










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkMastozoología neotropical
Print version ISSN 0327-9383
Mastozool. neotrop. vol.18 no.2 Mendoza July/Dec. 2011
ARTÍCULOS Y NOTAS
Morfometría de las paleocuevas de la "Fm." Chapadmalal y su asignación a Actenomys (Rodentia), Paedotherium (Notoungulata) y otros mamíferos fósiles hospedantes
Andrea Elissamburu1, Alejandro Dondas2 y Luciano De Santis1
1 Cátedra Anatomía Comparada, Facultad de Ciencias Naturales y Museo de La Plata, Universidad Nacional de La Plata, Calle 64 Nº 3 e/ 119 y 120, 1900 La Plata, Argentina [Correspondencia: Andrea Elissamburu <elissamburu@hotmail.com>].
2 Museo de Ciencias Naturales de Mar del Plata, Plaza España s/n, 7600 Mar del Plata, Argentina.
RESUMEN: En la "Formación" Chapadmalal, aflorante en las barrancas que se extienden desde Mar del Plata a Miramar, se diferencian paleocuevas que han sido asignadas al roedor fósil Actenomys sp. (Rodentia, Caviomorpha). La asignación se basa en la elevada frecuencia de restos de Actenomys sp. hallados en estas bioestructuras y en su capacidad cavadora. Si bien es evidente que Actenomys sp. construía cuevas, y muchas de las paleocuevas le corresponden, no es la única especie coetánea que pudo haberlas construido. Se analizaron morfométricamente las paleocuevas aflorantes en las barrancas costeras de Mar del Plata-Miramar correspondientes a los Pisos/Edad Chapadmalalense y Marplatense y la identificación de los posibles constructores. Se seleccionaron las paleocuevas en sección transversal y se tomaron dos medidas estructurales, correspondientes al diámetro transverso y dorsoventral, con las que se construyó un índice estructural. Se registró el material fósil asociado a las mismas en los casos donde estaba expuesto. Los análisis estadísticos incluyeron distribución de frecuencias, correlaciones, análisis de residuales y diferencia de medias, en las medidas individuales y en el índice. Se identifican cuevas construidas por Actenomys sp., en estadios juveniles y adultos, y por Paedotherium sp. Se reconocen diferencias métricas entre las cuevas del Chapadmalalense y Marplatense, posiblemente asociadas a una diferencia en la abundancia relativa de Paedotherium sp. y Actenomys sp. Asimismo se refuerza la existencia de cuevas de Lagostomopsis sp. y se identifica la posible construcción, o uso oportunista, de las mismas por Eumysops sp., Dolicavia sp. y otros cávidos.
ABSTRACT: Morphometry of paleocaves at the Chapadmalal "Formation" assigned to fossorial rodent Actenomys sp. (Rodentia, Caviomorpha), Paedotherium (Notoungulata), and to other fossil mammals. Paleocaves assigned to fossorial rodent Actenomys sp. (Rodentia, Caviomorpha) are present at the Chapadmalal "Formation", cropping out from Mar del Plata to Miramar. The assignation is based on the high frequency of fossil material of Actenomys sp. within the paleocaves and in the digging capacity recognized for the genera. Although it is evident that Actenomys sp. built caves and most of these correspond to the genera, it is not the only coetaneal mammal with the capacity of construction of these structures. Paleocaves of Chapadmalalan and Marplatan age were analyzed morphometrically and the generic assignation of their producers was evaluated. Paleocaves in transverse section were selected, its dorsoventral and lateromedial diameter was measured and used for calculating a structural index. The fossil material exposed in paleocaves was determined to a specific level in all possible cases. The statistical analyses performed include frequency distributions, correlations, residual analyses, and mean differences on individual measurements and on the index. Paleocaves constructed by Actenomys sp., in juvenile and adult stages, and by Paedotherium sp. are recognized. Metric difference between paleocaves of Chapadmalalan and Marplatan ages are recognized, in possible association with a different abundance of Paedotherium sp. and Actenomys sp. The presence of paleocaves of Lagostomopsis sp. is reinforced and is suggested the possible construction, or opportunistic use, of paleocaves by Eumysops sp., Dolicavia sp. and other Caviidae.
Palabras clave: Actenomys; "Fm." Chapadmalal; Paedotherium; Paleocuevas; Paleoecología.
Key words: Actenomys; Chapadmalal "Fm."; Paedotherium; Paleocaves; Paleoecology.
INTRODUCCIÓN
En la "Formación" Chapadmalal, aflorante en las barrancas que se extienden desde Mar del Plata a Miramar (Buenos Aires, Argentina), se diferencian paleocuevas (Ameghino, 1908; Frenguelli, 1921; Kraglievich, 1952) que han sido asignadas al roedor fósil Actenomys sp. (Rodentia, Caviomorpha, Octodontidae) (Genise, 1989). La asignación de las paleocuevas a este roedor se basa en la elevada frecuencia de restos articulados de Actenomys sp. hallados en el interior de estas bioestructuras y en la capacidad cavadora reconocida para el género (Genise, 1989; Fernández et al., 2000). Actenomys sp. se registra durante el Plioceno, en los Pisos/Edad Montehermosense, Chapadmalalense y Marplatense de la Provincia de Buenos Aires (Cione y Tonni, 1995a, b), reconociéndose la especie Actenomys priscus (Verzi, 2002).
