










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383
Mastozool. neotrop. vol.20 no.1 Mendoza jun. 2013
ARTÍCULO
Una nueva especie de musaraña del género Cryptotis (Soricomorpha: Soricidae) de la Serranía del Litoral en el norte de Venezuela
Marcial Quiroga-Carmona
Departamento de Biología, Facultad Experimental de Ciencias y Tecnología, Universidad de Carabobo, Valencia 2005, Venezuela [Correspondencia: <marcialquiroga@gmail.com>].
Recibido 9 octubre 2012.
Aceptado 25 febrero 2013.
Editor asociado: P Teta
RESUMEN. La presencia del género Cryptotis en la Serranía del Litoral (norte de Venezuela), se conocía sólo por un ejemplar colectado en El Junquito hace más de 40 años, y referido para entonces a C. thomasi. Recientemente, fueron capturados dos nuevos ejemplares en una localidad cercana a la del primer hallazgo (15 km aproximadamente). Con la revisión y comparación morfológica (craneana y corporal) de los tres ejemplares ahora disponibles, se determinó que esta población representa una especie nueva, perteneciente al grupo de C. thomasi, y que puede ser diferenciada por la siguiente combinación de características: surco nasolabial con una verruga larga y delgada, extendida desde la base del rinario hasta la boca; dedos con escamas granulares bien desarrolladas y yuxtapuestas, arregladas en tres líneas longitudinales; cola con escamas grandes (26 por cm); foramen lacrimal ancho y profundo; proceso angular recto, corto y ancho; proceso coronoides alto e inclinado hacia adelante. Esta población representa la segunda especie de Cryptotis con distribución no andina en América del Sur y eleva a seis el número de mamíferos endémicos de la Cordillera de la Costa, ecorregión venezolana que actualmente enfrenta graves problemas de conservación por la pérdida de hábitat.
ABSTRACT. A new species of shrew of the genus Cryptotis (Soricomorpha: Soricidae) from the Serranía del Litoral, Northern Venezuela. The presence of the genus Cryptotis in the Serrania del Litoral (north of Venezuela), is known only by a specimen collected in El Junquito more than 40 years ago, referred by this time to C. thomasi. Recently, two new specimens were collected in a locality near to the first finding (approximately 15 km). With the revision and morphological comparison (skull and body) of the three specimens now available, it was determined that this population represents a new species belonging to the C. thomasi group, and can be differentiated by the following combination of characteristics: philtrum with a long and thin wart, extending from the base of the rhinarium to the mouth; toes with well developed granular scales juxtaposed and arranged in three longitudinal lines; tail with large scales (26 cm); lacrimal foramen wide and deep; angular process straight, short and wide; high coronoid process and tilted forward. This population represents the second species of Cryptotis with non-Andean distribution in South America, and increases to five the number of endemic mammals of the Cordillera de la Costa, Venezuelan ecoregion that currently faces serious problems of conservation by habitat loss.
Palabras clave: Cordillera de la Costa; Eulipotyphla; Monumento Natural Pico Codazzi; Musarañas neotropicales.
Key words: Cordillera de la Costa; Eulipotyphla; Monumento Natural Pico Codazzi; Neotropical shrews.
INTRODUCCIÓN
El género Cryptotis Pomel, 1848 (Soricomorpha: Soricidae) está constituido por musarañas pequeñas, con el rostro cónico y alargado, ojos y orejas reducidas, extremidades cortas y hábitos semifosoriales (Hutterer, 1985, 2005; Linares, 1998; Díaz de Pascual y De Ascenção, 2000; Woodman, 2011). Sus especies están distribuidas de forma discontinua desde Canadá hasta Perú y habitan diferentes ecosistemas desde el nivel del mar hasta zonas de alta montaña (Woodman y Péfaur, 2008).
El género Cryptotis es reconocido como un grupo monofilético, cercanamente relacionado a los géneros Blarina (Ohdachi et al., 2006) y Paracryptotis† (Choate, 1970). Actualmente comprende 37 especies, tres fósiles y 34 vivientes (Repenning, 1967; Reumer, 1998; Hutterer, 2005; Woodman, 2010, 2011; Quiroga-Carmona y Molinari, 2012). Los tratamientos taxonómicos recientes reúnen a las especies vivientes en cuatro grupos informales propuestos con base en caracteres dentarios, craneanos, postcraneanos y externos (Choate, 1970; Woodman y Timm, 1993, 1999, 2000; Woodman, 1996, 2002, 2010; Woodman et al., 2003). Estos son: C. mexicanus (presente en México y el norte de América Central), C. nigrescens (distribuido desde México hasta Colombia), C. parvus (con ocurrencia desde Canadá hasta Costa Rica) y C. thomasi (exclusivo de Sudamérica) (Woodman y Péfaur, 2008).
En América del Sur Cryptotis está representado por especies de los grupos C. nigrescens y C. thomasi, las que habitan diferentes tipos de ecosistemas por encima de los 1200 m (Woodman y Péfaur, 2008). Las especies pertenecientes al grupo C. nigrescens sólo están presentes en la Cordillera Oriental (C. colombianus) y Central (C. brachyonyx) de Colombia (Woodman y Timm, 1993; Woodman, 2003). Por su parte, las 10 especies pertenecientes al grupo de C. thomasi están distribuidas en alopatría desde la Cordillera de la Costa en el norte de Venezuela hasta la Depresión de Huancabamba en el norte de Perú (Woodman y Péfaur, 2008; Quiroga-Carmona y Molinari, 2012).
En Venezuela se reconocen tres especies de Cryptotis (Hutterer, 2005; Quiroga-Carmona y Molinari, 2012): C. tamensis presente en Colombia y Venezuela y cuya distribución en el país se restringe al Páramo del Tamá (Macizo del Tamá; Woodman, 2002), C. meridensis que es endémica de la Cordillera de Mérida, distribuida desde el norte de la Depresión del Táchira hasta el Macizo de Guaramacal (Woodman y Díaz de Pascual, 2004) y C. aroensis que es exclusiva de la Sierra de Aroa (Quiroga-Carmona y Molinari, 2012). Adicionalmente, en Venezuela también se conoce la presencia del género en la Serranía del Litoral a través de un reporte de Ojasti y Mondolf (1968), con un ejemplar colectado en El Junquito. Posteriormente, la presencia de Cryptotis en la vertiente venezolana de la Sierra de Perijá fue sugerida por Duarte y Viloria (1992), basándose en un ejemplar colectado en el Departamento del César en Colombia. Ambos registros fueron realizados con base en ejemplares únicos que, oportunamente, fueron referidos como C. thomasi.
