










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383
Mastozool. neotrop. vol.19 no.2 Mendoza jul./dic. 2012
ARTÍCULOS Y NOTAS
Notice on the ocurrence of supernumerary teeth in the two-toad sloths Choloepus didactylus and C. hoffmanni
Robert K. McAfee1 and Virginia L. Naples2
1 Ohio Northern University, Department of Biological & Allied Health Sciences, 525 S. Main St. Ada, OH 45810, USA [Correspondence: <rkmcafee@gmaiLcom>].
2 Northern Illinois University, Department of Biological Sciences, Montgomery Hall, DeKalb, IL 60115 USA.
Recibido 13 junio 2012.
Aceptado 14 agosto 2012.
Editor asociado: A Abba
ABSTRACT: Supernumerary teeth occur in nearly all modern mammalian orders, although no previous cases have been published for extant pilosans. We report here three specimens of the sloth genus Choloepus, two C. hoffmanni and one C. didactylus, exhibiting supernumerary teeth. All duplicated teeth are maxillary, and all but one on the right side. One C. hoffmanni bears an additional left tooth, suggesting duplication is neither species nor toothrow-side specific. Sloth teeth erupt as simple rounded cones and the different "cusp" patterns characteristic of each extant and extinct genus arise from wear and the differential spacing of upper versus lower dentitions.
RESUMEN: Noticias sobre la ocurrencia de dientes supernumerarios en perezosos de dos dedos, Choloepus didactylus y C. hoffmanni. Si bien casi todos los órdenes de mamíferos modernos presentan dientes supernumerarios, no se habían publicado casos entre pilosos vivientes. En este estudio se documentan dientes supernumerarios en tres individuos del género de perezosos Choloepus: dos C. hoffmani y un C. didactylus. Todos los dientes duplicados se encuentran en el maxilar derecho. Uno de los dos C. hoffmani presenta, además, un diente duplicado del lado izquierdo, lo que sugiere que la duplicación no es específica a nivel de taxón o lateralidad. Los dientes de los perezosos erupcionan como conos simples redondeados y, por lo tanto, la variedad de patrones cuspidales característicos de cada género se produce por el desgaste y el desplazamiento en el espacio de las denticiones superior e inferior.
Key words. Choloepus; Dental anomaly; Megalonychidae; Supernumerary teeth; Xenarthra.
Palabras clave. Anomalía dentaria; Choloepus; Dientes supernumerarios; Xenarthra; Megalonychidae.
Members of the Xenarthra (Order Cingulata: armadillos, glyptodonts, pampatheres; Order Pilosa: anteaters, sloths), have long been regarded as noteworthy among mammalian species for the uniqueness of their dentition, both in structure and dental formula. The tooth- bearing members of this group (anteat-ers are edentulous) stand out from all other mammals, except the aardvark (Orycteropus afer), by lacking an enamel covering on the teeth (Shoshani et al., 1988). Instead, the teeth are composed of two layers of dentine, a soft inner region of orthovasodentine and a harder outer layer of orthodentine, both of which are surrounded by a covering of cementum (Ferigolo, 1985; Kalthoff, 2011). Like those of the aardvark, xenarthran teeth are ever-growing, high crowned but differ in dental formula and dentine structure (Schmidt and Keil, 1971). The dental formula in aardvarks is 0.0.2-3.3/0.0.2.3 (Shoshani et al., 1988), whereas no dental formula exists for xenarthrans because their teeth do not correspond with or have a known ho-mology to the typical dental classification used for mammals, i.e., incisors, canines, premolars and molars. The dentine of aardvarks is also unique to that taxon and is unlike that of any other mammals or vertebrates, whereas that of xenarthrans is similar to the orthodentine of other mammals but with some modifications (Schmidt and Keil, 1971). Among sloths, the teeth are often referred to as being molariform or caniniform, based on their shapes and locations along the toothrow.