A. priscus es actualmente identificado como dentoexcavador, aunque no se descarta que utilizara sus miembros anteriores en forma complementaria (Fernández et al., 2000) para cavar sus cuevas en suelos arcillosos y cohesivos (Scognamillo, 1993). Si bien sus hábitos no habrían sido mayormente hipogeos y tendría una estrecha relación con la superficie, se trataría de un excavador activo, que utilizaba las cuevas para funciones posiblemente distintas a la de obtención de alimento, como ser protección y cuidado de crías (Fernández et al., 2000).
Genise (1989) describió las paleocuevas de A. priscus como formadas por túneles de 13 cm de diámetro que penetran en el suelo con una inclinación de 30º a 80º y de hasta 5 m de largo. Éstas presentan porciones horizontales que frecuentemente preceden a otras oblicuas, con túneles rectos, que también pueden ser curvos, sigmoideos y ramificados y terminar en una cámara ovoide, en algunos casos con más de una cámara conectada al túnel principal por un corto túnel lateral (Genise, 1989) (Fig. 1).
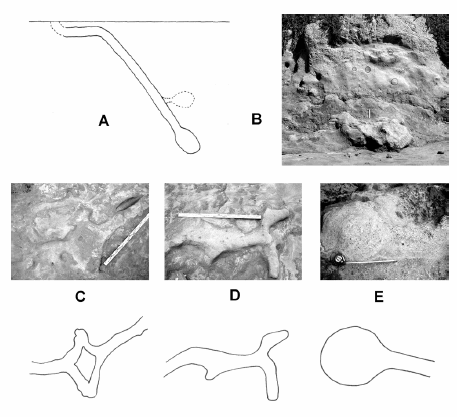
Fig. 1. Cuevas asignadas a Actenomys sp. A. Esquema adaptado del trabajo de Genise (1989); B. Imagen de las paleocuevas aflorantes en las barrancas de la "Fm." Chapadmalal; las mismas en vista transversal; C, D. Paleocuevas vistas en planta; E. Madriguera vista en planta.
Si bien resulta evidente que A. priscus construía cuevas y muchas de las estructuras observadas le corresponden, no es la única especie coetánea que presentaba capacidad cavadora y que además, dado el tamaño corporal, también puede haber construido cuevas similares. Genise (1989) reconoce la presencia de Lagostomopsis sp. como constructor de cuevas de similar diámetro o un poco mayor que las de A. priscus, y reconstruye el ambiente Chapadmalalense de la zona como ocupado en su mayoría por cuevas de A. priscus con algunas vizcacheras intercaladas.
Paedotherium sp. (Notoungulata, Hegetotheriidae) se ha encontrado asociado a las paleocuevas (Ameghino, 1908; Frenguelli, 1921; Genise, 1989; Scognamillo, 1993; Peña, 1997), constituye después de A. priscus el mamífero más estrechamente ligado a éstas (Genise, 1989) y se le ha atribuido la construcción de cuevas dada su capacidad cavadora (Elissamburu, 2001; Elissamburu, 2004). Peña (1997) reconoce restos de Paedotherium sp. en paleocuevas de tamaño similar a las de A. priscus y en paleocuevas un poco más grandes, por lo que es esperable que existan algunas diferencias morfométricas entre las cuevas de estas especies coetáneas, principalmente dado el mayor tamaño corporal de Paedotherium sp. Otros mamíferos que se han encontrado asociados a paleocuevas son Dolicavia sp. (Genise, 1989), Eumysops sp. y Microcavia sp. (Peña, 1997). El tamaño de estas especies es similar o menor al de A. priscus, por lo que pueden haber tenido un uso oportunista de las cuevas, o bien corresponderse con paleocuevas similares o de menor diámetro.
Hasta el momento, los trabajos vinculados a las paleocuevas de la "Formación" Chapadmalal comprenden el estudio de la estructura general de éstas y su atribución a Actenomys sp. (Genise, 1989), el estudio de estas estructuras asignadas a A. priscus junto con su significado paleoecológico y estratigráfico (Scognamillo, 1993) y el análisis desde un punto de vista tafonómico de las mismas que denota una preservación diferencial dentro de las paleocuevas y de acuerdo a la estructura del sedimento de relleno (Peña, 1997). Si bien estos trabajos son los primeros en abordar el tema y algunos conocimientos han avanzado desde entonces, en ellos no se enfatiza la detección de posibles diferencias métricas o estructurales con las de otros posibles constructores, como Paedotherium sp. Esta realidad evidencia la necesidad de un estudio detallado descriptivo que refleje la morfometría de las paleocuevas, así como la frecuencia morfométrica de las mismas en relación con la frecuencia del material fósil hospedante, que permita obtener una visión más clara de la dinámica, construcción y uso de cuevas en esta fauna fósil.
El objetivo del trabajo es analizar la morfometría de las paleocuevas aflorantes en la "Formación" Chapadmalal teniendo en cuenta posibles diferencias genéricas y específicas en la construcción de las mismas. En particular se pretende: a) analizar la morfometría de las paleocuevas en sección transversal, b) comparar métricamente las paleocuevas asociadas a distintos fósiles hospedantes, c) analizar posibles diferencias temporales y/o espaciales en la morfometría de las paleocuevas y d) interpretar la dinámica y uso de cuevas por la fauna asociada a las mismas.