La Serranía del Litoral forma parte del sistema montañoso de la Cordillera de la Costa. Cadena montañosa que se encuentra ubicada en el norte de Venezuela y se extiende desde la Sierra de Aroa hasta la Península de Paria, dividida en dos secciones principales -Cordillera Occidental y Cordillera Oriental- por la Depresión de Unare (Meier, 2011). La Cordillera Occidental está formada por tres serranías, la Sierra de Aroa, que es la sección más occidental, y está aislada del resto de la cordillera por la Depresión de Yaracuy, la cual da continuación a dos serranías paralelas -Serranía del Litoral y Serranía del Interior- separadas entre sí por los valles de Caracas, Nirgua y Valencia (Valles Centrales) y la Cordillera Oriental que está formada por las penínsulas de Araya y de Paria, y la Serranía de Turimiquire (IGVSB, 2003).
El reporte realizado por Ojasti y Mondolf (1968) representa el primer y único registro conocido hasta ahora para Cryptotis en la Serranía del Litoral. Recientemente fueron colectados dos nuevos ejemplares en una localidad a 15 km aproximadamente del sitio del primer registro. La revisión morfológica (craneana y corporal) de los tres ejemplares ahora disponibles y su comparación con ejemplares del Páramo del Tamá (C. tamensis), la Cordillera de Mérida (C. meridensis) y la Sierra de Aroa (C. aroensis), junto con datos de C. thomasi, demostraron que esta población no es asignable a C. thomasi y representa, en cambio, una especie nueva para la ciencia cuya descripción es el objetivo de este trabajo.
MATERIALES Y MÉTODOS
Se examinaron 66 ejemplares de Cryptotis (Apéndice) procedentes del Páramo del Tamá (C. tamensis), la Cordillera de Mérida (C. meridensis), la Sierra de Aroa (C. aroensis) y la Serranía del Litoral (Cryptotis sp. nov.). Estos animales se encuentran depositados en las siguientes instituciones de Venezuela: Museo de la Estación Biológica de Rancho Grande (EBRG), Museo de Biología de la Universidad Central de Venezuela (MBUCV), Colección de Vertebrados de la Universidad de los Andes (CVULA) y Museo de Historia Natural La Salle (MHNLS). Adicionalmente, se consultaron la descripción original (Merrian, 1897) y otras complementarias (Woodman, 1996; Vivar et al., 1997) de C. thomasi.
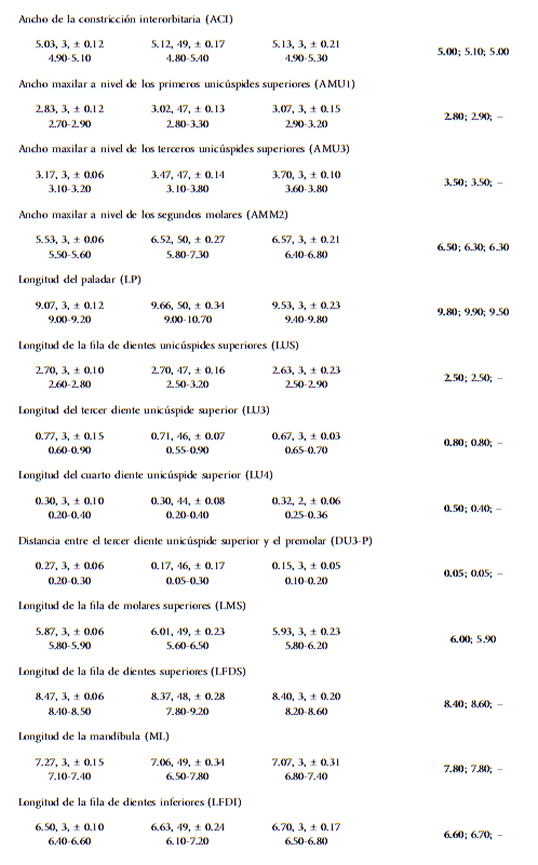
Para la descripción de Cryptotis sp. nov. se empleó la fórmula dentaria propuesta por Choate (1970), la nomenclatura dentaria descrita por Reumer (1984), la topografía craneana y mandibular descrita por Gaughran (1954) y la terminología cromática de Smithe (1975). Las medidas externas (Tabla 1) se tomaron con una regla con 0.5 mm de apreciación, o directamente de las etiquetas de museo. Para los cráneos, las 21 medidas fueron tomadas siguiendo la metodología empleada por Quiroga-Carmona y Molinari (2012), usando un micrómetro ocular con 0.01 mm de apreciación, adaptado a un microscopio estereoscópico binocular, a excepción de la longitud cóndilobasal y el ancho craneano, medidas que fueron tomadas con un calibre digital con apreciación de 0.01 mm. Para la descripción de los caracteres morfológicos se tomó en cuenta la variabilidad intragenérica de Cryptotis, por lo que los adjetivos empleados en la descripción de la especie nueva hacen referencia al estado de cada rasgo en función de su variabilidad dentro del género. Todas las medidas son presentadas en milímetros (mm) y el peso en gramos (g). Los acrónimos para las medidas utilizadas en el texto se describen en la Tabla 1.
Tabla 1
Medidas externas y craneanas de especies de Cryptotis con sus estadísticos descriptivos (media, n, desviación estándar y rango). Son presentadas las medidas de la serie tipo de C. venezuelensis sp. nov. Para una mayor serie de medidas de C. tamensis y C. thomasi ver Woodman (1996, 2002; tabla 1) y Vivar et al., (1997, tabla 1) respectivamente. Todas las medidas son presentadas en milímetros (mm) y el peso en gramos (g).


Para el análisis morfométrico sólo se consideraron ejemplares con cráneos completos (44 de C. meridensis, 3 de C. aroensis, 3 de C. tamensis y 2 de Cryptotis sp. nov.). Cada una de las medidas cráneo-mandibulares fue individualmente examinada con un análisis de varianza (ANOVA), para determinar cuáles son diferentes a nivel interespecífico. Solo las variables cráneo-mandibulares que resultaron significativamente diferentes (P ≤ 0.05) entre las especies se incluyeron en un análisis de componentes principales, realizado a partir de una matriz de correlación construida con variables transformadas con logaritmo. Los análisis estadísticos y gráficos fueron realizados con el software SPSS versión 15.
