










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383
Mastozool. neotrop. vol.19 no.1 Mendoza jun. 2012
ARTÍCULOS Y NOTAS
Rapto de una cría por hembra parturienta y cuidado compartido en el lobo fino Sudamericano Arctocephalus australis
Mayda Rivas y Valentina Franco-Trecu
Proyecto Pinnípedos, Sección Etología, Facultad de Ciencias, Universidad de la República, Iguá 4225, Montevideo 11400, Uruguay [correspondencia: Valentina Franco-Trecu <pinnipedosuy@gmail.com>].
Recibido 2 junio 2011.
Aceptado 5 octubre 2011.
Editor asociado: L Guichón
RESUMEN: Esta nota reporta el rapto y adopción de una cría por parte de una hembra de lobo fino sudamericano Arctocephalus australis durante el parto, con el posterior rechazo a su propio cachorro. La cría fue raptada cuando se encontraba junto a su madre, quien intentó recuperarla sin éxito. En el mes siguiente se observó a la cría amamantando de su madre y de la madre adoptiva. Este episodio aporta información sobre la alta sensibilidad de las hembras de esta especie durante el parto y el corto período de selectividad maternal que puede establecerse incluso antes del parto.
ABSTRACT: Kidnapping of a pup by a parturient female and shared care in the South American fur seal, Arctocephalus australis. This communication reports a case of kidnapping and adoption of a pup by a South American fur seal female (Arctocephalus australis) while she was in labor, rejecting her own pup when it was born. The kidnapped pup was taken from its mother, who tried unsuccessfully to retrieve it. In the following month, the pup was observed nursing both from its mother and the adoptive female. This episode provides new insights into the high maternal sensitivity during parturition and the short period of maternal selectivity that can be established even before birth in this species.
Palabras clave: Cuidado alomaternal; Otáridos; Período de selectividad; Sensibilidad maternal durante el parto.
Key words: Allomaternal care; Maternal sensitivity during parturition; Otariids; Selectivity period.
En los mamíferos, los mecanismos involucrados que aseguran a la hembra una correcta inversión en su descendencia difieren entre especies altriciales y precoces (Trillmich, 2010). En general, las especies con crías altriciales no muestran selectividad maternal, mientras que en las precoces existe un período crítico y sensible tras el parto, durante el cual se establece un vínculo selectivo con el neonato, proceso conocido como "imprinting materno" (e. g., Klopfer et al., 1964). Si bien el período sensible (fuerte atracción hacia características de los neonatos) y el crítico o de selectividad (fijación de determinados rasgos de la cría en la memoria de la madre) generalmente se dan al mismo tiempo, son procesos diferentes (e. g., Poindron et al., 1993).
En los otáridos (lobos y leones marinos) el cuidado de las crías es exclusivo de las hembras. Durante la lactancia éstas alternan períodos en tierra, donde cuidan y amamantan a sus crías, con viajes de alimentación en el mar, períodos que las crías permanecen en ayuno en la colonia (Boness y Bowen, 1996). El reconocimiento madre-cría se desarrolla tempranamente, durante los primeros días postparto, y permite que ambas se reencuentren luego de cada viaje de alimentación de la hembra (Charrier et al., 2001). En el lobo marino subantártico (Arctocephalus tropicalis) las hembras son capaces de reconocer a sus crías pocos minutos después de su nacimiento a través de señales acústicas y olfativas (Charrier et al., 2003). Sin embargo, experimentos de playback en A. tropicalis muestran que las crías no son selectivas frente a vocalizaciones de las hembras horas después del nacimiento, sino que tardan de dos a cinco días en responder exclusivamente a sus madres (Charrier et al., 2001). Si bien las hembras utilizan pistas espaciales, acústicas y olfativas para encontrar y reconocer a su cría (Insley et al., 2003), se ha sugerido que una sola modalidad sensorial sería efectiva de forma independiente (Trimble e Insley, 2010). Se han reportado casos en que sólo las señales olfativas fueron suficientes para una reunión madre-cría exitosa en A. australis (Phillips, 2003), A. gazella (Dobson y Jouventin, 2003) y Otaria flavescens (Fernández-Juricic et al., 1999; Trimble e Insley, 2010).