MATERIALES Y MÉTODOS
Se trabajó en las estaciones de muestreo definidas por Zárate (1989) correspondientes a las secciones Playa San Carlos, Playa Las Palomas (I, II), Playa Los Lobos, La Estafeta (I, III), Arroyo Lobería (I, II) y Punta Vorohué (Fig. 2). Los perfiles se analizaron de acuerdo al esquema estratigráfico propuesto por Zárate (1989). Los niveles muestreados en la estación Playa San Carlos, Playa Las Palomas, Playa Los Lobos, La Estafeta I y Punta Vorohué se corresponden con el Chapadmalalense inferior (P1, P2, P3, P4, P5) y superior (P6), con niveles más antiguos en Playa San Carlos (P1, P2) y Playa Las Palomas (P1, P2, P3), y más modernos en Playa Los Lobos (P2, P3, P4, P6), La Estafeta I (P5) y Punta Vorohué (P4, P5, P6) (Zárate, 1989). Los estratos muestreados de la estación Arroyo Lobería I y II se corresponden a los niveles P10 y P11 propuestos por Zárate (1989) de una antigüedad de 2.5 ma., tiempo que se corresponde con la "Formación" Vorohué (Cione y Tonni, 2001). Los estratos muestreados en la estación La Estafeta (III) se corresponden con el Barrancalobense (P7).

Fig. 2. Estaciones de muestreo definidas por Zárate (modificado de Zárate, 1989). En el trabajo se incluyen las estaciones Playa San Carlos, Playa Las Palomas (I, II), Playa Los Lobos, La Estafeta (I, III), Aº Lobería (I, II) y Punta Vorohué.
Se seleccionaron 370 paleocuevas en sección transversal (Fig. 1) a las que se referenció estratigráficamente. Sobre la sección transversa de las paleocuevas se midió el diámetro lateromedial (LM) y dorsoventral (DV) con cinta métrica (0.05 mm). En los casos en que la sección aflorante de la paleocueva no era transversal pero próxima a ser observada, se removió el material circundante hasta que la sección transversal fue visible. Con las medidas se construyó un índice estructural (Índice de Diámetros Relativo, IDR) calculado como el diámetro dorsoventral dividido el lateromedial (DV/LM).
Para cada paleocueva se registró si contenía o no restos fósiles asociados aflorantes en la sección expuesta y el estado del material (articulado/desarticulado). El material se determinó a nivel de género o especie en los casos en que fue posible. En cada caso, el material fósil asociado a cada paleocueva se consideró como correspondiente al posible taxón constructor de la misma. Diez ejemplares fósiles se registraron en paleocuevas que no pudieron ser medidas por la erosión que presentaban las mismas, sin embargo se consideraron los datos para el análisis de asociación del material fósil a estas bioestructuras. Asimismo se registró una madriguera en planta (Fig. 1) sin asociación a material fósil que se consideró para evaluar las medidas relativas entre la cámara y el túnel. Además se registró la estructura del sedimento de relleno de acuerdo a Peña (1997), clasificándola en caótico, macizo, o laminar.
Las medidas, en forma individual, y el índice IDR fueron analizados estadísticamente (Zar, 1984) mediante distribución de frecuencias, con la finalidad de detectar distintas tendencias métricas en la construcción de las paleocuevas. La normalidad de las distribuciones de frecuencias se testeó por medio de Kolmogorov-Smirnov test (K-S) y se describieron teniendo en cuenta la media, desvío estándar (DS) y moda. Por otro lado, se analizó la diferencia de medias (Student's t test) y distribución de frecuencias de estas variables entre las paleocuevas asociadas a distintos géneros fósiles hospedantes, con la finalidad de detectar posibles diferencias en los tamaños de las paleocuevas de cada especie fósil registrada y el efecto de la existencia de más de un tipo morfométrico de paleocueva en la distribución de frecuencias. Las variables se correlacionaron entre sí para analizar la relación entre las mismas y se analizó la distribución de los residuales (Chambers et al., 1983) con la finalidad de detectar desviaciones de la linearidad y normalidad de los datos que sirvan de indicio para el análisis de las variables y posibles factores asociados (como la existencia de más de un tipo morfométrico de paleocueva). El material fósil se analizó teniendo en cuenta la abundancia de cada género/especie asociada a las paleocuevas, la masa corporal registrada para los géneros/especies encontrados y su correspondencia con el tamaño de las paleocuevas, y los posibles hábitos en el uso de cuevas (constructores, oportunistas).
La masa corporal de los taxones considerados se estiman en 1.3 kg para Actenomys sp. (Fernandez et al., 2000), 1.86 kg para Paedotherium sp. (Elissamburu, 2004), 3 a 8.8 kg. para Lagostomus sp. viviente (Canevari y Vaccaro, 2007), 1.1 kg para Eumysops sp. (Candela, 2005), 286.1 g, oscilando entre 248.2 y 326 g, para Microcavia sp. (Redford y Eisenberg, 1992), y un tamaño similar al de Actenomys sp. para Dolicavia (similar al de Cavia que va de 500 g a 1.5 kg.; Nowak, 1999).
El estado del material fósil y la estructura del sedimento de relleno de las paleocuevas se analizaron descriptivamente. Estas características son indicativas de la muerte in situ del organismo generador del fósil así como del evento de colmatación de la cueva, es decir, si la cueva se rellenó en forma intempestiva o paulatina dando tiempo a abandonar la misma (Peña, 1997).
En una segunda etapa se analizaron comparativamente las características métricas de las paleocuevas entre las distintas unidades de muestreo, con la finalidad de detectar diferencias temporales y/o geográficas en las características estructurales y distribución de las mismas. En este caso se realizaron análisis de varianza (ANOVA) y Tukey Test entre las estaciones de muestreo (Zar, 1984). El material fósil fue comparado entre las distintas estaciones de muestreo sobre la base del número mínimo de materiales asociados a las paleocuevas.
RESULTADOS
Análisis general de las paleocuevas y los fósiles asociados
En la Fig. 3 se muestran las distribuciones de frecuencia para el diámetro dorsoventral (DV), lateromedial (LM) y el índice IDR; los datos descriptivos de estas distribuciones se dan en la Tabla 1.

