI, Buenos Aires. [ Links ]
18. COLWELL RK y JA CODDINGTON. 1994. Estimating terrestrial biodiversity through extrapoation. Philosophical Transactions (B) of the Royal Society of London 345:101-118. [ Links ]
19. COLWELL RK, CX MAO y J CHANG. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85:2717-2727. [ Links ]
20. CONTRERAS JR. 1999. El género Ctenomys en la Provincia de Tucumán, República Argentina, con la descripción de una nueva especie (Rodentia, Ctenomyidae). Ciencia Siglo XXI, Fundación Bartolomé Hidalgo, Buenos Aires, Argentina 3:1-31. [ Links ]
21. DIAZ GB y RA OJEDA. 2000. Libro Rojo de Mamíferos Amenazados de la Argentina. Sociedad Argentina para el Estudio de los Mamíferos. [ Links ]
22. DÍAZ MM, JK BRAUN, MA MARES y RM BARQUEZ. 2000. An update of the taxonomy, systematics, and distribution of mammals of Salta province, Argentina. Occasional Papers, Sam Noble Oklahoma Museum of Natural History 10:1-52. [ Links ]
23. DÍAZ MM y RM BARQUEZ. 2007. The wild mammals of Jujuy province, Argentina: Systematics and distribution. Pp. 417-578, en: The quintessential naturalist: Honoring the life and legacy of Oliver P. Pearson (DA Kelt, EP Lessa, J Salazar-Bravo y JL Patton, eds.). University of California Publications in Zoology, California. [ Links ]
24. DÍAZ MM, RM BARQUEZ, JK BRAUN y MA MARES. 1999. A new species of Akodon (Muridae: Sigmodontinae) from northwestern Argentina. Journal of Mammalogy 80:786-798. [ Links ]
25. DÍAZ MM, DA FLORES y RM BARQUEZ. 1998. Instrucciones para la preparación y conservación de mamíferos. Publicaciones especiales 1. PIDBA. Universidad Nacional de Tucumán, Tucumán. [ Links ]
26. DÍAZ MM, DA FLORES y RM BARQUEZ. 2002. A new species of gracile mouse opossum, genus Gracilinanus, from Argentina. Journal of Mammalogy 83:824-833. [ Links ]
27. FLORES DA, MM DÍAZ y RM BARQUEZ. 2007. Systematics and distribution of marsupials in Argentina: A review. Pp. 579-669 en: The quintessential naturalist: Honoring the life and legacy of Oliver P. Pearson (DA Kelt, EP Lessa, J Salazar-Bravo y JL Patton, eds.). University of California Publications in Zoology, California. [ Links ]
28. GIANNINI NP. 1999. Selection of diet and elevation by sympatric species of Sturnira in an Andean rainforest. Journal of Mammalogy 80:1186-1195. [ Links ]
29. GIL G y S HEINONEN FORTABAT. 2003. Lista comentada de los mamíferos del Parque Nacional Baritú (Salta, Argentina). Acta Zoológica Lilloana 47:117-135. [ Links ]
30. HEINONEN FORTABAT S y A BOSSO. 1994. Nuevos aportes para el conocimiento de la mastofauna del Parque Nacional Calilegua (Provincia de Jujuy, Argentina). Mastozoología Neotropical 1:51-60. [ Links ]
31. HERSHKOVITZ P. 1962. Evolution of Neotropical cricetine rodents (Muridae) with special reference to the phyllotine group. Fieldiana, Zoology 46:1-524. [ Links ]
32. JAYAT JP, RM BARQUEZ, MM DÍAZ y PJ MARTINEZ. 1999. Aportes al conocimiento de la distribución de los carnívoros del noroeste de Argentina. Mastozoología Neotropical 6:15-30. [ Links ]
33. JAYAT JP, G D'ELIA, UFJ PARDIÑAS y JG NAMEN. 2007. A new species of Phyllotis (Rodentia, Cricetidae, Sigmodontine) from the upper montane forest of the Yungas of northwestern Argentina. Pp. 775-798, en: The Quintessential Naturalist: Honoring the Life and Legacy of Oliver P. Pearson (D Kelt, E Lessa y J Salazar-Bravo, eds.). California: University of California Publications in Zoology. [ Links ]
34. JAYAT JP, G D'ELÍA, UFJ PARDIÑAS, MD MIOTTI y PE ORTIZ. 2008 a. A new species of the genus Oxymycterus (Mammalia: Rodentia: Cricetidae) from the vanishing Yungas of Argentina. Zootaxa 1911:31-51. [ Links ]