The dental formula also varies between the cingulates and tooth-bearing pilosans. Cingulate taxa tend to have a high number of teeth in the four dental quadrants with the extant forms ranging from as low as 7-8/8 to as high as 18/19 (Wetzel, 1985a, 1985b). The sloths, on the other hand, are united by a drastic reduction in the number of teeth, with most having a generalized formula of either 4/4, 4/3, or 5/4 (Pujos et al., 2011). Reduction in tooth number is a common character among sloths, with the two genera of extant tree sloths (the three-toed sloth, Bradypus, and the two-toed sloth, Choloepus) possessing a 5/4 dental formula. Tooth number and lack of enamel are the only commonalities between the two sloth taxa, as the two-toed sloths display the typical megalonychid condition of a triangular anterior tooth (caniniform) separated from the posterior molariforms by a diastema; a feature lacking in three-toed sloths. Instead, the anterior teeth in three-toed sloths are relatively smaller than the anterior caniniforms of two-toed sloths, and are peg or chisel-shaped (Naples, 1982, 1995).
Molecular data, as well as historical anatomical studies, have shown the close relationship between the cingulates and pilosans, with a divergence of the two groups at around 65 million years ago (Delsuc et al., 2002, 2004), indicating that sloths came from an ancestral stock with a dental formula including a larger number of teeth. Changes in dental formulae have been used to analyze the relationships among numerous mammal groups, mostly on the basis of the presence of extra or supernumerary teeth (Drehmer et al., 2004; Loch et al., 2010). The occurrence of supernumerary teeth has been well documented in extant members of all mammalian orders (at least those that possess teeth; e.g. Suborder Vermilingua is edentulous) (Wolsan, 1984) but this particular dental anomaly has not been observed in either of the two extant sloth genera until now. Here we present the first examples of supernumerary teeth in the dentition of members of the extant tree sloth genus Choloepus.
Specimens of Choloepus spp. (77 total) were examined from the collections of the Division of Mammals of the Field Museum of Natural History (FMNH) and the Kansas Museum of Natural History (KU). The specimens were all wild-caught and excluded zoo specimens. In total, only three specimens exhibited the supernumerary condition for a 3.89% occurrence rate (2/54 from FMNH: 35738 and 41503; 1/23 from KU: 115214). Both species, Choloepus didactylus (Linnaeus's two-toed sloth) and Choloepus hoffmanni (Hoffmann's two-toed sloth) exhibit the condition, and all supernumerary teeth are located in the right maxillary toothrow. The exception to this is KU 115214 (C. hoffmanni from Nicaragua) which has supernumerary teeth in both the left and right maxillary toothrows. Supernumerary teeth are designated with a 'd' prior to the abbreviation for the molariform position; i.e., dM1 for a duplicate/supernumerary first upper molariform.
The lone supernumerary tooth observed in C. didactylus (FMNH 41503: Ecuador) is an adult that is also missing one tooth in the upper right series (Fig. 1 A-B), but the remaining teeth and the empty alveolus demonstrate that the duplication is of either M1 or M2. Regardless of the exact tooth position, the posterior portion of the toothrow has been crowded and pushed slightly posteriorly. The right dentition of the mandibular toothrow is likewise affected with a gap occurring between m1 and m2 (Fig. 1 C), as a result of the posterior displacement of the latter. The m2 also lacks the typical pointed appearance of Choloepus molariform wear facets (Naples, 1982, 1995); instead resembling more of a roughened plateau with a slight slant along the most anterior and posterior edges. The wear facet morphology of m2 matches that of M3, which occludes directly with the tooth (Fig. 1 A), as opposed to being offset anteriorly by a half tooth length as in the typical sloth tooth spacing arrangement. M3 also bears a cusp anterolabially instead of posteriorly.

Fig. 1. Skull and mandible of Choloepus didac-tylus (FMNH 41503) in right lateral (A and C) and ventral (B) views. The supernumerary tooth position is difficult to differentiate between the second or third maxillary molariform on the right side (B). Scales = 10 mm.
In the Peruvian specimen of C. hoffmanni (FMNH 35738), the extra tooth appears to be a duplication of M1, occurring just anterior to the actual M1, based upon the orientation of the tooth (Fig. 2 A-B). In Choloepus, as in a number of other sloths genera, M1 is recurved posteriorly so that the wear facet also faces obliquely and posteriorly to contact the anterior edge of ml. The dMl in this specimen is not recurved, and the oblique wear facet faces anteriorly, where it comes into occlusion with the posterior edge of c1 (Fig. 2 A), giving the lower caniniform two wear facets (Fig. 2 C). The remainder of the dentition exhibits the normal wear patterns associated with Choloepus, with the exception of the lower left molariforms where the cusps are greater on the lingual side instead of the labial.