Fig. 3. Distribución de frecuencias para los diámetros dorsoventral (DV), lateromedial (LM), e índice de diámetros relativo, IDR. Los valores de los diámetros se expresan en cm.
Tabla 1
Estadísticos de las distribuciones de frecuencias de los diámetros dorsoventral (DV) lateromedial (LM) e índice de diámetros relativos (IDR) y diferencia de medias entre las paleocuevas asociadas a Actenomys sp. y Paedotherium sp. Para las distribuciones de frecuencias se indican el número muestral (n), la media, moda, frecuencia de la moda (Frec. moda), mínimo, máximo, rango y desvío estándar (DS). Para Actenomys sp. y Paedotherium sp. se indican entre paréntesis primero el desvío estándar y luego, a renglón seguido, el tamaño muestral. * Diferencia altamente significativa p < 0.01.

La distribución de frecuencias de los diámetros LM (K-S: d = 0.11; p < 0.01) y DV (K-S: d = 0.12; p < 0.01) no se distribuyen normalmente. Dentro de las distribuciones del diámetro DV (media = 15.28 cm, DS = 3.4) y LM (media = 16.6 cm, DS = 3.76) son más frecuentes los valores del diámetro DV comprendidos entre 12 y 16 cm (moda = 15 cm) y los del diámetro LM comprendidos entre 12 y 18 cm con un pico entre 14 y 16 cm (moda = 15 cm). Las frecuencias de los datos disminuyen en forma abrupta para los valores menores a 10 cm (DV) y 12 cm (LM) y presentan un desvío de los datos hacia los valores más altos con una disminución de frecuencias más paulatina en ambos casos. Este comportamiento de los datos, estaría asociado a una superposición de paleocuevas de distinto tamaño, ya sea debido a la existencia de más de un constructor (ver diferencia de medias y distribución de frecuencias entre especies) o a la construcción de cuevas de distinto diámetro para la misma especie, posiblemente asociado a distintos estadios de desarrollo (ver Discusión).
La correlación entre los diámetros LM y DV (r = 0.81), cuando se analiza la distribución de cada variable frente a la distribución normal esperada, muestra que los datos se apartan de lo esperado hacia los menores y mayores valores y que la dispersión de los mismos incrementa hacia los valores más elevados del diámetro LM.
El índice es muy poco variable (media = 0.9; DS = 0.1; n = 332) y se distribuye normalmente (K-S: d = 0.069; p < 0.1), con predominio de valores que indican paleocuevas prácticamente circulares. La correlación entre el índice IDR y las variables es muy baja (IDR-DV: r = 0.3; IDR-LM: r = -0.3). Sin embargo, en la correlación entre el índice y cada variable se puede observar que los valores más bajos del índice se apartan levemente de lo esperado. Estos valores se corresponden con paleocuevas ovales que presentan prácticamente el doble de diámetro LM que DV y constituyen un punto interesante de análisis en la interpretación de la construcción de las mismas (ver Discusión).
En la Tabla 1 y Fig. 4 se muestran los valores medios de las variables DV, LM y del índice IDR para las paleocuevas asociadas a A. priscus y Paedotherium sp. La diferencia de medias entre los diámetros de las paleocuevas para estas especies es altamente significativa, con paleocuevas de mayor diámetro asociadas a Paedotherium sp. No sucede lo mismo con el valor del índice, que no muestra diferencias significativas entre ambas especies. En el caso de A. priscus, en dos de las paleocuevas de menor diámetro (diámetros de 10 cm en un caso y de 11 cm en el otro) se encontraron asociados ejemplares juveniles. En el caso de Paedotherium sp., las dos especies reconocidas para el Plio-Pleistoceno (Cerdeño y Bond, 1998), P. typicum y P. bonaerense, se registraron en las paleocuevas pero con una mayor frecuencia de P. bonaerense que P. typicum (5:1).
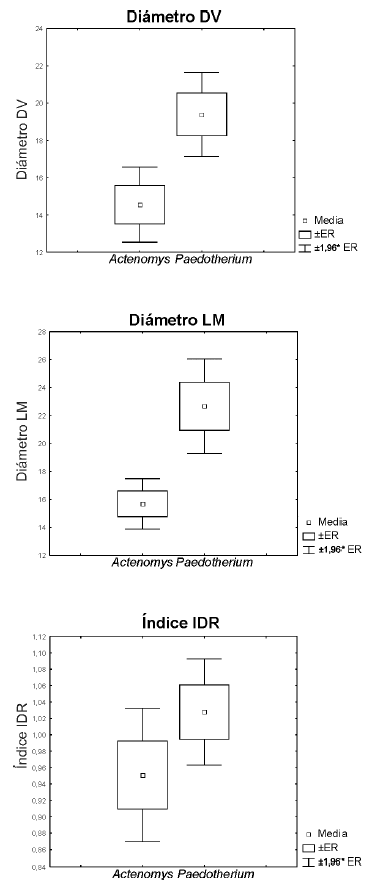
Fig. 4. Distribución de medias para las paleocuevas asociadas a Actenomys sp. y Paedotherium sp. Los valores de los diámetros se expresan en cm. ER: error estándar.
La distribución de frecuencias conjunta de las paleocuevas asociadas a estas dos especies presenta una tendencia similar a lo observado en el total de paleocuevas analizadas, ejemplificando el efecto que causa la superposición de paleocuevas de distintos tamaños. Las diferencias de medias y las distribuciones de frecuencia avalan la presencia de al menos tres tipos morfométricos de paleocuevas: paleocuevas de 10 a 18 cm asociadas a A. priscus, entre las que habría paleocuevas de 11 cm o menores asociadas a juveniles, y paleocuevas de 16 a 22 cm asociadas a Paedotherium sp.