RESULTADOS
Se reconoce la especie del género Cryptotis presente en la Serranía del Litoral como un miembro del grupo C. thomasi por exhibir las siguientes características morfológicas: cuerpo de talla mediana (LCC: 72-84 mm, intervalo para el grupo 58-102 mm); cola mediana (43% de la LCC, intervalo para el grupo 28-60%); primer molar superior con el elemento anterior del ectolofo más pequeño que el elemento posterior y el borde anterior del proceso coronoides unido a la rama horizontal de la mandíbula en un ángulo bajo (cf. Woodman y Péfaur, 2008). A su vez, la nueva forma puede ser reconocida por una combinación única de caracteres que se describe a continuación.
Cryptotis venezuelensis sp. nov.
(Figs. 1, 2 y 4)

Fig. 1. Cryptotis venezuelensis sp. nov., holotipo (EBRG 27336), fotografiado en el momento de su captura.

Fig. 2. Cryptotis venezuelensis sp. nov., holotipo (EBRG 27336), desde arriba hacia abajo, vistas lateral, dorsal y ventral del cráneo y lateral de la mandíbula. Escala = 5 mm.
Cryptotis thomasi: Ojasti y Mondolf 1968: 439.
Cryptotis meridensis: Hutterer 1993: 108 (en parte).
Cryptotis meridensis meridensis: Linares 1998: 106 (en parte).
Holotipo-EBRG 27336, hembra adulta colectada el 5 de noviembre de 2010 por M. Quiroga-Carmona (Fig. 1); se conserva cráneo y mandíbulas en buenas condiciones (Fig. 2) y cuerpo completo en etanol.
Localidad tipo-Sector Cerro Geremba del Monumento Natural Pico Codazzi, Municipio Tovar, Estado Aragua, Venezuela (coordenadas geográficas: 10° 24' 36" N, 67° 13' 16" W), 2238 m de elevación (localidad 2 en Fig. 3).

Fig. 3. Mapa que muestra la distribución de las especies del género Cryptotis presentes en el noroeste de América del Sur (Colombia y Venezuela). Referencias: C. aroensis = triángulos invertidos, C. meridensis = círculos, C. tamensis = cuadrados, C. thomasi = triángulos y C. venezuelensis sp. nov. = estrellas. Para la referencia numérica de las localidades, véase el Apéndice. Las localidades en Colombia son las registradas por Woodman (2002) para C. tamensis y C. thomasi. Las áreas por encima de 700 m de elevación son mostradas en gris claro, las áreas por encima de 1200 m de elevación son mostradas en gris oscuro y los asteriscos representan la ubicación de los Valles Centrales.
Paratipos-EBRG 27337, macho adulto colectado el 6 de noviembre de 2010 por M. Quiroga-Carmona en la localidad tipo, incluye cráneo y mandíbulas en buenas condiciones y cuerpo completo preservado en etanol. MBUCV 402, ejemplar adulto de sexo desconocido colectado en julio de 1962 por A. Gamero en El Junquito, Municipio Libertador, Distrito Capital, Venezuela (10° 27' 36" N, 67° 05' 16" W, 2100 m; localidad 1, Fig. 3), incluye cráneo en malas condiciones (sin mandíbulas, dientes unicúspides superiores, molares superiores y procesos timpánicos) y esqueleto parcial (escápula derecha y algunas vertebras caudales).
Distribución-Solo es conocida de los bosques nublados de la Serranía del Litoral en el norte de Venezuela, entre los 2100 m y 2238 m de elevación (Fig. 3).
Diagnosis-Una especie de Cryptotis perteneciente al grupo C. thomasi, que se caracteriza por la siguiente combinación de caracteres morfológicos: tamaño mediano (LCC: 72-84 mm, Peso: 9-11 g); pelaje dorsal gris oscuro con lustre marrón oscuro; vientre de color gris más pálido que el dorso; surco nasolabial con una verruga larga y delgada, extendida desde la base del rinario hasta el borde del labio superior; parte ventral de los dedos con escamas granulares bien desarrolladas y yuxtapuestas, arregladas en tres líneas longitudinales; escamas de la cola grandes (26 por cm); foramen lacrimal ancho y profundo; proceso angular recto, corto y ancho; proceso coronoides inclinado hacia delante.
Diagnosis-A Cryptotis species belong to C. thomasi group and characterized by the following combination of morphological characters: medium sized (head and body length: 72-84 mm, body mass: 9-11 g); dorsal pelage dark gray with dark brown luster; ventral part gray, paler than dorsum; philtrum with a long and narrow wart, extending from the base of the rhinarium to the edge of the upper lip; ventral part of the fingers whit granular, well developed and juxtaposed scales, arranged in three longitudinal lines; tail scales large (26 per cm); lacrimal foramen wide and deep; angular process straight, short and wide; coronoid process tilted forward.
Descripción-Una musaraña de tamaño mediano para el género Cryptotis (LCC: 72-84 mm, Peso: 9-11 g). Pelaje dorsal gris oscuro con brillo marrón oscuro. Vientre más pálido que el dorso, con pelos bicoloreados de base gris oscuro y punta marrón. Surco nasolabial con una verruga larga y delgada, que se extiende desde la base del rinario hasta la boca (Fig. 4). Orejas con pinnas más altas que anchas y cubiertas por el pelaje, con el hélix, antihélix, antitrago y la fosa escafoidea poco desarrolladas. Vibrisas mistaciales extendidas hasta el borde anterior de la oreja. Parte ventral de los dedos de las patas, con escamas granulares bien desarrolladas y yuxtapuestas, arregladas en tres líneas longitudinales (Fig. 4). Garras largas y poco curvas. Almohadilla tenar de las patas anteriores y posteriores, de forma semitriangular y en posición posterior respecto a la almohadilla hipotenar. Otras almohadillas de las patas claramente definidas y redondeadas. Cola mediana (42.94% de la LCC), con escamas grandes (26 escamas por cm), pelaje denso y gris, contrastante entre la parte dorsal y ventral.

Fig. 4. Cryptotis venezuelensis sp. nov, detalles morfológicos (EBRG 27336): arriba, vista ventral del hocico, nótese la verruga presente en el surco nasolabial; abajo, planta de la pata mostrando la disposición de las escamas de los dedos.
