










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkMastozoología neotropical
Print version ISSN 0327-9383
Mastozool. neotrop. vol.19 no.1 Mendoza June 2012
ARTÍCULOS Y NOTAS
Small mammal commnities in the Sierra de los cuchumatanes, Huehuetenago, Guatemala
John O. Matson1, Nicté Ordóñez-Garza2, Walter Bulmer3, and Ralph P. Eckerlin3
1 Department of Biological Sciences, San Jose State University, San Jose, CA 95192-0100 [Correspondence: <johnomatson@gmail.com>].
2 Department of Biological Sciences, Texas Tech University, Lubbock, TX 79409-3131.
3 Division of Natural Sciences and Mathematics, Northern Virginia Community College, Annandale, VA 22003-3796.
Recibido 4 octubre 2011.
Aceptado 23 febrero 2012.
Editor asociado: T Escalante
ABSTRACT: Very little is known concerning small mammal ecology and their distribution in the highlands of Guatemala. Small mammals were trapped from five different cloud forests in the Sierra de los Cuchumatanes, Huehuetenango, Guatemala. Cloud forest elevations ranged from 2600 m to 3350 m. Most sites had evidence of human disturbance with only Cerro Bobí having a relatively pristine forest. A total of 13 species was recorded with the species density at sites ranging from six to ten. Accounts of each species are given. The most common species present was Reithrodontomys sumichrasti, except at Cerro Bobí where Peromyscus guatemalensis was most abundant. Differences between years and between seasons (wet versus dry) were minor. Only the site at Cerro Bobí, with relatively pristine cloud forest, appeared to have a species composition that resembled other cloud forests in the highlands of Guatemala. Information on reproduction is summarized.
RESUMEN: Comunidades de pequeños mamíferos en la Sierra de los Cuchumatanes, Huehuetenango, Guatemala. Se sabe muy poco sobre la ecología y distribución de micromamíferos en las tierras altas de Guatemala. Los micromamíferos fueron capturados en cinco diferentes bosques nubosos de la Sierra de los Cuchumatanes, Huehuetenango, Guatemala. Las elevaciones de los sitios de muestreo oscilaron entre los 2600 y los 3350 m. La mayoría de los sitios tenían evidencia de perturbación humana, excepto por Cerro Bobí que tenía un bosque relativamente prístino. Se registraron 13 especies con densidad de especies entre sitios de seis a diez. Las especies más comunes presentes fueron Reithrodontomys sumichrasti, excepto en el Cerro Bobí, donde Peromyscus guatemalensis fue la especie más abundante. Las diferencias entre años y entre estaciones (húmeda versus seca) no fueron significativas. Sólo el sitio en el Cerro Bobí, con bosque nuboso relativamente prístino, parece tener una composición de especies similar a otros bosques nubosos de tierras altas de Guatemala. Adicionalmente, se presentan datos sobre las especies capturadas, incluyendo información reproductiva.
Key words: Cloud forests; Ecology; Mesoamerica; Rodents; Shrews.
Palabras clave: Bosque nublado; Ecología; Mesoamérica; Musaraña; Roedores.
INTRODUCTION
Few studies have been conducted on terrestrial small mammals from Neotropical Montane (Cloud) Forests. Most studies have concentrated on altitudinal gradients of species diversity (Patterson, et al., 1989; Patterson et al., 1998; Vargas-Contreras and Hernández-Huerta, 2001; Sanchez-Cordero, 2001; Monteagudo-Sabaté and León-Paniagua, 2002; McCain, 2004). With the exception of a few studies in Mexico (e.g. Rickart, 1977; Vázquez et al., 2000, 2004; Castro-Arellano, 2005, 2006), basic ecological data such as population structure, species diversity, and life history information are generally lacking for montane forest areas. Castro-Arellano (2005, 2006) studied small mammal communities in the northernmost cloud forest in the Neotropics, Tamaulipas, México. Castro-Arellano (2005) and Vázquez et al. (2000) are the only studies which address, in detail, the community structure of small mammals in montane forests. Rickart (1977) studied reproduction in two species of cloud forest Peromyscus. Some life history information is also available for selected mammal species (Janzen, 1983 and papers therein; Timm et al., 1989; Timm and LaVal, 2000; and Vázquez et al., 2004).
Montane forests (cloud forests) of Guatemala, indeed most montane forests throughout Central America, are threatened by on-going human activity. The fragility of montane forests, their importance in providing watershed and valuable habitat for floral and faunal elements (Doumenge et al., 1995), and the fact that the entire region is listed as one of the World's biodiversity hotspots (Myers et al., 2000), begs for a more complete understanding of their biota. Human activity, especially timber and agricultural activities are rapidly destroying these fragile habitats (Hamilton et al., 1995). Because the Central American highlands are rapidly being exploited, especially in the northern regions, such as Chiapas, Mexico (Cayuela et al., 2006), they are considered a priority area for conservation (Wege and Long, 1995).
The abundance and fine-scale distributions of mammals in the highland forests of Guatemala remain poorly known. As part of a long-term effort to better determine the communities of small mammals inhabiting highland habitats in this region, we surveyed the remnant mixed hardwood/coniferous cloud forests at elevations ranging from 2600 m to 3350 m in the Sierra de los Cuchumatanes, Guatemala.
Prior to this study, few works have been published related to small mammals from this mountain range. Goldman (1951) and Woodman (2010) summarized the collecting that occurred in the late 1800's under the auspices of the Biological Survey and the subsequent taxonomic papers that followed. Carleton and Huckaby (1975) described Peromyscus mayensis from the Sierra, and provided a list of small mammals associated with that species. Recently, Ordóñez-Garza et al. (2010) and Woodman (2010) have presented studies that help to clarify the taxonomic status of several species of small mammals that occur in the region, the latter publication included the descriptions of two new species of short-eared shrew (genus Cryptotis).
Because of the paucity of information on montane forest mammals and the precarious nature of these forests, we present some ecological information on small mammals from several highland (montane) forests in the Sierra de los Cuchumatanes, Guatemala.
STUDY AREAS
The Sierra de los Cuchumatanes is an immense limestone massif that covers a large area of northern Nuclear Central America (sensu Savage, 1982; Donnelly, 1989; Donnelly et al. 1990; Campbell, 1999) in Guatemala. The Sierra de los Cuchumatanes is the highest, largest, and most diverse geologic unit in Northern Central America (Anderson et al., 1973). The mountain range is composed of sedimentary rocks, mostly limestone (Anderson et al., 1973; Donnelly et al., 1990), bounded on the south by the Chixoy-Polochic fault zone and to the north and northeast by a series of step faults that reach into southern Chiapas, Mexico (Anderson et al., 1973).