Para los restantes géneros asociados a las paleocuevas (Lagostomopsis sp., Eumysops sp., Dolicavia sp., Caviidae indet.) los datos son escasos como para comparar estadísticamente las diferencias métricas. Sin embargo, es relevante notar que entre los géneros asociados se encuentran principalmente Lagostomopsis sp., Eumysops sp. y cávidos de tamaño similar o menor al de Actenomys sp. y que, de acuerdo a lo observado en las especies vivientes relacionadas y trabajos previos, construirían cuevas (Lagostomopsis sp.; Genise, 1989) o tendrían una construcción probable de las mismas (Eumysops sp., Dolicavia sp., Caviidae indet.), por lo que se plantea la existencia de cuevas correspondientes a alguno de estos géneros y un uso posiblemente oportunista en otros. Las paleocuevas asociadas a Lagostomopsis sp. tienen valores métricos comprendidos entre los de A. priscus y Paedotherium sp. Las dos paleocuevas asociadas a Eumysops sp. presentan valores muy dispares, con diámetros de 13 y 26 cm respectivamente, lo cual podría indicar un uso oportunista de las mismas. Los cávidos se asocian a paleocuevas comprendidas dentro del rango métrico de las de A. priscus.
Por otro lado, en la madriguera registrada en planta (Fig. 1), la cámara presenta un diámetro LM de 30 y 32.5 cm, casi circular, y el túnel un diámetro LM de 15 cm.
En relación con el relleno de las paleocuevas (macizo/caótico/laminar), predominó el relleno macizo tanto en las paleocuevas asociadas a material fósil (59:1:1) como en las que no presentaron restos asociados (255:5:43). Se registraron rellenos mixtos entre macizo y laminar en seis paleocuevas, una de ellas asociada a material fósil. Por otro lado, la proporción de restos articulados frente a los no articulados fue del 21.8% (12:43).
Análisis espacial de las paleocuevas y los fósiles asociados
En la Tabla 2 se indican los valores medios para los diámetros LM y DV entre las distintas estaciones de muestreo. Los diámetros medios de las paleocuevas fueron menores (ANOVA: DV: F = 7.48, df = 327, p < 0.001, LM: F = 8.82, df = 322, p < 0.001; Tukey test p < 0.05) en la estación Playa San Carlos, Playa Las Palomas y Playa Los Lobos, diferenciándose de los valores de las paleocuevas de Arroyo Lobería II.
Tabla 2
Diferencia de medias entre las estaciones de muestreo. Entre paréntesis se indican primero el desvío estándar y a renglón seguido, tamaño muestral. Playas San Carlos, Las Palomas y Los Lobos se diferencian significativamente de Arroyo Lobería II (p < 0.05).

De acuerdo con la estratigrafía de las distintas estaciones de muestreo, las paleocuevas son en promedio de menor diámetro en los sedimentos del P1, P2, P3, P4 y P6 del Chapadmalalense inferior y superior aflorantes en Playa San Carlos, Playa Las Palomas y Playa Los Lobos, difiriendo hacia paleocuevas de mayor diámetro en la estación Aº Lobería II correpondientes a la "Formación" Vorohué (nivel P11). Los diámetros de las paleocuevas de la "Formación" Vorohué tienen valores más próximos a los registrados para las paleocuevas de Paedotherium sp.
En relación con la asociación de Actenomys sp., Paedotherium sp. y otros restos a las paleocuevas, en Playa San Carlos fue del 0.60% para Actenomys sp. y 0.40 % para Paedotherium sp. (3:2). En Playa Las Palomas los restos asociados a paleocuevas corresponden a 64.71% de Actenomys sp. 23.53% de Paedotherium sp. y de 11.76% de cávidos (11:4:2). En Playa Los Lobos el 44.44% corresponde a Actenomys sp., el 11.12% a Paedotherium sp. y el 44.44% a otros géneros (Eumysops sp., Lagostomopsis sp., cavidos) (4:1:4). En Arroyo Lobería II se registraron Paedotherium sp. (66.66%) y Eumysops sp. (33.34%) (2:1). Cabe aclarar que la densidad de cuevas (transversales) en las estaciones de muestreo es mucho mayor en Playa Los Lobos (215 cuevas/km) y Playa Las Palomas (190 cuevas/km) que en Playa San Carlos (37.9 cuevas/km) y Arroyo Lobería II (30.9 cuevas/km).
Análisis general de las paleocuevas y los fósiles asociados
La distribución de los diámetros de las cuevas de individuos adultos, de acuerdo a lo esperado, debería aproximarse a una distribución normal, con los límites máximo y mínimo de la distribución impuestos por el tamaño corporal del constructor. Esto plantea que los valores morfométricos de las paleocuevas estén acotados dentro de un rango de variación reducido, concordante con la variación de tamaño de los individuos. Por ejemplo, en el roedor actual Ctenomys mendocinus, el tamaño de las cuevas es bastante constante, con un diámetro transverso de 8.03 cm (SD:1.72) y 8.5 cm (SD:1.51), para machos y hembras respectivamente, y con un rango de masa corporal que va de los 140 a 260 g (Rosi et al., 1996).