35. JAYAT JP y MD MIOTTI. 2005. Primer registro de Monodelphis kunsi (Didelphimorphia, Didelphidae) para Argentina. Mastozoología Neotropical 12:253-256. [ Links ]
36. JAYAT JP, PE ORTIZ y MD MIOTTI. 2008 b. Distribución de sigmodontinos (Rodentia: Cricetidae) en pastizales de neblina del noroeste argentino. Acta Zoológica Mexicana 24:137-177. [ Links ]
37. JAYAT JP, PE ORTIZ y MD MIOTTI. 2009. Mamíferos de la selva pedemontana del noroeste argentino. Pp. 1-44, en: Ecología, historia natural y conservación de la selva pedemontana de las yungas australes (AD Brown, PG Blendinger, T Lomáscolo y P García Bes, eds.). Ediciones del Subtrópico. [ Links ]
38. JAYAT JP, PE ORTIZ, P TETA, UFJ PARDIÑAS y G D'ELÍA. 2006. Nuevas localidades argentinas para algunos roedores sigmodontinos (Rodentia: Cricetidae). Mastozoología Neotropical 13:51-67. [ Links ]
39. LAVILLA EO, M VAIRA y L FERRARI. 2003. "A new species of Elachistocleis (Anura: Microhylidae) from the Andean Yungas of Argentina, with comments on the Elachistocleis ovalis - E. bicolor controversy". Amphibia-Reptilia 24:269-284. [ Links ]
40. MAGURRAN. 1998. Ecological diversity and its measurement. Princeton University Press. [ Links ]
41. MARES MA, RA OJEDA y RM BARQUEZ. 1989. Guía de los mamíferos de la provincia de Salta, Argentina. University of Oklahoma Press, Norman, Oklahoma. [ Links ]
42. MARES MA, RA OJEDA, JK BRAUN y RM BARQUEZ. 1997. Systematics, distribution, and ecology of the mammals of Catamarca Province, Argentina. Pp. 89-141, en: Life among the muses: papers in honor of James S. Findley (TL Yates, WL Gannon y DE Wilson, eds.). The Museum of Southwestern Biology, The University of New Mexico, Albuquerque. [ Links ]
43. MIATELLO R y V COBOS. 2008. Nuevos aportes sobre la distribución del aguará guazú (Chrysocyon brachyurus, Carnivora: Canidae) en las provincias de Córdoba y Santiago del Estero, Argentina. Mastozoología Neotropical 15:209-213. [ Links ]
44. MINETTI JL, WM VARGAS y SA ALBARRACÍN. 2005. La variabilidad espacio-temporal de las precipitaciones en el noroeste argentino. Pp. 187-215, en: El clima del noroeste argentino (JL Minetti, ed.). Editorial Magna. Tucumán. [ Links ]
45. MIRANDE JM, G AGUILERA y M DE LAS M AZPELICUETA. 2004 a. A new genus and species of small characid (Ostariophysi, Characidae) from the upper Río Bermejo basin, northwestern Argentina. Revue Suisse de Zoologie 111:715-728. [ Links ]
46. MIRANDE JM, G AGUILERA y M DE LAS M AZPELICUETA. 2004 b. A new species of Astyanax (Characiformes, Characidae) from the upper Río Bermejo basin. Revue suisse de Zoologie 111:213-223. [ Links ]
47. MIRANDE JM, G AGUILERA y M DE LAS M AZPELICUETA. 2006. Astyanax endy (Characiformes: Characidae), a new fish species from the upper Río Bermejo basin, northwestern Argentina. Zootaxa 1286:57-68. [ Links ]
48. MOLINERI C. 2001. Description of a new genus of Leptohyphidae (Ephemeroptera). Pp. 337-345, en: Trends in Research in Ephemeroptera and Plecoptera (E Dominguez, ed.). Kluwer Academic/Plenum Publishers, New York. [ Links ]
49. MOLINERI C y MC ZÚÑIGA. 2004. Lumahyphes, a new genus of Leptohyphidae (Ephemeroptera). Aquatic Insects 26:19-30. [ Links ]
50. MYERS P. 1989. A preliminary revision of the varius group of Akodon (A. dayi, dolores, molinae, neocenus, simulator, toba, and varius). Pp. 5-54, en: Advances in Neotropical Mammalogy (KH Redford y JF Eisenberg, eds.). Sandhill Crane Press, Inc., Gainesville, FL. ix+614 pp. [ Links ]
51. MYERS P, JL PATTON y MF SMITH. 1990. A review of the boliviensis group of Akodon (Muridae: Sigmodontinae) with emphasis on Peru and Bolivia. Miscellaneous Publications of the Museum of Zoology, University of Michigan 177:1-89. [ Links ]