While our sampling included seven localities over the years 2004-2008, only five had sufficient trapping effort, at least 300 trap nights, to warrant analysis. All study areas were in the Province of Huehuetenango (Fig. 1). Tree species listed were determined using Parker (2008).
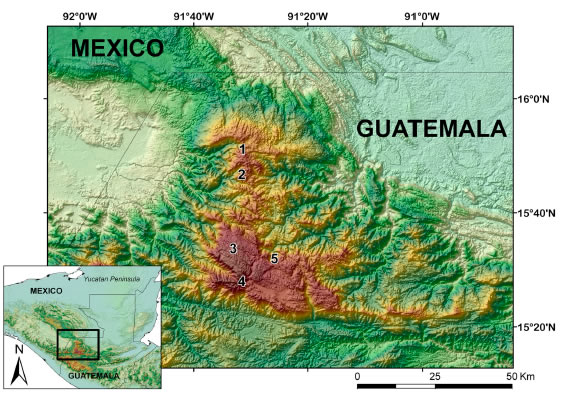
Fig. 1. Map depicting the collecting localities at Cuchumatanes, Guatemala. 1) Cerro Bobí. 2) Yaiquich. 3) Puerta del Cielo. 4) Aldea El Rancho. 5) Laguna Magdalena.
Aldea El Rancho
This site was 6.5 km SE Todos Santos Cuchumatán, Aldea el Rancho, 3020 m, Reserva Forestal Todos Santos Cuchumatán, (15° 29' 22" N, 91° 33' 16" W). We sampled this site on two occasions, both in the dry season (30 December 2004 through 2 January 2005 and 5-8 January 2008). The weather was cold, frost formed on the ground each morning. Maximum temperatures during the day averaged 17°C, while low temperatures averaged 2°C. A "SPER" Scientific max-min thermometer was placed in the shade at 1.5 m above the ground for all temperature recordings. Maximum and minimum temperatures of the previous day and night were recorded each morning.
The site was located on a west facing slope, in a small valley leading east, up the mountain side from the main dirt road that leads to Todos Santos. The small community of El Rancho was down slope from the road. The locality was along the same ridge, but farther south, that Goldman (1951) described for his Todos Santos locality.
The site was a coniferous cloud forest composed of several species of conifers, including pines (Pinus oaxacana and P. hartwegii), Guatemala fir (Abies guatemalensis), and Mexican cypress (Cupressus lusitanica) with a few broad-leaved trees scattered about, such as oak (Quercus sp.) and monkey hand tree (Chiranthodendron pentadactylon). A variety of bushes dotted the area, but were not identified. Ground cover along the base of limestone cliffs consisted of several grasses and a thick layer (to 20 cm) of moss. In other areas the ground cover was more open with little vegetation. A small, ephemeral stream was located in the valley. During the course of the study, only small pools of water were available.
Our trap-lines were set along the stream, up the forested slopes, and at the base of the limestone cliffs. In 2004/2005 we recorded 600 trap nights and 76 pitfall nights. In 2008 we recorded 863 trap nights and 247 pitfall nights. Thus, we recorded 1463 trap nights and 323 pitfall nights. See Methods below for explanation of trap nights and pitfall nights.
Even though the area is protected, there was evidence of lumbering activity on both occasions that we visited.
Puerta del Cielo
This site was 5 km N Todos Santos Cuchumatán, Aldea Puerta del Cielo, 3350 m, Reserva Forestal Todos Santos Cuchumatán, (15° 33' 16" N, 91° 36' 00" W). We sampled this site on two occasions (2-5 January 2005 and 16- 17 July 2008). The 2005 visit was during the dry season while the 2008 visit was during the wet season. Maximum temperatures averaged 16 °C, and lows were 1 °C during the dry season. We did not measure the temperatures during the wet season. During the times we visited there was almost a constant wind, sometimes estimated at 60+ kph.
The coniferous cloud forest was composed almost exclusively of Guatemala fir with a few scattered pines and a thick (30 cm) ground cover of moss. This east-facing slope was quite steep being at least 60°. Again, we saw evidence of lumbering activity. The site was located just over the ridge north of Todos Santos. It was on the eastern slope of the same mountain ridge as El Rancho. The site is at the top of a pass leading down slope, north to the town of Jacaltenango. We set our trap-lines along this east-facing slope. We recorded 600 trap nights and 50 pitfall nights in 2005, while in 2008 we had only 109 and 20, respectively. A total of 709 trap nights and 70 pitfall nights was recorded.
Laguna Magdalena
This site was 22 km NNE Chiantla, Laguna Magdalena, 2925 m, (15° 32' 06" N, 91° 23' 33" W). A dirt road from Aldea Paquix leads east for about 18 km to the laguna. We visited this site twice (5-4 January 2006 and 9-11 January 2008), both times were during the dry season. Maximum temperatures averaged 16 °C, lows averaged 2 °C. Frost on the ground in the morning was common.
When we sampled the site in 2006, there was a lush growth of grasses along the stream and in the meadows leading to the picturesque lagoon. By 2008 the area had been badly overgrazed by sheep, goats, and cattle.
The mountains surrounding Laguna Magdalena were covered in coniferous cloud forest, somewhat similar to those described for El Rancho and Puerta del Cielo. There is a small stream that emerges from the hillside upslope to the southwest that feeds the lagoon which is about 3 km northeast. In 2006, we watched as men took several mule-loads of large cypress timbers out of the forest. A bundle of at least 40 planks (20 cm x 20 cm x 4 m) was stacked next to our camp waiting to be transported by truck. In 2008, we did not see this activity, but the surrounding slopes had far fewer trees than we saw in 2006. One of us (NOG) had visited this locality several years earlier and remembers heavily forested slopes surrounding Laguna Magdalena.
Our trap-lines were set along the forested slopes and in the flat grassy meadows. In 2006 we recorded 175 trap nights and 20 pitfall nights. In 2008 we recorded 260 trap nights and 48 pitfall nights for a total of 435 and 68, respectively.
Cerro Bobí
This site is 5 km SW San Mateo Ixtatán, Cerro Bobí, 3110 m, (15° 48' 36" N, 91° 30' 19" W). We sampled this site two times, 26-30 July 2005 and 29-31 December/1-2 January 2006. The former sampling period was during the wet season while the latter was during the dry season. Maximum temperatures during the wet season averaged 20 °C while lows averaged 3 °C. During the dry season the highs were 15 °C while lows were 1 °C. During the dry season ground and grasses were often covered with frost in the mornings. Woodman (2010) records this site as the type locality for Cryptotis lacerotosus.