Las distribuciones de frecuencia de los diámetros lateromedial y dorsoventral de las paleocuevas estudiadas muestran que la distribución no se corresponde con lo esperado para un solo tipo morfométrico de paleocueva. No sólo la distribución no es normal, sino que los límites de la distribución para el diámetro dorsoventral (5.5-28.5 cm) y lateromedial (7.5-33.5 cm) representan un rango métrico demasiado amplio (23 y 26 cm, respectivamente) como para que puedan corresponderse a un solo tipo de tamaño corporal de constructor, o a un mismo esquema de construcción de cuevas dado un tamaño corporal similar. La distribución no normal, con la caída abrupta hacia los valores más bajos y la disminución de frecuencias más paulatina hacia los valores más elevados, hacen que la distribución de frecuencias se adapte mejor a un esquema de más de un tipo métrico de paleocuevas superpuestos, que en su conjunto conforman la distribución observada.
La asociación de los juveniles de A. priscus a las paleocuevas de menor diámetro avala una diferencia etaria en la morfometría de las paleocuevas para esta especie. Si bien se trata de dos observaciones, es significativo a nivel paleontológico. La diferencia de medias y la distribución de frecuencias de las paleocuevas asociadas a Actenomys sp. y Paedotherium sp. respaldan la distribución general observada y la existencia de al menos tres tipos morfométricos de paleocuevas: paleocuevas de 10 a 18 cm asociadas a A. priscus, entre las que se diferencian paleocuevas de 11 cm o menores asociadas a juveniles, y paleocuevas de 16 a 22 cm asociadas a Paedotherium sp. El diámetro de las paleocuevas reconocido para A. priscus incluye al descripto por Genise (1989) para las paleocuevas de esta especie (13 cm). Esta superposición de tamaños de cuevas y tipos de constructores hace dificultosa la diferenciación a simple vista de las cuevas referidas a la "Formación" Chapadmalal.
Similar a lo que sucede en las especies actuales del roedor Ctenomys sp. (Malizia et al., 1994), la construcción de las cuevas para Actenomys comenzaría en la etapa juvenil, condición que permite comprender en más detalle el comportamiento de esta especie fósil. En las especies actuales, los juveniles comienzan la construcción de sus cuevas a temprana edad, apartándose de la cueva parental en etapas muy tempranas (Malizia et al., 1994), por lo que resulta esperable encontrar cuevas de menor diámetro asociadas a esta etapa de desarrollo. En el caso particular de Paedotherium no se encontraron asociados individuos juveniles a las paleocuevas. Entre las especies de Paedotherium, los ejemplares determinados respaldan una mayor asociación con las paleocuevas en P. bonaerensis. Ambas especies presentan capacidad para cavar, sin embargo la estructura del esqueleto apendicular de P. bonaerense respalda un mayor desarrollo de fuerza y posiblemente una mayor adaptación cavadora en esta especie (Elissamburu, 2004; Elissamburu y Vizcaíno, 2005). Esto supone posibles diferencias paleoecológicas y comportamentales entre ambas especies en relación con el uso de cuevas.
Respecto a los diámetros de las paleocuevas, hay una mayor variación en el diámetro lateromedial que el dorsoventral. Esto plantea, por un lado, que el diámetro dorsoventral es más representativo del tamaño del constructor de la cueva, y por otro que el diámetro lateromedial de las cuevas era fluctuante, con predominio de valores similares al del diámetro dorsoventral pero con incrementos que, en los casos extremos, llegaban casi a duplicar el alto de la cueva. El ensanchamiento del túnel correspondiente puede estar asociado tanto al tránsito simultáneo de más de un individuo en el mismo, como a ensanchamientos similares a los que se observan actualmente en las cuevas de Ctenomys sp. asociados a los puntos de bifurcación del túnel principal (Rosi et al., 1996). En el caso particular de las cuevas que duplican el diámetro en sentido lateromedial, podrían referirse, además, a madrigueras. La madriguera registrada en planta es de forma prácticamente circular, con el diámetro lateromedial de la cámara que duplicaba al del túnel, por lo que se mantienen las medidas proporcionales citadas. Las cámaras reconocidas por Genise (1989) son de forma ovoide en la sección aflorante (que no coincide necesariamente con la sección transversa de la cámara), con un largo máximo de 33.1 cm y un ancho máximo de 19.5 cm, sin embargo estas medidas no se corresponden exactamente con el diámetro dorsoventral y transverso de la cámara, sino con los diámetros máximos observados, por lo que pueden estar representando las mismas madrigueras referidas pero vistas en otra sección.
La presencia de otras especies asociadas a las paleocuevas se debe a la existencia de otras especies cavadoras constructores de cuevas, como Lagostomopsis sp., y otras posibles constructoras u oportunistas en el uso de las mismas, que hacen más compleja la interpretación de la dinámica en el uso de cuevas y diferenciación de estas estructuras. En el caso particular de Lagostomopsis sp., es constructor de cuevas (Genise, 1989) y es muy probable que las cuevas a las que se encontró asociado le pertenezcan, aunque aún faltan registros para poder diferenciar métricamente las paleocuevas de esta especie, dada su baja frecuencia de aparición. Los datos del presente trabajo se corresponden con una paleocueva circular asociada a un juvenil y a una paleocueva en la que sólo pudo medirse el diámetro lateromedial, por lo que no es posible inferir el tamaño de las mismas, ni la forma transversal asociada a ejemplares adultos, aunque es esperable que en promedio sean mayores a las de A. priscus (Genise, 1989) dado que es un género que duplica el tamaño corporal de esta especie (3 a 8.8 kg; Canevari y Vaccaro, 2007).