En los otáridos el reconocimiento madre-cría es importante debido a que son especies gregarias que viven en colonias numerosas y las hembras deben identificar a su cachorro entre otros cientos para evitar realizar el gasto energético de cuidar crías ajenas, maximizando su éxito. Sin embargo, la ocurrencia de adopción y aloamamantamiento (i.e., amamantamiento de crías no filiales) en este grupo sugiere errores en el reconocimiento entre madres y crías, o ausencia de dicho proceso (Insley et al., 2003). Estos comportamientos son raros entre los otáridos pero más comunes en los fócidos (focas verdaderas), ya que en este grupo la lactancia tiene corta duración y las hembras permanecen junto a sus crías desde el nacimiento hasta el destete, por lo que no sería necesario un reconocimiento madre-cría tan estricto (Insley et al., 2003).
Esta comunicación reporta el rapto y adopción de una cría por parte de una hembra de lobo marino sudamericano A. australis en trabajo de parto, con el posterior rechazo de su propio cachorro. Conjuntamente se informa un caso de crianza compartida entre dos hembras.
Se realizaron observaciones de comportamiento sobre una colonia de lobo fino sudamericano en el marco de una investigación desarrollada en Isla de Lobos, Uruguay (35º 01' S, 54º 52' O). El 3 de diciembre de 2008 a las 19:33 hs, durante el registro de un parto caudal, la hembra focal (H1) tomó una cría ajena que rápidamente fue recuperada por su madre. Posteriormente la H1 tomó y retuvo otra cría ajena (C2) que estaba junto a su madre (H2), iniciándose en ese momento un muestreo focal hasta las 21:00 hs sobre los tres individuos (H1, C2 y H2; Fig. 1). Cuando la H1 tomó la C2, la H2 intentó recuperarla y ambas hembras trataron de retener la cría simultáneamente, hasta que se interpuso un macho. La H2 intentó acercarse a su cría para recuperarla pero la H1 la retuvo con su boca interponiendo su cuerpo. La C2 vocalizó numerosas veces en respuesta a los llamados de su madre (H2). Mientras la H1 retenía a la C2, la olfateó pero no emitió vocalizaciones. Transcurridos 11 minutos del inicio del muestreo focal, la H1 expulsó completamente a su cría (C1), sin interrumpir su contacto cercano con la C2, dándose por finalizado el parto. En los minutos siguientes, la H1 olfateó a su cría recién nacida (C1) en tres ocasiones, siendo el único comportamiento que la H1 realizó hacia la C1, para luego ignorarla completamente. La C1 vocalizó varias veces sin buscar de forma activa el contacto corporal con su madre. Esto es común en la mayoría de las crías recién nacidas ya que son estimuladas por sus madres a través de vocalizaciones y contactos de su hocico (VFT datos no publicados). Durante el registro focal la H2 se mantuvo cerca de su cría emitiendo vocalizaciones que fueron respondidas por la C2, indicando que la C2 ya había grabado la firma vocal de su madre (H2), sin embargo permaneció junto a la H1 y se alimentó de ella.
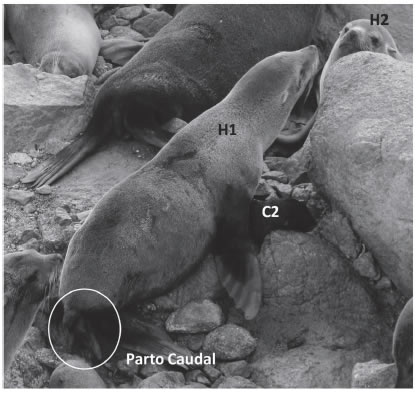
Fig. 1: La foto muestra a la hembra focal (H1) reteniendo a la cría ajena (C2), mientras está en trabajo de parto. A su lado se encuentra la madre de la cría raptada (H2). Isla de Lobos, Uruguay.