The forest on Cerro Bobí is a remnant of primary hardwood/coniferous cloud forest. Areas around the base of the mountain have been heavily logged, but the forest where we trapped was relatively pristine with a closed canopy. The forest is composed of several broad-leaved hardwoods, such as oak (Quercus sp.), wild avocado (Persia vesticula), and monkey hand tree. Conifers included the Guatemalan podocarp (Podocarpus guatemalensis), various pines, Guatemalan fir, and Mexican cypress. Tree ferns (Cyathea sp.) were numerous. Ground cover consisted of a heavy litter of fallen leaves, moss, lycopodium, and ferns. Fallen logs were numerous on the forest floor. A grassy meadow occurred in a flat area along an ephemeral stream.
Our camp and trap-lines were on the northwest slope of the mountain in the forest and meadow. During the dry season of 2005/2006 we recorded 835 trap nights and 145 pitfall nights. In the wet season of 2005 we used 678 trap nights and 100 pitfall nights. A total trapping effort of 1513 trap nights and 245 pitfall nights was recorded for this site.
Apparently, illegal lumbering occurs in the area. One evening we heard a large truck drive into the area above our camp, maybe 2 km distant. We heard chain saws and falling trees. The truck left before sunrise.
Yaiquich
This site was 6 km NW Santa Eulalia, Yaiquich, 2680 m, (15° 45' 44" N, 91° 30' 10" W). We visited this locality from 21 to 25 July 2005. The maximum temperatures during our stay averaged 17 °C, while minimum temperatures averaged 5 °C. It rained every day we were at this locality.
Carleton and Huckaby (1975) recorded Yaiquich as the type locality for Peromyscus mayensis. Their locality was 7 km NW of Santa Eulalia, 2950 m. From their description of the type locality, we thought we were there, especially since several local people said we were at Yaiquich. After speaking with Carleton and Huckaby (personal communication), we were close but probably about one-half to one kilometer to the southeast. However, from examination of Kodachrome slides by Huckaby, we had to be very close. His slide of the pasture/meadow looks very much like the one we trapped. Also, according to our GPS unit (Garmin III Plus), the elevation we recorded was much lower.
This disturbed cloud forest was composed of oaks, wild avocado, other broad-leaved trees, and pines. The forest floor was covered with mosses, leaf litter, and scattered small bracken fern. Fallen logs were numerous and usually covered with mosses. One area had an abundance of blackberry bushes. The meadow, with a running stream, was deep with grass.
Our trap-lines were set in the forest and in the meadow. We recorded 550 trap nights and 120 pitfall nights at this site.
While we were there, a crew from the Guatemalan Forestry Department was planting trees. Unfortunately, they were planting only pines (Pinus guatemalensis) to replace trees that were being removed. We observed workers taking from 2 to 4 cord stacks of firewood out of the forest each day. Most of this was hardwood. Along the road (RN-9) for several kilometers there were stacks of firewood waiting to be picked up by trucks and taken to the villages to be sold.
METHODS
Trapping effort
Small mammals were removal trapped using a combination of Victor rat, Museum Special, Sherman collapsible live, and pitfall (1 liter) traps. Each study area was sampled for a minimum of 3 nights (3-8 nights) using various arrays of traps and pitfalls. Trapping at each site ranged from 435 to 1513 trap nights (a trap night is one trap set per 24 hour period). Pitfall traps were used to optimize capture of shrews. Pitfall effort at sites ranged from 68 to 323 pitfall nights (a pitfall night is one pitfall trap set for 24 hrs). In general, traps were set at 5 m to 10 m intervals in straight lines. Traps were set to optimize capture of small mammals. Trapping procedures, data collection, and animal handling follow the guidelines of the American Society of Mammalogists (Gannon et al., 2007) as approved by San Jose State University, Institutional Animal Care and Use Committee, Protocol 851. All specimens collected are housed in the Museum of Natural History, University of San Carlos, Guatemala (USCG not catalogued but with field numbers of NOG), United States National Museum, Washington D. C. (USNM), or the Museum of Texas Tech University, Lubbock, Texas (TTU).
Statistical Analyses
Species diversity and similarity indices follow those given in Magurran (1988). Because our sampling was not standardized among sites, we use a simple count of species (species density) "S" as an index of diversity. To compare different sites, we use the Sǿrensen index of similarity "QS". Since trapping effort was different at each site, we use % of total capture as an index of relative abundance for each species.
Chi-Square was used to test for independence of male to female sex ratios when sample size from year and season for a species was 10 or greater.
Species Accounts
An annotated account of each species is given. These include, where data is available, comments on the ecology and natural history, as well as information as to species deposition. Information on reproductive activity was taken from personal catalogs and/ or data recorded on specimen tags.
RESULTS
Species diversity
A total of 445 individuals (28 shrews and 417 rodents) representing 13 species of small mammals was collected during the study (Table 1). With a total of 4670 trap nights and 726 pitfall nights, our catch was relatively low (about 8 percent). Two species (Reithrodontomys sumichrasti and Sorex veraepacis) were present at all sites. Two species were present in only one site each (R. mexicanus and Peromyscus mayensis).
Table 1. The total number of individuals per site with per cent relative abundance (in parentheses).
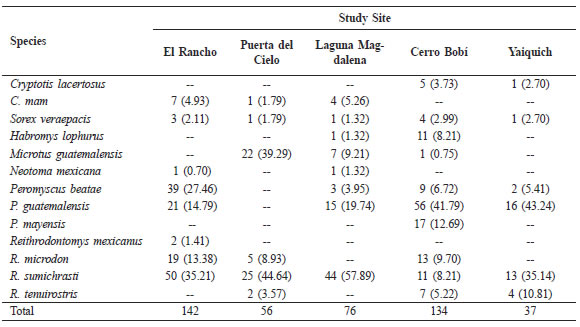
The number of species per site (S) ranged from 6 to 10 (Table 1 and 2). The greatest species density was found in the relatively pristine cloud forest at Cerro Bobí (S = 10), while the lowest was found in Puerta del Cielo and Yaiquich (both with S = 6).
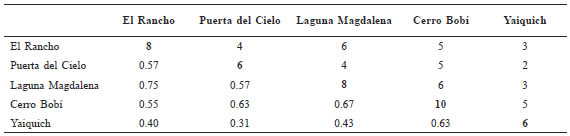
Table 2. Species density (S) and similarity (QS) among study sites. S (in bold) is on the diagonal; species in common to each of the two sites is above the diagonal; and, QS is below the diagonal.