Rostro moderadamente largo (45.84% de la LCB). Paladar moderadamente ancho (65.41% de la LP). Región interorbitaria relativamente ancha (23.70% de la LCB). Placa cigomática moderadamente ancha (10.52% de la LCB y 22.95% LP), con el borde anterior alineado con el metacrista del primer molar superior y el borde posterior alineado con el metacono del tercer molar superior. Proceso maxilar continuo con el borde posterior del paladar, extendido hasta el borde posterior del tercer molar superior. Con dos forámenes dorsales a lo largo de la sutura entre el frontal derecho e izquierdo (los dos paratipos exhiben esta condición y el holotipo sólo posee un foramen). Foramen oval ancho. Petromastoideo largo, con el proceso anterior corto y delgado y un foramen pequeño ubicado en el borde posterior del proceso timpánico (en uno o ambos lados del cráneo). Mandíbula con la región posterior corta (65.38% de la LM) y la escotadura sigmoidea inferior somera. Proceso coronoides alto e inclinado hacia delante. Proceso angular corto, ancho en la base y con la punta redondeada.
Dientes unicúspides superiores no bulbosos, con el cíngulo bajo y bien definido y las cúspides posterolinguales completamente desarrolladas. Fila superior de dientes unicúspides moderadamente larga (11.77% de la LCB). Cuarto unicúspide superior de tamaño mediano (56.25% de la LU3), en posición lingual y poco visible en vista lateral del cráneo. Espacio entre el tercer unicúspide superior y el premolar superior muy reducido (DU3-P: 0.17 mm), por lo que parecen estar en contacto, haciendo que el cuarto unicúspide superior sea escasamente visible en vista lateral del cráneo. Premolar superior con el parastilo y el protocono poco desarrollado y sin pigmentación. Primer molar superior con el elemento anterior del ectolofo menor que el elemento posterior. Crestas del protocono y el hipocono del primer y segundo molar superior pigmentadas. Tercer molar superior complejo, con el postcentrocrista desarrollado posteriormente y con mesostilo, metacono e hipocono presentes. Entocónidos del primer y segundo molar inferior con las cúspides redondeadas. Talónido del tercer molar inferior bien desarrollado, con un hipocónido conspicuo y un entocónido pequeño.
Etimología-Derivado del nombre Venezuela (Venezuel[a] + ensis [terminación en latín que denota lugar de origen]), país de colecta de la serie tipo.
Análisis morfométrico-El análisis estadístico de las muestras estudiadas determinó que 8 de 21 variables cráneo-mandibulares son significativamente diferentes (P ≤ 0.05) a nivel interespecífico (Tabla 2). Estas variables fueron empleadas para construir una matriz de correlación analizada con un análisis de componentes principales, cuyos tres primeros componentes explican el 68% de la varianza, siendo AMU1, AMU2, AMM2, LU4 y DU3-P las variables que mayor información aportan al sistema (Tabla 3).
Tabla 2
Resultados obtenidos en el análisis de varianza realizado para evaluar diferencias significativas (P≤ 0.05, en negrita) a nivel interespecífico en Cryptotis, en función de las medidas cráneomandibulares tomadas. Acrónimos como en Tabla 1.

Tabla 3
Resultados obtenidos en el análisis de componentes principales realizado a partir de 8 variables cráneo-mandibulares. Acrónimos como en Tabla 1.

En la Fig. 5 se muestran los gráficos de dispersión realizados con los tres primeros componentes principales obtenidos. En estos, C. aroensis aparece siempre discriminada a lo largo del primer componente principal, al igual que C. venezuelensis sp. nov., cuya separación se da a lo largo del tercer componente principal. Cryptotis aroensis es la especie incluida en el análisis con el menor ancho maxilar a nivel de los terceros unicúspides y segundos molares (Tabla 1), variable que a su vez posee el mayor peso en el primer componente principal (Tabla 3). En el caso de C. venezuelensis sp. nov., esta especie posee en promedio la mayor longitud del cuarto unicúspide superior (Tabla 1), que es la variable con mayor aporte al tercer componente principal. Estos resultados demuestran la diferenciación morfométrica existente entre C. aroensis y C. venezuelensis sp. nov. y de estas especies frente al resto de las especies de Cryptotis incluidas en el análisis. El solapamiento existente entre C. meridensis y C. tamensis refleja la similitud morfométrica existente entre ambas especies, hecho que ha sido anteriormente reportado por Woodman (2002).

Fig. 5. Dispersión de individuos de Cryptotis en los tres primeros componentes principales a partir de una matriz de correlación de 8 variables cráneo-mandibulares. Referencias: C. aroensis = cruces, C. meridensis = cuadrados vacíos, C. tamensis = triángulos y C. venezuelensis sp. nov. = cuadrados rellenos.
Comparaciones morfológicas-Cryptotis venezuelensis sp. nov. se puede diferenciar de las especies del grupo de C. thomasi geográficamente más cercanas (C. aroensis, C. meridensis, C. tamensis y C. thomasi) por las siguientes características: pelaje dorsal gris oscuro con lustre marrón (marrón grisáceo en C. aroensis, gris a gris oscuro en C. meridensis, marrón chocolate en C. tamensis y marrón medio a marrón oscuro en C. thomasi); orejas con las estructuras del pabellón externo (hélix, antihélix, antitrago y fosa escafoidea) poco desarrolladas (bien desarrolladas en C. aroensis); surco nasolabial con una verruga larga y delgada, extendida a lo largo de toda su longitud (en C. aroensis esta verruga está restringida a la parte anterior del surco nasolabial y en C. meridensis y C. tamensis la verruga es pequeña y de forma ovalada, y está ubicada en la parte más anterior del surco y en contacto con el rinario); parte ventral de los dedos con escamas granulares yuxtapuestas arregladas en tres hileras longitudinales (en C. aroensis, C. meridensis y C. tamensis las escamas están arregladas en dos hileras); cola larga, LC: 42.94% de la LCC (corta en C. thomasi, LC: 29.54% de la LCC), y con escamas grandes, 26 escamas por cm (pequeñas, 30 escamas por cm o menos en C. aroensis, C. meridensis y C. tamensis); foramen lacrimal ancho y profundo (estrecho y somero en C. meridensis, C. tamensis y C. thomasi, ver fg. 2 de Woodman y Díaz de Pascual, 2004; fg. 1 de Woodman, 2002 y fg. 1 de Vivar et al., 1997, respectivamente); foramen del borde posterior del proceso timpánico del petromastoideo pequeño y poco visible (grande y claramente visible en C. thomasi); paladar ancho, AMU3: 35.97% de la LP y AMM2: 65.41% de la LP (paladar angosto en C. aroensis, AMU3: 34.95% de la LP y AMM2: 60.97% de la LP); proceso coronoides alto e inclinado hacia delante (bajo e inclinado hacia atrás o recto en todas las especies incluidas en la comparación); proceso angular corto y ancho (largo y delgado en todas las especies incluidas en la comparación); entocónido del tercer molar inferior presente (ausente en C. meridensis y C. thomasi); dientes unicúspides superiores robustos y no bulbosos, con las cúspides posterolinguales desarrolladas (gráciles y no bulbosos, con las cúspides posterolinguales poco desarrolladas o ausentes en C. meridensis y C. tamensis); cuarto diente unicúspide superior mediano y poco visible en vista lateral del cráneo (de tamaño pequeño en C. aroensis y C. tamensis y grande en C. thomasi, y en las tres especies completamente visible); tercer diente unicúspide superior y premolar superior a menos de 0.1 mm de separación (en todas las especies incluidas en la comparación, ambos dientes están a más de 0.1 mm de separación).