Fig. 2. Skull and mandible of Choloepus hoffmanni (FMNH 35738) in occlusion in right lateral view (A), with dM1 visible in contact with the posterior aspect of C1. The ventral view of the skull (B) and the left dorsolateral view of the mandible (C) are positioned to better illustrate the dental wear features of the maxillary and mandibular teeth. Scales = 10 mm.
KU 115214 is a juvenile C. hoffmanni, based on the simplicity of cranial sutures and the lack of their closure/fusion (Figs. 3and 4 A). This specimen also stands out by having a duplicate tooth in both upper toothrows; an extra right M2 and a left M1 (Fig. 4 A). On the right side, dM2 abuts posteriorly with the anterior edge of the proper M2, such that both together have a pointed appearance (Fig. 3 A) due to the oppositely oriented wear facets; dM2 nearly touches the posterior edge of M1, faces anteriorly, and occludes with the posterior m1 facet; M2 proper faces posteriorly and occludes with the anterior facet of m2. The right mandibular dentition has the typical Choloepus morphology (Fig. 4 B-C). The supernumerary left dM1 occurs anterior to M1 proper and is conical in shape with only the slightest portion of the tip showing any sort of wear (Fig. 3 B). All other teeth in the series, maxillary and mandibular, appear unchanged in morphology (Fig. 4 A-C).

Fig. 3. Occluded skull and mandible of Choloepus hoffmanni (KU 115213) in right (A) and left (B) lateral views, illustrating the supernumerary teeth in both toothrows. Scales = 10 mm.
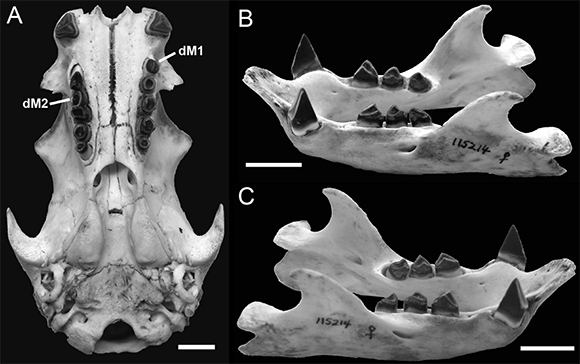
Fig. 4. Ventral view of the skull (A) and lateral views of the mandible (B and C) of C. hoffmanni (KU 115214), to further demonstrate the dental morphologies and wear facets of all the teeth. Scales = 10 mm.
Various dental anomalies are common among mammals (Miles and Grigson, 1990; Wolsan, 1984) and with the above specimens, a supernumerary tooth condition is now demonstrated as occurring in all tooth-bearing orders. The three specimens of Choloepus exhibit additional teeth in their maxillary dentition, without restriction to a specific species level of occurrence. Interestingly, the supernumerary condition in sloths appears to be an artifact of the Family Megalonychidae, as one occurrence has been noted in the extinct genus Megalonyx, in which the additional tooth was also restricted to the maxillary dentition (McDonald, 1978; Miles and Grigson, 1990). Of the numerous other extinct sloth taxa, no other genus has demonstrated a supernumerary condition. The only tooth number modification in the extinct forms has been the loss of a tooth (adontia).
This adontia has so far been confined or noted only in some members of the subfamily Mylodontinae; specifically in the Plio-Pleistocene genera of Mylodon from South America and Paramylodon from North America (McAfee, 2007, 2009). It remains to be seen what other dental anomalies are present within the group and what effects they would have had upon the feeding behaviors in those individuals.
To conclude, herein is presented the irst documented cases of supernumerary teeth in the extant members of the mammalian Order Pilosa. The supernumerary condition currently occurs only within the two species of two-toed sloths, C. didactlyus and C. hoffmanni. Sloth dental morphology arises from the unique contact wear by the occluding teeth and the half-tooth length offset of the spacing of the upper and lower dentitions. Additional teeth disrupt these relationships and may alter the orientation of the typical anterolingual masticatory power stroke in these animals. It remains to be seen what effect the changes in tooth position have upon the dental morphologies and subsequent feeding mechanics, and additional research is required to determine whether this and other dental anomalies also occur in three-toed sloths.
Acknowledgements. The authors are thankful to W. Stanley of the Field Museum and R. Timm of the University of Kansas for access to the mammalogy collections and their assistance. Thanks also to A. Calvo and A. Cordoba for translational assistance. This manuscript benefitted from the comments and suggestions by reviewers J. Green and F. Galliari.