Only two sites had sufficient trapping effort (a minimum of 150 trap nights each year) to warrant comparison of dry season sampling between years, Aldea El Rancho and Laguna Magdalena. Species diversity did not change greatly within the dry season between collecting years. Aldea El Rancho had only one species different between the two sampling periods. Neotoma mexicana was captured in December 2004, but not in 2008. Laguna Magdalena showed a bit more difference since two species (Habromys lophurus and N. mexicana) were taken in 2006 that were not taken in 2008. Also, S. veraepacis was taken in 2008 while it was not taken in 2006.
Only the Cerro Bobí site had sufficient trapping effort (a minimum of 150 trap nights each season) to warrant comparison of dry season (December/January 2005/2006) to wet season (July 2005) sampling. During each season the species density (S) was 9. During the dry season S. veraepacis was not recorded; however, it was taken during the wet season. On the other hand, M. guatemalensis was recorded during the dry season but not the wet season.
Species similarity
The similarity (QS) between sample sites (Table 2) ranged from 0.31 (between Puerta del Cielo and Yaiquich) and 0.75 (between Aldea El Rancho and Laguna Magdalena). The number of species in common (Table 2) ranged from two to six.
Relative abundance
Overall, R. sumichrasti was the most abundant small mammal taken during the study (Table 1). It was the dominant species in 3 (Aldea El Rancho, Puerta del Cielo, and Laguna Magdalena) out of the 5 study sites, and second in abundance at Yaiquich. Only at Cerro Bobí was R. sumichrasti at a relatively minor component of the community. At Cerro Bobí, the most abundant small mammal was P. guatemalensis. In all sites, two species accounted for over 50 percent of the catch; although, those species varied among sites.
Sex Ratios
Seventeen samples (year and season for a species) had n = 10. Of these, only one species, M. guatemalensis, from Puerta del Cielo, had a sex ratio that deviated from equality (n = 21, 15 male, 6 female; ?2 = 3.86, 1 df, p = 0.05).
Reproduction
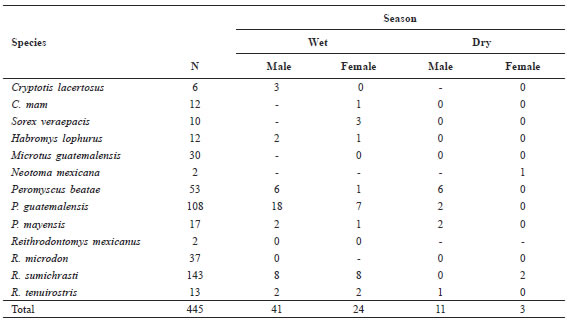
A summary of reproductive activity by season is given in Table 3 and recorded in the Species Accounts below. The data indicate that 79 (18%) of the 445 specimens collected showed evidence of reproduction. Of these reproductively active individuals, 82.3% show reproductive activity occurring during the wet season.
Table 3. Incidence, recorded as number of individuals showing evidence of reproductive activity, by season, in the Sierra de los Cuchumatanes small mammals. For males this is testes "scrotal" or "enlarged". For females it is evidence of lactation or presence of embryos. Wet season is July. Dry season is December and January.
Species Accounts
Species accounts are arranged by Order and Family, then alphabetically by genus and species following Wilson and Reeder (2005).
Order Soricomorpha Family Soricidae Cryptotis lacertosus Woodman, 2010
Comments.-This recently described species is known only from a restricted area in the Sierra de los Cuchumatanes. Woodman (2010) summarized the known ecology and natural history of C. lacertosus. He based his description on this series of specimens. Three males taken at Cerro Bobí in the wet season (July 2005) had enlarged testes. Two females (one in the wet season; one in the dry season, December 2005) did not show any sign of reproductive activity
Referred material.-(N = 6): Cerro Bobí (USNM 569420, 569431, 569442, 569443, 569503); Yiaquich (USNM 569368). Cryptotis mam Woodman, 2010
Comments.-Woodman (2010) described this species based upon specimens taken in 1895 by E. A. Goldman and E. W. Nelson. The type locality is only a few kilometers from our study site of Aldea El Rancho in essentially the same kind of habitat. Woodman (2010) mistakenly recorded one locality as Puerto al Cielo. The female specimen taken on 17 July 2008 at Puerta del Cielo was lactating.
Referred material.-(N = 12): Aldea El Rancho (USCG-NOG #'s 771, 804, 805; USNM 570256, 570257, 570313, 570314); Puerta del Cielo (USNM 570248); Laguna Magdalena (USNM 569554, 569555, 570337, 570340).
Sorex veraepacis veraepacis Alston, 1877
Comments.- Verapaz shrews were taken at all five study sites. While never very abundant in the Sierra de los Cuchumatanes, they were the second most abundant species in the Sierra de los Minas (Matson and McCarthy, 2005). Two of three adult female specimens from Cerro Bobí, taken during the wet season (28 and 30 July 2005) contained two embryos each, all in the right uterine horns. Crown rump lengths of the most proximal embryo from each female were 12 mm and 16 mm. The specimen we have from Yiaquich was lactating on 25 July 2005. None of the five specimens taken during the dry season (January 2005 and 2008) showed any sign of reproductive activity.
Referred material.-(N = 10): Aldea El Rancho (USCG-NOG 806; USNM 570293, 570294); Puerta del Cielo (USCG-NOG 823); Laguna Magdalena (USNM 570338); Cerro Bobí (USNM 569418, 569419, 569440, 569441); Yiaquich (USNM 569375).
Order Rodentia Family Cricetidae Habromys lophurus (Osgood, 1904)
Comments.- The type locality for this species is in Todos Santos Cuchumatán, Huehuetenango (Osgood 1904). We did not take them from the two sites near Todos Santos. We only found this species at two localities, Laguna Magdalena and Cerro Bobí. Two of four males taken during the wet season (July 2005) had scrotal testes (one was sub-adult), while one of three females contained 2 embryos, one in each uterine horn (crown rump length of the embryo in the left uterine horn measured 12 mm. None of the four specimens (two male, two female) taken in the dry season showed reproductive activity.
Referred material.-(N = 12): Laguna Magdalena (USNM 569556); Cerro Bobí (USNM 569392, 569423, 569433, 569434, 569436, 569448, 569449, 569527, 569545, 569546, 569548)
Microtus guatemalensis Merriam, 1898
Comments.-All but one of our specimens was taken in the dry season. None (including the one female taken in the wet season) showed evidence of reproductive activity. This is the only species that had an unequal sex ratio. The specimens from Puerta del Cielo, taken in the dry season, had a sex ratio of 2.5:1 in favor of males.
The type locality for M. guatemalensis, Todos Santos Cuchumatán (Merriam 1898), is close to where we sampled (Aldea El Rancho and Puerto del Cielo).