Eumysops sp. habría tenido hábitos epigeos (Olivares, 2009) y los registros hasta el momento se encuentran tanto fuera (Genise, 1989) como dentro (en el presente trabajo y en Peña, 1997) de las paleocuevas, en muy baja frecuencia. Además el tamaño dispar de las paleocuevas a las que se encuentra asociado hace pensar en un hábito oportunista en el uso de las mismas. El tamaño corporal, estimado en 1.1 kg (Candela, 2005) es similar al estimado para A. priscus, por lo que en el caso de construir cuevas, éstas serían morfométricamente semejantes a las registradas para A. priscus adultos.
Para Dolicavia sp. se ha propuesto un uso periférico de las cuevas de Actenomys sp. y convivencia conjunta (Genise, 1989), basándose en la asociación de restos de estos dos géneros a una misma paleocueva. Esta hipótesis no puede respaldarse con los datos disponibles en el presente trabajo, no obstante, la existencia futura de nuevos registros de la preservación conjunta de ambos géneros asociados a paleocuevas puede llegar a ser un indicio de este comportamiento. El tamaño corporal de Dolicavia sp. sería similar al de A. priscus, por lo que el tamaño de las cuevas sería semejante. Entre los Caviidae vivientes, por otro lado, se registra un uso compartido de cuevas y convivencia conjunta entre géneros como Microcavia australis y Galea musteloides (Canevari y Vaccaro, 2007), aunque ambos son gregarios.
Teniendo en cuenta los datos de trabajos previos, también debe considerarse la asociación de Microcavia sp. a las paleocuevas (Peña, 1997), con un posible uso oportunista o con cuevas propias de menor tamaño que las de A. priscus, dado el tamaño corporal bastante menor al de A. priscus (286.1 g, oscilando entre 248.2 y 326 g; Redford y Eisenberg, 1992).
Por otro lado, el tipo de relleno, predominantemente macizo, indica que la colmatación de las cuevas fue en general rápida, no dando tiempo al escape de los organismos habitantes (Peña, 1997). Dada la proporción de restos articulados, la proporción de muerte in situ es alta, con escaso transporte del material. Esta condición tafonómica favorece el estudio de las paleocuevas en relación con sus fósiles hospedantes, siendo elevado el grado de asociación entre las paleocuevas con sus constructores u habitantes oportunistas.
Análisis espacial de las paleocuevas y los fósiles asociados
Las características de las paleocuevas de los yacimientos correspondientes al P1, P2, P3, P4 y P6 del Chapadmalalense inferior y superior de las estaciones Playa San Carlos, Playa Las Palomas y Playa Los Lobos, de menor tamaño, con predominio de restos fósiles de A. priscus, y mayor densidad en el número de paleocuevas (principalmente en Playa Las Palomas y Playa Los Lobos), contrastan con las paleocuevas de mayor tamaño, menor densidad en el número de las mismas y predominio de restos de Paedotherium sp. registradas en el Vorohuense, aflorantes en la estación Arroyo Lobería II. El tamaño de las paleocuevas de la "Formación" Vorohué se corresponde con el registrado para las paleocuevas de Paedotherium sp., por lo que podría tratarse de la especie predominante en la zona.
Esta diferencia en el tamaño general de las paleocuevas posiblemente se deba a una diferencia en la abundancia relativa de A. priscus y Paedotherium sp. entre estos dos estadios de tiempo. Estas diferencias temporales en el tamaño de las paleocuevas plantean la existencia de diferencias de abundancia relativa entre ambas especies, con un predominio de A. priscus en los niveles más antiguos del Chapadmalalense, en la zona de Playa San Carlos, Playa Las Palomas y Playa Los Lobos, y predominio de Paedotherium sp. en la "Formación" Vorohué, en la zona de Aº Lobería.
CONCLUSIONES
Las características morfométricas de la sección transversal de las paleocuevas observadas y los análisis descriptivos realizados permiten inferir la existencia de al menos tres rangos crecientes de tamaños de paleocuevas, correspondientes a cuevas construidas por A. priscus, en estadios juveniles y adultos, y a Paedotherium sp. respectivamente.
Se reconocen además diferencias temporales en el tamaño de las paleocuevas, que se asocian con variaciones de abundancia relativa en la distribución de ambas especies. Las paleocuevas de A. priscus predominan en el Chapadmalalense, mientras que las de Paedotherium predominan en la "Formación" Vorohué.
Los géneros registrados en asociación con las paleocuevas plantean una realidad compleja en relación con la construcción y uso de las mismas y en la dinámica de cuevas en el Chapadmalalense y en la "Formación" Vorohué, con la presencia conjunta de al menos tres constructores de cuevas (A. priscus, Paedotherium sp. y Lagostomopsis sp.) y especies oportunistas en el uso de cuevas ya construidas por los géneros citados, o posibles constructoras (Eumysops sp., Dolicavia sp., caviidae indet.).
AGRADECIMIENTOS
A los revisores por las correcciones y sugerencias. El presente trabajo fue financiado por CONICET.