Observaciones-Además de los genitales, no fueron observadas características alternativas que permitieran diferenciar entre ambos sexos, por lo que aparentemente Cryptotis venezuelensis sp. nov. carece de dimorfismo sexual. Esto es consistente con lo descrito para C. aroensis, C. meridensis y C. tamensis (ver Durant y Péfaur, 1984; Woodman, 2002; Woodman y Díaz de Pascual, 2004; Quiroga-Carmona y Molinari, 2012).
DISCUSIÓN
La presencia del género Cryptotis en la Serranía del Litoral había sido considerada por mucho tiempo un hecho enigmático, ya que desde el registro de Ojasti y Mondolf (1968) no habían sido colectados nuevos ejemplares de esta población a pesar de los posteriores inventarios de fauna realizados en esta serranía (Handley, 1976; Fernández-Badillo y Ulloa, 1990; Rivas y Salcedo, 2005). El animal en el que se basó el primer registro, fue entregado a A. Gamero por un amigo que indicó su captura en El Junquito, y fue identificado siguiendo el esquema clasificatorio propuesto por Cabrera (1958) para el género Cryptotis en América del Sur. Para entonces, no se contaba en Venezuela con colecciones de referencia de este género, lo que limitó la posibilidad de realizar una revisión taxonómica exhaustiva de esta población (J. Ojasti, com. pers.). Con la colecta de los dos nuevos ejemplares y la descripción de Cryptotis venezuelensis sp. nov. se ratifica la presencia del género Cryptotis en la Serranía del Litoral y se dilucida la situación taxonómica de esta población, la que se mantuvo por más de 40 años sin ser esclarecida.
Los bosques nublados de la Cordillera de la Costa se encuentra entre los 1700 m y 2400 m de elevación y se caracterizan por la presencia de diferentes plantas epífitas (familias Bromeliaceae y Orchidaceae), abundantes helechos arborescentes (géneros Cyathea, Elaphoglosum y Pteridium) y la presencia de las familias Annonaceae, Bignonaceae y Sapindaceae, siendo Podocarpaceae la familia más importante y representativa de estos ecosistemas (Meier, 2011). Estas características concuerdan con lo observado en el bosque de Cerro Geremba (Fig. 6) y en menor medida con algunas de las características del hábitat descrito por Díaz de Pascual (1993) y Woodman y Díaz de Pascual (2004) para C. meridensis. Este hecho es de esperarse, ya que en la Cordillera de la Costa son encontrados elementos botánicos típicos de los Andes Venezolanos, tales como Ceroxylon ceriferum, C. alpinum, Geissanthus fragrans, Libanothamnus neriifolius, Montanoa quadrangularis, Podocarpus oleifolius, Prumnopitys harmsianus y Ruagea pubescens (Meier, 2011). Otras de las características resaltantes del bosque de Cerro Geremba, es la presencia de mucha materia orgánica y humedad en el suelo. Estas condiciones favorecen la proliferación de varios grupos de invertebrados (e. g. Coleoptera, Gastropoda, Isopoda, Oligochaeta, Orthoptera y Tricladida) (Olson, 1994), los que fueron ampliamente observados durante el trabajo en campo, en la hojarasca y vegetación del suelo. Cryptotis meridensis está asociado positivamente a las dos características edafológicas mencionadas y sus presas consisten principalmente en invertebrados hipógeos y epígeos (e. g. Annelidae, Chilopoda, Coleoptera, Crustacea, Gastropoda, Lepidoptera y Orthoptera) (Díaz de Pascual, 1993; Díaz de Pascual y De Ascenção, 2000). Esta información y la similitud morfológica existente entre C. venezuelensis sp. nov. y C. meridensis, sugiere que posiblemente estas especies compartan hábitos alimenticios, así como la preferencia por el mismo tipo de hábitat. Esto es consistente con lo descrito por Quiroga-Carmona y Molinari (2012) sobre el hábitat de C. aroensis y da soporte a la propuesta hecha por estos autores sobre la restricción ecológica que los miembros del grupo C. thomasi tienen a habitar bosques nublados.
En América del Sur, Cryptotis puede ser considerado exclusivamente montano: todas las especies presentes en esta parte del continente habitan en ecosistemas ubicados por encima de los 1200 m de elevación (ver Woodman y Péfaur, 2008; Quiroga-Carmona y Molinari, 2012). Cryptotis venezuelensis sp. nov. sólo se conoce de dos localidades en la Serranía del Litoral, formación montañosa que se encuentra separada de la Sierra de Aroa por la Depresión de Yaracuy, de la Serranía del Interior por los Valles Centrales y de la Cordillera Oriental por la Depresión de Unare (Fig. 3), discontinuidades geográficas que no superan los 500 m de elevación (IGVSB, 2003). Tomando en cuenta el patrón de distribución alopátrico general de las especies del grupo C. thomasi y la situación geográfica del área de distribución conocida para C. venezuelensis sp. nov. podría considerarse a esta especie como endémica de la Serranía del Litoral, en vista de la discontinuidad del ambiente que esta especie habita en la Cordillera de la Costa.