Referred material.-(N = 30): Puerta del Cielo (USCG-NOG 824-827, 848-855, 857, 863-869, 871; USNM 570249); Laguna Magdalena (USNM 569561-569566, 570339); Cerro Bobí (USNM 569504)
Neotoma mexicana Baird, 1855
Comments.-The taxonomic status of wood rats from south of the Isthmus of Tehuantepec is confused (Edwards and Bradley, 2002). Because of this and the assignment of all named taxa under the name N. mexicana by Hall (1981), we refer our specimens to this species.
A female taken in the dry season (January 2006) was lactating. Another female, also taken in the dry season (December 2004), showed no sign of reproductive activity.
Referred material.-(N = 2): Aldea El Rancho (USCG-NOG 784); Laguna Magdalena (USNM 569553).
Peromyscus beatae sacarensis Dickey, 1928
Comments.- Bradley et al. (2000) recognized P. b. sacarensis as the subspecies in the highlands of Guatemala.
Only five of 24 males taken in the dry seasons (December 2004, January 2005 and 2008) at Aldea El Rancho had scrotal testes. One female of 15 taken in the dry season was lactating. In the dry season (January 2006 and 2008) at Laguna Magdalena, one of two males had scrotal testes. The female did not evince any evidence of reproductive activity. In the wet season (July 2005) at Yiaquich, the two males had scrotal testes. At Cerro Bobí in the wet season (July 2005) all four males had scrotal testes, while none of the three females showed evidence of reproductive activity. Neither the male nor the female taken in the dry season (December 2005) at Cerro Bobí appeared to be reproductively active.
Referred material.-(N = 53): Aldea El Rancho (USCG-NOG 782, 783, 785, 786, 789, 796, 798, 799, 801, 802, 807, 820-822; USNM 570263- 570277, 570279, 570280, 570284, 570292, 570299, 570300, 570315-570318); Laguna Magdalena (USNM 569559, 569560, 570358); Yiaquich (USNM 569352, 569353); Cerro Bobí (USNM 569390, 569391, 569424, 569425, 569435, 569447, 569450, 569490, 569491).
Peromyscus guatemalensis Merriam, 1998
Comments.-None of the 21 specimens taken during the dry season (December 2004 and January 2008) at Aldea El Rancho showed sign of reproductive activity. Two males taken on 4 January (dry season, 2006) at Laguna Magdalena had scrotal testes; the female taken at the same time did not evince reproductive activity. One female of four taken at Laguna Magdalena in the dry season of 2008 had enlarged mammary glands. One of eight males from the same season had enlarged testes. All eight males taken at Yiaquich in July 2005 (wet season) had scrotal testes. Of eight females, at Yiaquich, two were lactating and one contained three embryos (two in the right uterine horn and one in the left). The crown rump length of the most proximal embryo in the right horn was 25 mm. At Cerro Bobí, in the wet season (July 2005), 10 of 14 males had scrotal testes, and four of 15 females had evidence of reproductive activity. Two were lactating. Two contained embryos: one had two embryos, one in each uterine horn (crown rump length 26 mm); the other had three embryos in the right uterine horn (crown rump length of the most proximal embryo was 12 mm).
Referred material.-(N = 108): Aldea El Rancho (USCG-NOG 779-781, 787, 788, 791-795, 800, 803; TTU 155365, 155366, 155376, 155378, 155379, 155381, 155382, 155384, 155385); Laguna Magda lena (USNM 569557, 569558, 569587; TTU 155364, 155367- 155369, 155370- 155375, 155377, 155380, 155583); Yiaquich (USNM 569351, 569354-569357, 569364, 569365, 569369, 569370, 569376-569382); Cerro Bobí (USNM 569393-569395, 569398, 569399, 569401, 569404-569417, 569427-569430, 569437-569439, 569451, 569452, 569492-569502, 569514-569518, 569531-569537, 569549-569552).
Peromyscus mayensis Carleton and Huckaby, 1975
Comments.- Important in this collection is the recording of Peromyscus mayensis, not formally reported since its original description by Carleton and Huckaby (1975). Although Reid (1997, 2009) reported upon some specimens, she did not give specific locality data. We did not capture P. mayensis in the cloud forest at Yaiquich near the type locality in July 2005. From the description of the habitat given by Carleton and Huckaby (1975), the forest had been considerably more disturbed by 2005. However, we did observe a heavy leaf and moss layer that seems to be a requirement for their presence (Carleton and Huckaby, 1975; Reid, 2009). They were quite abundant at Cerro Bobí, representing 9 percent of the wet season catch and 16 percent of the dry season catch.
In their original paper describing P. mayensis, Carleton and Huckaby (1975) recorded that all of the specimens taken in May were adults. Reid (1997) reported that individuals taken in December (dry season) were non-reproductive. Our data indicate that at least two of six individuals taken in July (wet season) were subadult, two males had scrotal testes, and one female contained two embryos both in the right uterine horn. The crown rump length of the most proximal embryo was 20 mm. In the dry season, two of four males had scrotal testes, while the seven females showed no evidence of reproductive activity.
Reid (1997, 2009) suggested that P. mayensis was insectivorous. An analysis of the stomach contents of six specimens taken in the dry season indicated only vegetation in the diet. There was no indication of any invertebrate material. Fleshy fruit was the major food item noted.
Referred material.-(N = 17): Cerro Bobí (USNM 569396, 569397, 569400, 569402, 569403, 569426, 569484-569486, 569505-569508, 569528-569530, 569547)
Reithrodontomys mexicanus howelli Goodwin, 1932
Comments.- Mexican harvest mice were taken only at Aldea El Rancho during this study. Anderson and Jones (1960) suggested this species is found at moderate elevations to 7000 ft (2272 m) in their study. Neither of the specimens showed evidence of reproductive activity.
Referred material.-(N = 2): Aldea El Rancho (USCG-NOG 808; USNM 570307)
Reithrodontomys microdon microdon Merriam, 1901
Comments.-Only one specimen of eight males and 11 females, a male taken on 8 January 2008 at Aldea El Rancho, showed evidence of reproductive activity. It had scrotal testes. None of the five specimens taken at Puerta del Cielo were reproductively active. None of the 13 specimens taken at Cerro Bobí was reproductively active.