LITERATURA CITADA
1. AMEGHINO F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal. Anales del Museo Nacional de Buenos Aires 10(3):343-428. [ Links ]
2. CANDELA AM. 2005. Los roedores del "Mesopotamiense" (Mioceno tardío, Formación Ituzaingó) de la provincia de Entre Ríos (Argentina). INSUGEO Miscelánea 14:37-48. [ Links ]
3. CANEVARI M y O VACCARO. 2007. Guía de Mamíferos del Sur de América del Sur. Editorial L.O.L.A. Buenos Aires, Argentina. [ Links ]
4. CERDEÑO E y M BOND. 1998. Taxonomic revision and phylogeny of Paedotherium and Tremacyllus (Pachyrukhinae, Hegetotheriidae, Notoungulata) from the Late Miocene to the Pleistocene of Argentina. Journal of the Vertebrate Paleontology 18:799-811. [ Links ]
5. CHAMBERS JM, WS CLEVELAND, B KLEINTER y PA TUKEY. 1983. Graphical methods for data analysis. Wadsworth y Brooks/Cole, Pacific Grove, California. [ Links ]
6. CIONE AL y EP TONNI. 1995a. Los Estratotipos de los pisos Montehermosense y Chapadmalalense (Plioceno) del esquema cronológico sudamericano. Ameghiniana 32:369-374. [ Links ]
7. CIONE AL y EP TONNI. 1995b. Chronostratigraphy and "Land-Mammal Ages" in the Cenozoic of Southern South America: principles, practices, and the "Uquian" problem. Journal of Paleontology 69:135-159. [ Links ]
8. CIONE AL y EP TONNI. 2001. Correlation of Pliocene to Holocene southern South American and European Vertebrate-Bearing units. Bolletino della Società Paleontologica Italiana 40(2):167-173. [ Links ]
9. ELISSAMBURU A. 2001. Análisis morfométrico y morfofuncional del esqueleto apendicular de Paedotherium (Mammalia, Notoungulata) del Plioceno y Pleistoceno Temprano de la provincia de Buenos Aires. Correlación con su aptitud fosorial. Tesis de Licenciatura en Ciencias Biológicas inédita, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Mar del Plata, Argentina. [ Links ]
10. ELISSAMBURU A. 2004. Análisis morfométrico y morfofuncional del esqueleto apendicular de Paedotherium (Mammalia, Notoungulata). Ameghiniana 41(3):363-380. [ Links ]
11. ELISSAMBURU A y SF VIZCAÍNO. 2005. Diferenciación morfométrica del húmero y fémur de las especies de Paedotherium (Mammalia, Notoungulata) del Plioceno y Pleistoceno temprano. Ameghiniana 42(1):159-166. [ Links ]
12. FERNÁNDEZ ME, AI VASSALLO y MA ZÁRATE. 2000. Functional Morphology and Paleobiology of the Pliocene rodent Actenomys (Caviomorpha:Octodontidae): the evolution to a subterranean mode of life. Biological Journal of the Linnean Society 71(1):71-90. [ Links ]
13. FRENGUELLI J. 1921. Los terrenos de la costa atlántica de los alrededores de Miramar y sus correlaciones. Boletín Academia Nacional de Ciencias, Córdoba 24:325-428. [ Links ]
14. GENISE JF. 1989. Las cuevas con Actenomys (Rodentia, Octodontidae) de la formación Chapadmalal (Plioceno Superior) de Mar del Plata y Miramar (Provincia de Buenos Aires). Ameghiniana 26(1-2):33-42. [ Links ]
15. KRAGLIEVICH J. 1952. El perfil geológico de Chapadmalal y Miramar, provincia de Buenos Aires. Revista Museo Municipal de Ciencias Naturales y Tradicional Mar del Plata 1:8-37. [ Links ]
16. MALIZIA AI, RR ZENUTO y C BUSCH. 1994. Demographic and reproductive attributes of disperser in two populations of the subterranean rodent Ctenomys talarum (tuco-tuco). Canadian Journal of Zoology 73:732-738. [ Links ]
17. NOWAK RM. 1999. Walker´s Mammals of the World. 6th ed. The Johns Hopkins University Press, Baltimore. [ Links ]
18. OLIVARES AI. 2009. Anatomía, sistemática y evolución de los roedores caviomorfos sudamericanos del género Eumysops Ameghino [Rodentia, Echimyidae]. Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Argentina. [ Links ]
19. PEÑA MI. 1997. Tafonomía de restos óseos y condiciones ambientales del Plioceno Tardío, Playa San Carlos (Pdo. de Gral. Pueyrredón). Tesis de Licenciatura en Ciencias Biológicas inédita, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Mar del Plata, Argentina. [ Links ]
20. REDFORD KH y JF EISENBERG. 1992. Mammals on the Neotropics. The Southern Cone. Vol. 2. The University of Chicago Press, Chicago, USA. [ Links ]
21. ROSI MI, MI CONA, S PUIG, F VIDELA y VG ROIG. 1996. Size and structure burrow systems of the fossorial rodent Ctenomys mendocinus in the piedemont of Mendoza province, Argentina. Z. Säugetierkunde 61:352-364. [ Links ]
22. SCOGNAMILLO DG. 1993. Estructuras de las cuevas de Actenomys (Rodentia: Octodontidae) de la Aloformación Playa San Carlos, Plioceno tardío (Barranca de los Lobos, Pdo. Gral. Pueyrredón): significado paleoecológico y estratigráfico. Tesis de Licenciatura en Ciencias Biológicas inédita, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Mar del Plata, Argentina. [ Links ]
23. VERZI DH. 2002. Patrones de evolución morfológica en Ctenomyinae (Rodentia, Octodontidae). Mastozoología Neotropical 9(2):309-328. [ Links ]
24. ZAR JH. 1984. Biostatistical Analysis. Prentice-Hall, INC., Englewood Cliffs, New Jersey. [ Links ]
25. ZÁRATE MA. 1989. Estratigrafía y geología del Cenozoico Tardío en los acantilados marinos comprendidos entre Playa San Carlos y Aº Chapadmalal. Partido de General Pueyrredón. Prov. De Buenos Aires. Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Argentina. [ Links ]
Recibido 18 enero 2011.
Aceptado 13 abril 2011.
Editor asociado: F Prevosti