Recientemente fue descrita C. aroensis, una especie que por ahora solo es conocida de la Sierra de Aroa (Quiroga-Carmona y Molinari, 2012), formación montañosa cuya historia geológica está relacionada a la de la Serranía del Litoral (Rodriguez-Olarte et al., 2009), con la que comparte condiciones climáticas y elementos botánicos (Hubber, 1997; Meier, 2011). Esta información permite pensar en la hipótesis de que el género Cryptotis colonizó la Serranía del Litoral desde la Cordillera de Mérida a través de la Sierra de Aroa, tal como es propuesto por Quiroga-Carmona y Molinari (2012). Dicho evento pudo haber sido favorecido por el descenso de los pisos térmicos, lo que ocurrió hace dos millones de años y trajo como consecuencia la continuidad de los bosques nublados y los páramos (Hooghiemstra et al., 2002; Hooghiemstra y van der Hammen, 2004), ecosistemas que representan el hábitat actual de las especies suramericanas de Cryptotis (Woodman y Péfaur, 2008).
La Cordillera de la Costa es reconocida como una de las ecorregiones de Venezuela con mayor nivel de endemismo (Péfaur y Rivero, 2000; Hilty, 2003; La Marca, 2003a, 2003b; Lentino, 2003; Ochoa y Aguilera, 2003), donde hasta ahora se conocían cinco especies endémicas de mamíferos (Cryptotis aroensis, Heteromys catopterius, Ichthyomys pittieri, Nephelomys caracolus y Rhipidomys venustus, Anderson y Gutiérrez, 2009; Quiroga-Carmona y Molinari, 2012), número que se eleva a seis con la descripción de C. venezuelensis sp. nov. En este sistema montañoso han sido establecidas varias áreas protegidas (Parque Nacional Yurubí, Parque Nacional Tirgua, Zona Protectora del Macizo de Nirgua, Parque Nacional San Esteban, Parque Nacional Henri Pittier, Monumento Natural Pico Codazzi, Parque Nacional Macarao, Parque Nacional Guatopo y Parque Nacional Waraira Repano), que conservan más de 350 000 ha de este sistema montañoso. Sin embargo, esta es una de las regiones del país con mayor grado de afectación ambiental, como resultado de las actividades humanas (ganadería, agricultura y expansión urbana), las que principalmente impactan áreas submontanas y subsecuentemente deterioran los ecosistemas a mayor altura, causando la degradación de los bosques nublados (Meier, 2011). En Venezuela los bosques nublados son clasificados como críticamente amenazados, según la Lista Roja de los Ecosistemas Terrestres de Venezuela, esto debido a la acelerada declinación que estos ecosistemas han experimentado en los últimos años (Rodriguez et al., 2010). Por esta razón Cryptotis venezuelensis sp. nov. puede ser considerada una especie en riesgo por la pérdida de hábitat, situación que deberá ser confirmada con estudios posteriores.
AGRADECIMIENTOS
Inicialmente quiero agradecer a la Fundación Provita por brindar parte de los fondos necesarios para la realización de este trabajo (revisión taxonómica y trabajo de campo), a través de la Iniciativa de Especies Amenazadas (IEA). A los curadores de las colecciones científicas visitadas durante la revisión taxonómica: F. Bisbal y J. Sánchez (EBRG), P. Soriano y J. Murillo (CVULA), C. Ferreira y M. Salazar (MBUCV), y A. Ferrer (MHNLS). A E. Blanco por la elaboración de las ilustraciones presentadas en este trabajo. A C. Do Nascimiento, S. Solari, R. Pine y B. Morey por las sugerencias dadas a las primeras versiones del manuscrito. A los editores U. Pardiñas y P. Teta por la atención y el respeto brindado. Especialmente quiero agradecer a J. Molinari por las recomendaciones y el apoyo brindado durante la preparación de este articulo y a E. Camargo, cuya ayuda fue indispensable para el éxito del trabajo en campo.
LITERATURA CITADA
1. ANDERSON RP y EE GUTIÉRREZ. 2009. Taxonomy, distribution, and natural history of the genus Heteromys (Rodentia: Heteromyidae) in central and eastern Venezuela, with the description of a new species from the Cordillera de la Costa. Pp. 33-93, en: Systematic mammalogy: Contributions in honor of Guy G. Musser (RS Voss y MD Carleton, eds.). Bulletin of American Museum of Natural History 331. [ Links ]
2. CABRERA A. 1958. Catálogo de los mamíferos de América del Sur. Vol I. Metatheria, Unguiculata, Carnivora. Revista del Museo Argentino de Ciencias Naturales Bernandino Rivadavia 4:1-307. [ Links ]
3. CHOATE JR 1970. Systematics and zoogeography of Middle American shrews of the genus Cryptotis. University of Kansas Publication, Museum of Natural History 19:195-317. [ Links ]
4. DÍAZ DE PASCUAL A. 1993. Caracterización del hábitat de algunas especies de pequeños mamíferos de la selva nublada de Monte Zerpa, Mérida. Ecotrópicos 6:1-9. [ Links ]
5. DÍAZ DE PASCUAL A y A DE ASCENÇÃO. 2000. Diet of the cloud forest shrew Cryptotis meridensis (Insectivora: Soricidae) in the Venezuelan Andes. Acta Teriologica 45:3-24. [ Links ]
6. DUARTE M y A VILORIA. 1992. Nuevo hallazgo de Cryptotis thomasi (Merriam, 1897) (Mammalia: Insectivora) en la Sierra de Perijá, Noreste de Colombia. Acta Científica Venezolana 43:240-242. [ Links ]
7. DURANT P y J PÉFAUR. 1984. Sistemática y ecología de la musaraña de Mérida, Soricidae: Insectivora, Cryptotis thomasi. Revista de Ecología, Conservación y Ornitología Latinoamericana 1:3-14. [ Links ]