Referred material.-(N = 37): Aldea El Rancho (USCG-NOG 816,819; USNM 570258-570261, 570283, 570295-570298, 570309, 570319-570325); Puerta del Cielo (USCG-NOG 829,832,836,844; USNM 570251); Cerro Bobí (USNM 569389, 569487, 569509-569511, 569523, 569524, 569538- 569543)
Reithrodontomys sumichrasti dorsalis Merriam, 1901
Comments.-Reproductive activity in this harvest mouse seems to be primarily during the wet season. Of 13 specimens taken during July 2005 (wet season) at Yiaquich, four males had scrotal testes, eight females were either lactating (3) or contained embryos (5). The pregnant specimens had from three to six embryos. At Cerro Bobí, in the wet season (July 2005), two of three males were scrotal, but none of the three females showed any sign of reproductive activity. In the dry season at Cerro Bobí (December 2005 and January 2006) none of the six individuals showed reproductive activity. Again, in the dry season (January 2006 and 2008) none of the 44 specimens from Laguna Magdalena showed sign of reproductive activity. At Puerta del Cielo none of the 21 specimens (9 female and 12 male) showed evidence of reproductive activity during the dry season (3-5 January 2005), while two males had scrotal testes on 17 July 2008. Two females on the same date did not have evidence of reproductive activity. In the dry seasons (31 December 2004 - 2 January 2005 and 6-8 January 2008) at Aldea El Rancho of 50 specimens (29 female and 21 male) only two females showed evidence of reproduction (both were lactating).
Referred material.-(N = 143): Aldea El Rancho (USCG-NOG 772-778, 790, 797, 809-815, 817, 818; USNM 570262, 570278, 570281, 570282, 570285- 570291, 570301-570306, 570308, 570310-570312, 570326-570336); Puerta del Cielo (USCG-NOG 828, 830, 831, 833-835, 837-843, 845-847, 856, 859-862; USNM 570252-570255); Laguna Magdalena (USNM 569567-569586, 570341-570357, 570359-570365); Yiaquich (USNM 569358-569363, 569366, 569371, 569383-569387); Cerro Bobí (USNM 569388, 569422, 569432, 569445, 569446, 569488, 569489, 569512, 569513, 569525, 569526).
Reithrodontomys tenuirostris tenuirostris Merriam 1901
Comments.-Neither of the two females (one in the wet season and one in the dry season) from Puerta del Cielo showed sign of reproduction. At Yiaquich in the wet season (July 2005) two males had scrotal testes and two females were lactating. None of the females taken at Cerro Bobí had evidence of reproductive activity, two in the wet season (July 2005) and four in the dry season (January 2006), while one male had scrotal testes in the dry season.
Referred material.-(N = 13): Puerta del Cielo (USCG-NOG 858; USNM 570250); Yiaquich (USNM 569367, 569372-569374); Cerro Bobí (USNM 569421, 569444, 569519-569522, 569544).
DISCUSSION
Recording 13 species from one mountain range represents a relatively high species density for highland habitats in Northern Nuclear Central America (Matson and McCarthy 2005; Woodman, 2010). However, these 13 are from five separate localities. The highest species density (S = 10) was found at only one site (Cerro Bobí). Rickart (1977) recorded 10 species of small mammals (three were shrews) from a cloud forest in Oaxaca, Mexico. Carleton and Huckaby (1975) also record 10 species for a site relatively near our site of Yaiquich, but with slightly different species composition. Matson and McCarthy (2005) recorded 10 and 12 species of small mammals from individual sites at San Lorenzo Marmól, Zacapa and Cerro Pinalón, El Progreso, respectively, in the Sierra de los Minas both with more extensive collecting effort. Woodman (2010) referring to E. W. Nelson and E. A. Goldman field notes and (Goldman 1951) recorded 11 species from the mountain ridge above Todos Santos. We only recorded eight species from this same mountain ridge. Species we did not record from Aldea El Rancho were H. lophurus, M. guatemalensis, and R. tenuirostris. However, the latter two species were taken over the same mountain ridge to the north of Todos Santos at Puerta del Cielo. One of us (NOG) had visited Aldea El Rancho several years earlier and took several specimens of M. guatemalensis. Why these three species did not occur during our visits is unknown.
Vásquez et al. (2000) considered cloud forest habitats to be among the richest in species density of any Neotropical or Nearctic habitats, and they recorded only seven species of small rodents. If we consider only small rodents, Cerro Bobí has eight species. Cerro Bobí also has the most mature of the cloud forests we studied. Again, Vásquez et al. (2000) demonstrated that mature cloud forest had a richer rodent fauna than disturbed habitats.
The capture of only six species of small mammals in a cloud forest habitat (Puerta del Cielo and Yaiquich) may be the lowest diversity recorded for the highlands of Guatemala. The low species density may be due to habitat disturbance in these areas or an artifact of our trapping regimen.
It would appear that the westernmost study sites (Aldea El Rancho, Puerta del Cielo, and Laguna Magdalena) differ in their small mammal communities compared to the easternmost sites (Cerro Bobí and Yaiquich). Especially important in these eastern sites are the presence of the Mayan mouse (P. mayensis), found only at Cerro Bobí in the present study, but also at Yaiquich by Carleton and Huckaby (1975) and the newly named Cryptotis lacertosus (Woodman, 2010). Inspection of Figure 1 and another map (1982. Defense Mapping Agency Aerospace Center, St. Louis Air Force Station, Missouri 63118. Map: 1:500,000; Tactical Pilotage Chart, Series TPC, Sheet K-25A, Edition 3) reveals that there are several deep valleys and high ridges between the western and eastern sites. Whether or not this diverse topography has influenced speciation in these groups cannot be determined at present. However, species diversity patterns can be affected by multiple speciation events (Rosenzweig, 1995; Brooks and McLennan, 2002). This is especially true in regions of high topographic relief, such as mountain ranges that may act as "...speciation machines" (Rosenzweig, 1995:89). Nuclear Central America (Savage, 1982; Campbell, 1999) and especially the Sierra de los Cuchumatanes (Anderson et al., 1973) represent an area with especially rugged mountain topography where the effects of allopatric speciation may have an effect on species diversity patterns such as community structure in small mammals.
The difference found during dry season samples during different years at Aldea El Rancho were not major. Only N. mexicana was missing from one year (2008). We attribute this to the difficulty in trapping wood rats at any locality. The differences recorded for Laguna Magdalena are probably more attributed to insufficient trapping effort.
The species density at Cerro Bobí is fairly consistent between wet and dry seasons. The differences in species can be attributed to chance, since both S. veraepacis and M. guatemalensis occurred in small numbers. However, Vásquez et al. (2000) demonstrated that there was temporal variation in community composition.
Species similarity among sites was rather low. The highest QS was only 0.75 (Aldea El Rancho and Laguna Magdalena). The number of species held in common between sites was quite low (Table 2). Six species were shared between Laguna Magdalena and El Rancho. Also, six (but not the same six) were held in common between La Laguna and Cerro Bobí. Six species were also held in common between Yaiquich and Cerro Bobí. We expected more equivalency between sites. Part of this lack of similarity may be due to the difference between the western versus eastern sites described above. We are at a loss to explain the difference between Aldea El Rancho and Puerta del Cielo. The major environmental differences are the steep slope, heavy winds, and relatively low vegetative heterogeneity at Puerta del Cielo. We can not at this time determine if the difference in small mammal community structure is due to these factors or to some other combination of factors.