Inmediatamente después de finalizar la observación focal, ambas hembras y crías fueron marcadas a distancia con una pequeña marca de pintura de diferentes colores. Esta metodología es ampliamente utilizada en pinnípedos (e.g., Trimble e Insley, 2010) sin registrarse ningún efecto en el reconocimiento madre-cría. Posteriormente se realizó un seguimiento de dichos individuos por 30 días, considerando el tres de diciembre como el día uno. En el día dos la C1 fue observada muerta, mientras la H1 continuaba en contacto con la C2. Durante los siguientes 30 días ambas hembras (H1 y H2) realizaron viajes de alimentación y durante sus estadías en tierra ambas fueron observadas amamantando únicamente a la C2. El día cinco (7 de diciembre) la C2 fue capturada, sexada como hembra y marcada con caravanas Allflex N° 12. Su peso inicial fue de 4800 g y su peso final (24 de diciembre) fue de 5100 g. La tasa de crecimiento (tc) se calculó como dP/dt, donde dP es la diferencia entre el peso final y el inicial, y dt la diferencia en días entre ambos registros. La tc de la C2 (17.65 g/día) se ubicó dentro del intervalo de confianza de las tc para las crías durante dicha temporada (X=38.64 g/día, IC 95%: 14.61 - 63.69, n = 42, VFT datos no publicados). La tc se calculó para todas las crías de la colonia dado que en esta especie no se han reportado diferencias significativas en las tasas de crecimiento entre sexos (Franco-Trecu, 2010). A pesar de que la C2 se alimentó de ambas hembras, su tc fue relativamente baja en comparación con el promedio de la población.
Si bien durante el muestreo focal se observó que la H1 olfateó en varias ocasiones a la C2 (grabando su señal olfativa) no emitió vocalizaciones, contrariamente a lo que se observa luego o incluso durante un parto en esta especie (MR y VFT datos no publicados). Si bien estas observaciones sugieren que no es necesario el uso de ambos tipos de señales para reencuentros madre-cría exitosos, es posible que la H1 haya vocalizado luego de que terminó el muestreo focal (e. g. durante la noche) y que la C2 haya grabado su firma. De otro modo sería difícil el reencuentro entre ambas luego de los viajes de alimentación de la H1, considerando que las crías tienen un rol importante en el éxito de los reencuentros madre-cría en esta especie (Phillips, 2003). En estas condiciones, la cría habría grabado las firmas vocales de ambas hembras, indicando que podrían ser capaces de memorizar más de una firma vocal.
El registro de este evento, además de comunicar un episodio poco frecuente en la especie, sugiere que si el período de alta sensibilidad de las hembras comienza durante el parto, las hembras pueden sentirse atraídas por crías ajenas, debido a que su propio cachorro aún no está presente. El hecho de que este período se inicie antes de que finalice el parto podría tener consecuencias negativas para las hembras, ya que se encuentran en grupos donde hay cientos de crías. En este contexto, un período de sensibilidad establecido previo a la finalización del parto podría no ser adaptativo para la especie, por lo que interpretamos este registro como un caso raro. Asimismo, dado que la C2 no era recién nacida, la sensibilidad maternal durante el parto sería amplia, ya que la hembra fue atraída por estímulos no necesariamente propios de un neonato. Por otro lado, la selección natural operaría sobre un período crítico o de selectividad maternal de corta duración, ya que las hembras se encuentran rodeadas de crías ajenas. Este caso sugiere que el período de selectividad podría darse incluso antes del parto si la hembra es atraída por otro cachorro, viéndose reflejado en el rechazo de la H1 hacia su propia cría, ya que durante su nacimiento la hembra grabó las señales de la C2, ignorando posteriormente a la C1. Finalmente, si bien este registro sugiere que los casos de adopción en otáridos podrían surgir a partir de errores en el establecimiento del reconocimiento madre-cría, también cuestiona la selectividad de los cachorros de A. australis a la hora de alimentarse, aportando información para explicar las altas frecuencias de aloamamantamiento en esta colonia (Franco- Trecu et al., 2010) y en otras especies de lobos finos (Gemmell, 2003; de Bruyn et al., 2010).
Agradecimientos.