8. FERNÁNDEZ-BADILLO A y G ULLOA. 1990. Fauna del Parque Nacional Henri Pittier, Venezuela: composición y diversidad de la mastofauna. Acta Científica Venezolana 41:50-63. [ Links ]
9. GAUGHRAN G. 1954. A comparative study of the osteology and myology of the craneal and cervical regions of the shrew, Blarina brevicauda and the mole, Scalopus aquaticus. Miscellaneous Publications of University of Michigan Museum of Zoology 80:1-85. [ Links ]
10. HANDLEY CO. 1976. Mammals of the Smithsonian Venezuelan project. Brigham Young University Science Bulletin (Biological series) 20 (5):1-91. [ Links ]
11. HILTY SL. 2003. Birds of Venezuela. 2nd ed. Princeton University Press, New Jersey. [ Links ]
12. HOOGHIEMSTRA H, T VAN DER HAMMEN y A CLEEF. 2002. Paleoecología de la flora boscosa. Pp. 43-58, en: Ecología y conservación de bosques neotropicales (MR Guariguata y GH Kattan eds.). Libro Universitario Regional, Cartago. [ Links ]
13. HOOGHIEMSTRA H y T VAN DER HAMMEN. 2004. Quaternary Ice-Age dynamics in the Colombian Andes: Developing an understanding of our legacy. Philosophical Transactions: Biological Sciences 359:173-181. [ Links ]
14. HUBBER O. 1997. Ambientes fisiográficos y vegetales de Venezuela. Pp. 279-298, en: Vertebrados actuales y fósiles de Venezuela (E La Marca, ed.). Museo de Ciencia y Tecnología, Mérida. [ Links ]
15. HUTTERER R. 1985. Anatomical adaptations of shrews. Mammal review 15:43-55. [ Links ]
16. HUTTERER R. 1993. Order Insectivora. Pp. 69-130, en: Mammal species of the world, a taxonomic and geographic reference (DE Wilson y DM Reeder, eds.). Segunda edicion. Smithsonian Institution Press, Washington. [ Links ]
17. HUTTERER R. 2005. Order Soricomorpha. Pp. 124-298, en: Mammal species of the world: a taxonomic and geographic reference (DE Wilson y DM Reeder, eds.). Tercera edicion. Johns Hopkins University Press, Baltimore. [ Links ]
18. INSTITUTO GEOGRÁFICO DE VENEZUELA SIMÓN BOLÍVAR (IGVSB). 2003. Mapa del relieve de Venezuela. Caracas. [ Links ]
19. LA MARCA E. 2003a. Anfibios. Pp. 582-695, en: Biodiversidad en Venezuela (MA Aguilera, A Azócar, y E González-Jiménez, eds.). Tomo II. Fundación Empresas Polar, Ministerio de Ciencia y Tecnología, Caracas. [ Links ]
20. LA MARCA E. 2003b. Reptiles. Pp. 596-608, en: Biodiversidad en Venezuela (MA Aguilera, A Azócar, y E González-Jiménez, eds.). Tomo II. Fundación Empresas Polar, Ministerio de Ciencia y Tecnología, Caracas. [ Links ]
21. LENTINO M. 2003. Aves. Pp. 610-648, en: Biodiversidad en Venezuela (MA Aguilera, A Azócar y E González-Jiménez, eds.). Tomo II. Fundación Empresas Polar, Ministerio de Ciencia y Tecnología, Caracas. [ Links ]
22. LINARES O. 1998. Mamíferos de Venezuela. Sociedad Conservacionista Audubon de Venezuela, Caracas. [ Links ]
23. MEIER W. 2011. Los Bosques Nublados de la Cordillera de la Costa en Venezuela. Pp. 106-121, en: Bosques de Venezuela, un homenaje a Jean Pierre Veillon (G Aymard, ed.). Biollania edición especial Nº 10: 10. [ Links ]
24. MERRIAM CH. 1897. Descriptions of five new shrews from Mexico, Guatemala and Colombia. Proceedings of the Biological Society of Washington 11:227-230. [ Links ]
25. OCHOA J y M AGUILERA. 2003. Mamíferos. Pp. 650-672, en: Biodiversidad en Venezuela (MA Aguilera, A Azócar y E González-Jiménez, eds.). Tomo II. Fundación Empresas Polar, Ministerio de Ciencia y Tecnología, Caracas. [ Links ]
26. OHDACHI S, M HASEGAWA, M IWASA, P VOGEL, T OSHIDA, LK LIN y H ABE. 2006. Molecular phylogenetics of soricid shrews (Mammalia) based on mitochondrial cytochrome B gene sequences: With special reference to the Soricinae. Journal of Zoology 270:177-191. [ Links ]
27. OJASTI J y E MONDOLFI. 1968. Capitulo noveno, Esbozo de la Fauna de Mamíferos de Caracas. Pp. 410-461, en: Ecología vegetal y fauna. Estudio de Caracas (M Crema, ed.). Vol. I. Caracas: Ediciones de la Biblioteca de la Universidad Central de Venezuela. [ Links ]
28. OLSON DM. 1994. The distribution of leaf litter invertebrates along a neotropical altitudinal gradient. Journal of Tropical Ecology 10:129-150. [ Links ]
29. PÉFAUR J y J RIVERO. 2000. Distribution, species– richness, endemism, and conservation of Venezuelan amphibians and reptiles. Amphibian & Reptile Conservation 2:43-70. [ Links ]
30. QUIROGA-CARMONA M y J MOLINARI. 2012. Description of a new shrew of the genus Cryptotis (Mammalia: Soricomorpha: Soricidae) from the Sierra de Aroa, an isolated mountain range in northwestern Venezuela, with remarks on biogeography and conservation. Zootaxa 3441:1-20. [ Links ]
31. REUMER J. 1984. Ruscinian and early Pleistocene Soricidae (Insectivora. Mammalia) from Tegelen (The Netherlands) and Hungary. Scripta Geologica 73:1-173. [ Links ]
32. REPPENING CA. 1967. Subfamilies and genera of the Soricidae. Classification, historical zoogeography, and temporal correlation of the shrews. U.S. Geological Survey Professional Paper 565:1-74. [ Links ]
33. RIVAS BA y MA SALCEDO. 2005. Lista actualizada de los mamíferos del Parque Nacional El Ávila, Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales 164:29-56. [ Links ]