Species relative abundances differed greatly between sites. Two species, S. veraepacis and R. sumichrasti were present at all sites. This is not unexpected for R. sumichrasti due to its relatively ubiquitous distribution (Sullivan et al., 2000) and its relatively high abundance at most sites (Table 1). On the other hand, the presence of S. veraepacis at all sites is more difficult to explain given its relatively low abundance. More interesting is the lack of P. guatemalensis from Puerta del Cielo. Because of its relatively high abundance in all other sites, we expected it to be at Puerta del Cielo as well. In fact, the genus Peromyscus appears to be entirely absent from this locality. Again it may be that the steep slope, heavy winds, and low vegetative heterogeneity of Puerta del Cielo have some effect on the species presence.
In general, each site had two species that were very abundant with the remaining species contributing less to the overall population structure. In general, a similar pattern of two dominant species was found in cloud forests in Western Mexico (Vásquez et al., 2000).
The three species of shrew and the wood rat were relatively low in abundance, usually representing less than 5 percent of the total. In the Sierra de las Minas (Matson and McCarthy, 2005) and at Todos Santos (in 1895/1896, according to Woodman 2010) shrews were much more abundant. In fact, at Todos Santos, according to Woodman (2010: 576) speaking about Cryptotis, "C. mam is the 2nd most abundant species represented."
Interestingly, only one species (M. guatemalensis), out of 17 possible samples, showed a sex ratio disparity. In the only other study in Guatemala to report sex ratios, Matson and McCarthy (2005) reported that for S. veraepacis, females outnumbered males three to one. The significance of sex ratio data is unknown at this time. In fact, since we tested 17 species, the uneven sex ratio for M. guatemalesis could be due to chance alone.
Reproduction in the Sierra de los Cuchumatanes seems to take place primarily during the wet season. In general, the wet season seems to be more conducive to reproduction than the dry season (Vásquez et al., 2000) for some species, although reproductive activity was recorded throughout the year in cloud forests of Western Mexico. Rickart (1977) found that, while P. melanocarpus was a seasonal breeder (wet season), there was a trend for aseasonal breeding in cloud forest deer mice. Our data indicate similar trends in reproductive activity, but with decidedly more reproductive activity during the wet season. Whether or not this is due to differences in the disturbed nature of some cloud forest habitats, as found by Vásquez, et al. (2000 and 2004), cannot be determined from our data.
The small mammal communities in the Sierra de los Cuchumatanes show considerable differences between sites. Whether this is due to real differences or due to chance because of different sampling regimens is unknown. We do know that in at least two sites, Todos Santos (near our Aldea El Rancho) and Yaiquich (type locality for P. mayensis), earlier studies show higher species densities than the present study (Woodman, 2010; Carleton and Huckaby, 1975).
ACKNOWLEDGMENTS
We are greatly in debt to the people of the local communities in the Sierra de los Cuchumatanes for allowing us to sample small mammals on their reserves and granting collecting permits. We thank the following institutions for partial funding of the field work: Department of Biological Sciences at San Jose State University (to JOM) and Northern Virginia Community College (WB and RPE). We thank the following personnel from the United States National Museum for help with species identifications: M. D. Carleton, A. Gardner, N. Woodman, and S. Peurach. We greatly appreciate the generous help in many facets of this project from S. Parsons, M. Eckerlin, and J. Bulmer. Neal Woodman provided much needed assistance during our January 2008 field work. Robert D. Bradley provided important suggestions and comments on the manuscript. We thank three anonymous reviewers for their suggestions and comments. Special thanks to Franklin Herrera at Consejo Nacional de Áreas Protegidas, Guatemala for providing collecting permits and other valuable assistance during the field work.
LITERATURE CITED
1. ANDERSON TH, B BURKART, RE CLEMONS, OH BOHNENBERGER, and DN BLOUNT. 1973. Geology of the Western Altos Cuchumatanes, Northwestern Guatemala. Geological Society of America Bulletin 84:805-826. [ Links ]
2. BRADLEY RD, I TIEMANN-BOEGE, CW KILPATRICK, and DJ SCHMIDLY. 2000. Taxonomic status of Peromyscus boylii sacarensis: inferences from DNA sequences of the mitochondrial cytochrome-b gene. Journal of Mammalogy, 81:875-884. [ Links ]
3. BROOKS DR and DA MCLENNAN. 2002. The nature of discovery. The University of Chicago Press, Chicago. [ Links ]
4. CAMPBELL JA. 1999. Distribution patterns of amphibians in Middle America. Pp. 111-210, en: Patterns of distribution of amphibians: a global perspective. (WE Duellman, ed.). Johns Hopkins University Press, Baltimore. [ Links ]
5. CARLETON MD and DG HUCKABY. 1975. A new species of Peromyscus from Guatemala. Journal of Mammalogy 56:444-451. [ Links ]
6. CASTRO-ARELLANO I. 2005. Ecological patterns of the small mammal communities at El Cielo Biosphere Reserve, Tamaulipas, Mexico. Tesis de doctorado inédita, Texas A&M University, College Station, Texas. [ Links ]
7. CASTRO-ARELLANO I. 2006. Ecological patterns of the small mammal communities at El Cielo Biosphere Reserve, Tamaulipas, Mexico. Mastozoología Neotropical 13:153-154. [ Links ]
8. CAYUELA L, DJ GOLICHE, and JM REY-B. 2006. The Extent, Distribution, and Fragmentation of Vanishing Montane Cloud Forest in the Highlands of Chiapas, Mexico. Biotropica 38:544-554. [ Links ]
9. DONNELLY TW. 1989. Geologic history of the Caribbean and Central America. Pp. 299-32, en: The Geology of North America: an overview (AW Bally and AR Palmer, eds.). Geological Society of America, Boulder, Colorado. [ Links ]