52. OJEDA RA, RM BARQUEZ, J STADLER y R BRANDL. 2008. Decline of mammals species diversity along the Yungas forest of Argentina. Biotropica 40:515-521. [ Links ]
53. OJEDA RA, J STADLER y R BRANDL. 2003. Diversity of mammals in the tropical-temperate Neotropics: hotspots on a regional scale. Biodiversity and Conservation 12:1431-1444. [ Links ]
54. OJEDA RA y MA MARES. 1989. A biogeographic analysis of the mammals of Salta Province, Argentina. Special Publication 27, The Museum, Lubbock, Texas Tech University Press. [ Links ]
55. OLDS N. 1988. A revision of the Genus Calomys (Rodentia: Muridae). Ph. D. dissertation, City University of New York, New York, USA, 279 pp. [ Links ]
56. OLROG CC. 1979. Los mamíferos de la selva húmeda, Cerro Calilegua, Jujuy. Acta Zoológica Lilloana 33:9-14. [ Links ]
57. ORTIZ PE y UFJ PARDIÑAS. 2001. Sigmodontinos (Mammalia: Rodentia) del Pleistoceno tardío del valle de Tafí (Tucumán, Argentina): taxonomía, tafonomía y reconstrucción paleoambiental. Ameghiniana 38:3-26. [ Links ]
58. PEARSON OP. 1958. A taxonomic revision of the rodent genus Phyllotis. University of California Publications, Zoology 56:391-496. [ Links ]
59. PEROVIC PG. 2002. La comunidad de félidos de las selvas nubladas del noroeste argentino. Tesis doctoral inédita, Facultad de Ciencias Exactas, Físicas y Naturales. Universidad Nacional de Córdoba, Córdoba, Argentina. [ Links ]
60. PEROVIC PG y M HERRÁN. 1998. Distribución del jaguar Panthera onca en las provincias de Jujuy y Salta, noroeste de Argentina. Mastozoología Neotropical 5:47-52. [ Links ]
61. PREVOSTI FJ, BONOMO M y EP TONNI. 2004. La distribución de Chrysocyon brachyurus (Illiger, 1811) (Mammalia: Carnivora: Canidae) durante el Holoceno en la Argentina: implicancias paleoambientales. Mastozoología Neotropical 11:27-43. [ Links ]
62. ROIG VG. 1991. Desertification and distribution of mammals in the Southern Cone of South America. Pp. 239-279, en: Latin American mammals: their conservation, ecology, and evolution (MA Mares y DJ Schmidly, eds.). University of Oklahoma Press. [ Links ]
63. SÁNCHEZ F, P SÁNCHEZ-PALOMINO y A CADENA. 2008. Species richness and indices of abundance of medium-sized mammals in Andean forest and reforestations with Andean Alder: a preliminary analysis. Caldasia 30:197-208. [ Links ]
64. SIMMONS NB y DP VOSS. 1998. The mammals of Paracou, French Guiana: a Neotropical lowland rainforest fauna Part 1. Bats. Bulletin of the American Museum of Natural History 263:1-219. [ Links ]
65. THOMAS O. 1918. On small Mammals form Salta and Jujuy collected by Mr. E. Budin. Annals and Magazine of Natural History (9)1:186-193. [ Links ]
66. THOMAS O. 1921. New Rhipidomys, Akodon, Ctenomys and Marmosa from the Sierra Santa Barbara, S.E. Jujuy. Annals and Magazine of Natural History (9)7:181-187. [ Links ]
67. VARELA RO y AD BROWN. 1995. Tapires y pecaríes como dispersores de plantas en los bosques húmedos subtropicales de Argentina. Pp. 129-140, en: Investigación, Conservación y Desarrollo en Selvas Subtropicales de Montaña. (AD Brown y HR Grau, eds.). Proyecto de Desarrollo Agroforestal / L.I.E.Y. [ Links ]
68. VOSS RS, DP LUNDE y NB SIMMONS. 2001. The mammals of Paracou, French Guiana: a Neotropical lowland rainforest fauna Part 2. Nonvolant species. Bulletin of the American Museum of Natural History 263:1-236. [ Links ]
69. YEPES J. 1935. Consideraciones sobre el genero Andinomys (Cricetinae) y descripción de una forma nueva. Anales del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 38:333-348. [ Links ]
Recibido 24 marzo 2009.
Aceptado 25 julio 2009.
Editor asociado: UFJ Pardiñas