Agradecemos a Rosina Frau, "Bocha" Olivera, "Mulato" Veiga y "Palito" Casella por su asistencia en el campo. A la DI.NA.R.A. por el apoyo logístico y por los permisos (490/08) para llevar a cabo la investigación en Isla de Lobos, Uruguay. A Annabel Ferreira por sus valiosos aportes al manuscrito y a Micaela Trimble por su colaboración con el inglés. Finalmente, queremos agradecer a Florencia Grandi y un revisor anónimo por los valiosos aportes que realizaron.
LITERATURA CITADA
1. BONESS DJ y WD BOWEN. 1996. The evolution of maternal care in pinnipeds. Bioscience 46:645-654. [ Links ]
2. CHARRIER I, N MATHEVON y P JOUVENTIN. 2001. Mother's voice recognition by seal pups - Newborns need to learn their mother's call before she can take off on a fishing trip. Nature 412:873. [ Links ]
3. CHARRIER I, N MATHEVON y P JOUVENTIN. 2003. Fur Seal mothers memorize subsequent versions of developing pups' calls: adaptation to long-term recognition or evolutionary by-product? Biological Journal of the Linnean Society 80:305-312. [ Links ]
4. DE BRUYN PJN, EZ CAMERON, CA TOSH, WC OOSTHUIZEN, RR REISINGER, NT MUFANADZO, MV PHALANNDWA, M POSTMA, M WEGE, DS VAN DER MERWE y MN BESTER 2010. Prevalence of allosuckling behaviour in Subantarctic fur seal pups. Mammalian Biology 75:555-560. [ Links ]
5. DOBSON FS y P JOUVENTIN. 2003. How mothers find their pups in a colony of Antarctic fur seals. Behavioural Processes 61:77-85. [ Links ]
6. FERNANDEZ-JURICIC E, C CAMPAGNA, V ENRIQUEZ y CL ORTIZ. 1999. Vocal communication and individual variation in breeding South American sea lions. Behaviour 136:495-517. [ Links ]
7. FRANCO-TRECU V. 2010. Éxito de crianza y hábitos alimenticios en hembras de Arctocephalus australis y su relación trófica con hembras de Otaria flavescens. Tesis de Maestría inédita, PEDECIBA Biología, Universidad de la República, Montevideo, Uruguay. [ Links ]
8. FRANCO-TRECU V, B TASSINO y A SOUTULLO. 2010. Allo-suckling in the South American fur seal (Arctocephalus australis) in Isla de Lobos, Uruguay: cost or benefit of living in a group? Ethology Ecology & Evolution 22(2):143-150. [ Links ]
9. GEMMELL NJ. 2003. Kin selection may influence fostering behaviour in Antarctic fur seals (Arctocephalus gazella). Proceedings of the Royal Society of London. Series B. Biological Sciences 270:2033-2037. [ Links ]
10. INSLEY SJ, AV PHILLIPS y I CHARRIER. 2003. A review of social recognition in pinnipeds. Aquatic Mammals 29(2):181-201. [ Links ]
11. KLOPFER PH, DK ADAMS y MS KLOPFER. 1964. Maternal'imprinting' in goats. Proceedings of the National Academy of Sciences of the United States of America 52:911-914. [ Links ]
12. PHILLIPS AV. 2003. Behavioral cues used in reunions between mother and pup South American fur seals. Journal of Mammalogy 84(2):524-535. [ Links ]
13. POINDRON P, R NOWAK, F LÉVY, RH PORTER y B SCHAAL. 1993. Development of exclusive mother-young bonding in sheep and goats. Oxford reviews of Reproductive Biology 15:311-364. [ Links ]
14. TRILLMICH F. 2010. Parental care: adjustments to conflict and cooperation. Pp. 267-298, en: Animal Behaviour: Evolution and Mechanisms (P Kappeler, ed.). Springer Berlin Heidelberg. [ Links ]
15. TRIMBLE M y SJ INSLEY. 2010. Mother-offspring reunion in the South American sea lion Otaria flavescens at Isla de Lobos (Uruguay): use of spatial, acoustic and olfactory cues. Ethology Ecology & Evolution 22(3):233-246. [ Links ]