10. DONNELLY TW, GS HORNE, RC FINCH, and E LÓPEZ-RAMOS. 1990. Northern Central America; the Maya and Chortis blocks. Pp. 37-76 en: The Geology of North America: The Caribbean Region. (G Dengo and JE Case, eds.), Volume H. Geological Society of America, Boulder, Colorado. [ Links ]
11. DOUMENGE C, D GILMOUR, MR PÉREZ, and J BLOCKHUS. 1995. Tropical montane cloud forests: conservation status and management issues. Pp. 24-37 en: Tropical montane cloud forests. (LS Hamilton, JO Juvik, and F N Scatena, eds.). Springer-Verlag, New York. [ Links ]
12. EDWARDS CW and RD BRADLEY. 2002. Molecular systematics and historical phylobiogeography of the Neotoma mexicana species group. Journal of Mammalogy 83:20-30. [ Links ]
13. GANNON WL, RS SIKES, and THE ANIMAL CARE AND USE COMMITTEE OF THE AMERICAN SOCIETY OF MAMMALOGISTS. 2007. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. Journal of Mammalogy 88:809-823. [ Links ]
14. GOLDMAN EA. 1951. Biological Investigations in Mexico. Smithsonian Miscellaneous Collections, 115:1-476. [ Links ]
15. HALL ER. 1981. The Mammals of North America. 2nd ed. John Wiley & Sons, New York, 1:1-600 + 90. [ Links ]
16. HAMILTON, LS, JO JUVIK, and FN SCATENA (eds.). 1995. Tropical Montane Cloud Forests. Springer- Verlag, New York. [ Links ]
17. JANZEN DH (ed.). 1983. Costa Rican Natural History. The University of Chicago Press, Chicago. [ Links ]
18. MAGURRAN AE. 1988. Ecological diversity and its measurement. Princeton University Press, Princeton. [ Links ]
19. MATSON JO and TJ MCCARTHY. 2005. A new subspecies of verapaz shrew (Sorex veraepacis) from Guatemala. Pp. 63-70, en: Advances in the Biology of Shrews II (JF Merritt, S Churchfield, R Hutterer, and BI Sheftel, eds.). Special Publication of the International Society of Shrew Biologists. [ Links ]
20. MERRIAM CH. 1898. Descriptions of two new subgenera and three new species of Microtus from Mexico and Guatemala. Proceedings of the Biological Society of Washington 12:105-108. [ Links ]
21. MCCAIN CM. 2004. The mid-domain effect applied to elevational gradients: species richness of small mammals in Costa Rica. Journal of Biogeography 31:19-31. [ Links ]
22. MONTEAGUDO-SABATÉ D and L LEÓN-PANIAGUA. 2002. Estudio comparativo de los patrones de riqueza altitudinal de especies en mastofaunas de áreas montañosas Mexicanus. Revista Mexicana de Mastozoología 6: 60-82. [ Links ]
23. MYERS N, RA MITTERMEIER, CG MITTERMEIER, GA B DA FONSECA, and J KENT. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858. [ Links ]
24. ORDÓÑEZ-GARZA N, JO MATSON, RE STRAUSS, RD BRADLEY, and J SALAZAR-BRAVO. 2010. Patterns of phenotypic and genetic variation in three species of endemic Mesoamerican Peromyscus (Rodentia: Cricetidae). Journal of Mammalogy 91:848-859. [ Links ]
25. OSGOOD WH. 1904. Thirty new mice of the genus Peromyscus from Mexico and Guatemala. Proceedings of the Biological Society of Washington 17:55-77 [ Links ]
26. PARKER T. 2008. Trees of Guatemala. The Tree Press, Austin. [ Links ]
27. PATTERSON BD, PL MESERVE and BK LANG. 1989. Distribution and abundance of small mammals along an elevational transect in temperate rainforests of Chile. Journal of Mammalogy 70:67-78. [ Links ]
28. PATTERSON BD, DF STOTZ, S SOLARI, JW FITZPATRICK, and V PACHECO. 1998. Contrasting patterns of elevational zonation for birds and mammals in the Andes of Southeastern Peru. Journal of Biogeography 25:593-607 [ Links ]
29. REID FA. 1997. A field guide to the mammals of Central America and Southeast Mexico. Oxford University Press, New York. [ Links ]
30. REID FA. 2009. A field guide to the mammals of Central America and Southeast Mexico. 2nd ed. Oxford University Press, New York. [ Links ]
31. RICKART EA. 1977. Reproduction, growth and development in two species of cloud forest Peromyscus from southern Mexico. Occasional Papers, Museum of Natural History, University of Kansas 67:1-22. [ Links ]
32. ROSENZWEIG ML. 1995. Species diversity in space and time. Cambridge University Press, Cambridge. [ Links ]
33. SÁNCHEZ-CORDERO V. 2001. Elevation gradients of diversity for rodents and bats in Oaxaca, Mexico. Global Ecology & Biogeography 10:63-76 [ Links ]
34. SAVAGE JM. 1982. The enigma of the Central American herpetofauna: dispersals or vicariance? Annals of the Missouri Botanical Garden 69:464-597. [ Links ]
35. SULLIVAN JK, E ARELLANO, and D ROGERS. 2000. Comparative phylogeography of Mesoamerican highland rodents: concerted versus independent response to past climatic fluctuations. The American Naturalist, 155:755-768. [ Links ]
36. TIMM RM and RK LAVAL. 2000. Mammals. Pp. 223-244, en: Ecology and Conservation of a Tropical Cloud Forest (NM Nadkarni and NT Wheelwright, eds.). Oxford University Press, New York. [ Links ]
37. TIMM RM, DE WILSON, BL CLAUSON, RK LAVAL, and CS VAUGHAN. 1989. Mammals of the La Selva-Braulio Carrillo Complex, Costa Rica. North American Fauna 75:1-162. [ Links ]
38. VARGAS-CONTRERAS JA and A HERNÁNDEZ-HUERTA. 2001. Distribución altitudinal de la mastofauna en la Reserva de la Biosfera "El Cielo", Tamaulipas, México. Acta Zoológica Mexicana 82:89-109. [ Links ]
39. VÁZQUEZ LB, GN CAMERON, and RA MEDELLÍN. 2004. Characteristics of diet of Peromyscus aztecus and Reithrodontomys fulvescens in montane Western Mexico. Journal of Mammalogy 85:196-205. [ Links ]
40. VÁZQUEZ LB, RA MEDELLÍN, and GN CAMERON. 2000. Population and community ecology of small rodents in montane forest of Western Mexico. Journal of Mammalogy 81:77-85. [ Links ]
41. WEGE DC and AJ LONG. 1995. Key areas for threatened birds in the Neotropics. BirdLife Conservation Series, No. 5. BirdLife International, Cambridge. [ Links ]
42. WILSON DE and DM REEDER (eds.). 2005. Mammal species of the world: a taxonomic and geographic reference. 3rd ed. Johns Hopkins University Press, Baltimore. [ Links ]
43. WOODMAN N. 2010. Two new species of shrews (Soricidae) from the western highlands of Guatemala. Journal of Mammalogy 91:566-579. [ Links ]