34. RODRÍGUEZ JP, F ROJAS-SUAREZ y D GIRALDO-HERNÁNDEZ. 2010. Libro Rojo de los ecosistemas terrestres de Venezuela, primera edición. Provita, Lenovo, Shell de Venezuela, Caracas. [ Links ]
35. RODRÍGUEZ-OLARTE D, DC TAPHORN y J LOBÓN-CERVIÁ. 2009. Patterns of freshwater fishes of the Caribbean versant of Venezuela. Review of Hydrobiology 94:67-90. [ Links ]
36. SMITHE F. 1975. Naturalist´s color guide. American Museum of Natural History, New York. [ Links ]
37. VIVAR E, V PACHECO y M VALQUI. 1997. A new species of Cryptotis (Insectivora: Soricidae) from Northern Peru. American Museum Novitates 3202:1-15. [ Links ]
38. WOODMAN N. 1996. Taxonomic status of the enigmatic Cryptotis avia (Mammalia: Insectivora: Soricidae), with comments on the distribution of the Colombian small-eared shrews, Cryptotis colombiana. Proceedings of the Biological Society of Washington 109:409-418. [ Links ]
39. WOODMAN N. 2002. A new species of small-eared shrew from Colombia and Venezuela (Mammalia: Soricomorpha: Soricidae: Genus Cryptotis). Proceedings of the Biological Society of Washington 115:249-272. [ Links ]
40. WOODMAN N. 2003. A new small-eared shrew of the Cryptotis nigrescens-group from Colombia (Mammalia: Soricomorpha: Soricidae). Proceedings of the Biological Society of Washington 116:853-872. [ Links ]
41. WOODMAN N. 2010. Two new species of shrews (Soricidae) from the western highlands of Guatemala. Journal of Mammalogy 91:566-579. [ Links ]
42. WOODMAN N. 2011. Patterns of morphological variation amongst semifossorial shrews in the highlands of Guatemala, with the description of a new species (Mammalia, Soricomorpha, Soricidae). Zoological journal of the Linnean Society 163:267-288. [ Links ]
43. WOODMAN N, CA CUARTAS-CALLE y CA DELGADO. 2003. The humerus of Cryptotis colombiana and its bearing on the species' phylogenetic relationships (Soricomorpha: Soricidae). Journal of Mammal 84:832-839. [ Links ]
44. WOODMAN N y A DÍAZ DE PASCUAL. 2004. Cryptotis meridensis. Mammalian Species. American 761:1-5. [ Links ]
45. WOODMAN N y J PÉFAUR. 2008. Order Soricomorpha. Pp. 177-187, en: Mammals of South America, Vol. I: marsupials, xenarthrans, shrews and bats (AL Gardner, ed.). University of Chicago Press, Chicago, Vol.I. [ Links ]
46. WOODMAN N y RM TIMM. 1993. Intraspecific and interspecific variation in the Cryptotis nigrescens species complex of small-eared shrews (Insectivora: Soricidae), with the description of a new species from Colombia. Fieldiana Zoology 74:1-30. [ Links ]
47. WOODMAN N y RM TIMM. 1999. Geographic variation and evolutionary relationships among broad-clawed shrews of the Cryptotis goldmani-group (Mammalia: Insectivora: Soricidae). Fieldiana 91:1-35. [ Links ]
48. WOODMAN N y RM TIMM. 2000. Taxonomy and evolutionary relationships of Phillips' small-eared shrew, Cryptotis phillipsii (Schaldach, 1966), from Oaxaca, Mexico (Mammalia: Insectivora: Soricidae). Proceedings of the Biological Society of Washington 113:339-355. [ Links ]
Especímenes examinados y sus localidades de colecta (véase la Fig. 3).
C. venezuelensis sp. nov. (n =3): VENEZUELA. Vargas: El Junquito, 10°27' N 68°49' W, 2100 m: MBUCV 402 (Localidad 1 en Fig. 3). Aragua: Sector Cerro Geremba, Monumento Natural Pico Codazzi, 10°24' N 67°13' W, 2283 m: EBRG 27336-27337 (Localidad 2).
C. aroensis (n = 3): VENEZUELA. Yaracuy: Sierra de Aroa, Sector Las Cumaraguas, 10°22' N 68°49' W, 1730 m: CVULA 8546-8548 (Localidad 3).
C. meridensis (n = 57): VENEZUELA. Mérida: Páramo de Mucubají, Laguna Negra, 08° 47' N 70° 48' W, 3460 m: MBUCV 1882-1885, 2773; MHNLS 11221 (Localidad 4); km 36 of road Apartaderos-Barinas, 10 km E Santo Domingo, 08° 52' N 70° 37' W, 1670 m: CVULA 821 (Localidad 5); El Salvajal, 8 km W Mérida, 08° 36' N 71° 13' W, 2000 m: CVULA 2458 (Localidad 6); Monte Zerpa, 6 km N Mérida, 08° 38' N 71° 09' W, 2160 m: CVULA 1211, 5888, 6890, 6891 (Localidad 7); Escagüey, 1.5 km SSW Mucurubá, 08° 42' N 71° 00' W, 2200 m: CVULA 6899, 6900 (Localidad 8); La Carbonera, 14 km SE La Azulita, 08° 37' N 71° 22' W, 2250 m: CVULA 7912 (Localidad 9); NE of Laguna Brava, Páramo de Mariño, 7 km W Tovar, 08° 18' N 71° 49' W, 2300 m: CVULA 2352 (Localidad 10); Asentamiento Monterrey, 8 km NNE Mérida, 08° 41' N 71° 07' W, 2300 m: CVULA 3502, 7078 (Localidad 11); 5.5 km E and 2 km S Tabay, 08° 37' N 71° 02' W, 2630 m: EBRG 3639, 3640, 3641 (Localidad 12); Páramo de San José, 4.7 km SSE San José, 08° 20' N 71° 18' W, 3100 m: CVULA 8537-8545 (Localidad 13); 7.5 km E and 6 km S Tabay, 8° 35' N 71° 00' W, 3545 m: EBRG 3642 (Localidad 14); cercanías de la Laguna de Mucubají, Páramo de Mucubají, 08° 47' N 70° 49' W, 3600 m: CVULA 333-335, 337, 359, 421, 766, 874, 1168, 1486, 1487-1496, 1650, 1762, 8549; MHNLS 11220 (Localidad 15). Táchira: Cerro Alto Duque, 2.5 km SE El Cobre, 08° 01' N 72° 01' W, 2500 m: MBLUZ 167 (Localidad 16); Páramo el Zumbador, 10 km SSW El Cobre, 07° 58' N 72° 06' W, 2750 m: CVULA 5744 (Localidad 17).
C. tamensis (n = 3): VENEZUELA. Táchira: Buena Vista, cerca del Páramo del Tamá, 41 km SW San Cristóbal, 7°27' N 72° 22' W, 2460 m: EBRG 3643, 3644, 11743 (Localidad 18).