LITERATURE CITED
1. DELSUC F, M SCALLY, O MADSEN, MJ STANHOPE, WW DE JONG, FM CATZEFLIS, M SPRINGER and EJP DOUZERY. 2002. Molecular phylogeny of living xenarthrans and the impact of character and taxon sampling on the placental tree rooting. Molecular Biology Evolution 19:1656-1671. [ Links ]
2. DELSUC F, SF VIZCAÍNO, and EJP DOUZERY 2004. Influence of Tertiary paleoenvironmental changes on the diversification of South American mammals: a relaxed molecular clock study within xenarthrans. BMC Evolutionary Biology 4:1-11.
3. DREHMER CJ, ME FABIÁN, and JO MENEGHETI. 2004. Dental anomalies in the Atlantic population of South American sea lion, Otaria bryonia (Pinnipedia, Otariidae): Evolutionary implications and ecological approach. Latin American Journal of Aquatic Mammals 3:7-18. [ Links ]
4. FERIGOLO J. 1985. Evolutionary trends of the histological pattern in the teeth of Edentata (Mammalia). Archives of Oral Biology 30:71-82. [ Links ]
5. KALTHOFF DC. 2011. Microstructure of dental hard tissues in fossil and recent Xenarthrans (Mammalia: Folivora and Cingulata). Journal of Morphology 272:641-661. [ Links ]
6. LOCH C, PC SIMÓES-LOPES, and CJ DREHMER. 2010. Numerical anomalies in the dentition of southern fur seals and sea lions (Pinnipedia: Otariidae). Zoologia 27:477-482. [ Links ]
7. MCAFEE RK. 2007. Reassessing the taxonomy and affinities of the Mylodontinae sloths, Glossotherium and Paramylodon (Mammalia: Xenarthra: Tardigrada). Doctoral Thesis, Northern Illinois University, DeKalb, Illinois. [ Links ]
8. MCAFEE RK. 2009. Reassessment of the cranial characters of Glossotherium and Paramylodon (Mammalia: Xenarthra: Mylodontidae). Zoological Journal of the Linnean Society 155:885-903. [ Links ]
9. MCDONALD HG. 1978. A supernumerary tooth in the ground sloth, Megalonyx (Edentata Mammalia). Florida Scientist 41:12-14.
10. MILES AEW and C GRIGSON. 1990. Colyer's variations and diseases in the teeth of animals. Revised edition. Cambridge University Press, Cambridge. [ Links ]
11. NAPLES VL. 1982. Cranial osteology and function in the tree sloths, Bradypus and Choloepus. American Museum Novitates 2739:1-41. [ Links ]
12. NAPLES VL. 1995. The artificial generation of wear patterns on tooth models as a means to infer mandibular movement during feeding in mammals. Pp. 137-150, in: Functional Morphology in Vertebrate Paleontology (JJ Thomason, ed.). Cambridge University Press, Cambridge. [ Links ]
13. PUJOS F, G DE IULIIS, and BM QUISPE. 2011. Hiskatherium saintandrei gen. and sp. nov.: An unusual megatheroid sloth from the middle Miocene of Quebrada Honda (Bolivia). Journal of Vertebrate Paleontology 31:1131-1149. [ Links ]
14. SCHMIDT WJ and A KEIL. 1971. Polarizing microscopy of dental tissues: Theory, methods and results. Pergamon Press, New York. [ Links ]
15. SHOSHANI J, CA GOLDMAN, and JGW THEWISSEN. 1988. Orycteropus afer. Mammalian Species 300:1-5. [ Links ]
16. WETZEL RM. 1985a. The identification and distribution of Recent Xenarthra (=Edentata). Pp. 5-21, in: The Evolution and Ecology of Armadillos, Sloths, and Vermillinguas (GG Montgomery, ed.). Smithsonian Institution Press, Washington D.C. [ Links ]
17. WETZEL RM. 1985b. Taxonomy and distribution of armadillos, Dasypodidae. Pp. 23-46, in: The Evolution and Ecology of Armadillos, Sloths, and Vermillinguas (GG Montgomery, ed.). Smithsonian Institution Press, Washington D.C. [ Links ]
18. WOLSAN M. 1984. The origin of extra teeth in mammals. Acta Theriologica 29:128-133. [ Links ]

