










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383
Mastozool. neotrop. vol.21 no.2 Mendoza dic. 2014
ARTÍCULO
Nuevos datos morfológicos y una hipótesis filogenética para Phaenomys (Rodentia, Cricetidae)
Ulyses F. J. Pardiñas1, Guillermo D'Elía2, Gisele Lessa3, Marcelo Passamani4 y Pablo Teta1
1 Unidad de Investigación Diversidad, Sistemática y Evolución, Centro Nacional Patagónico, Casilla de Correo 128, 9120 Puerto Madryn, Chubut, Argentina [correspondencia: Ulyses F. J. Pardiñas <ulyses@cenpat.edu.ar>].
2 Instituto de Ciencias Ambientales y Evolutivas, Facultad de Ciencias, Universidad Austral de Chile, campus Isla Teja s/n, Valdivia, Chile.
3 Museu de Zoologia, Departamento de Biologia Animal, Universidade Federal de Viçosa, 36571-000 Viçosa, Minas Gerais, Brasil.
4 Setor de Ecologia, Universidade Federal de Lavras, 372000-000 Lavras, Minas Gerais, Brasil.
Recibido 3 julio 2014.
Aceptado 15 septiembre 2014.
Editor asociado: CA Galliari
RESUMEN.
Sobre la base de 2 ejemplares del roedor Phaenomys ferrugineus (Thomas), endémico de la Selva Atlántica, se aporta información novedosa sobre su anatomía y resultados del primer análisis filogenético basado en secuencias de ADN que incluye al género. Este sigmodontino se caracteriza por un cráneo con crestas supraorbitarias marcadas, un importante desarrollo de la región basicraneana, un patrón de circulación carotídea primitiva, tegmen timpánico presente, barra alisfenoidea ausente y mandíbula con gran desarrollo del sector donde inserta el músculo pterigoideo interno. Phaenomys presenta 12 costillas, un estómago unilocular-hemiglandular, carece de vesícula biliar y muestra un ciego de gran tamaño. Una hipótesis filogenética basada en datos moleculares combinados de 2 marcadores sugiere su condición de hermano del género Wiedomys. Se discute la plausible existencia de un clado que podría estar conformado por géneros endémicos de la región sudoriental del subcontinente.
ABSTRACT.
New morphological data and a phylogenetic hypothesis for Phaenomys (Rodentia, Cricetidae).
Based on 2 collected specimens of the rodent Phaenomys ferrugineus (Thomas), an endemic species of the Atlantic Forest, novelty information about its anatomy is provided as well as the results of the first phylogenetic analysis based on DNA sequences including this genus. This sigmodontine is characterized by a skull with marked supraorbitary crests, a well developed basicranial region, a primitive circulatory carotid pattern, the presence of a tegmen tympani, the absence of alisphenoid strut, and a mandible showing an important development of the region where the internal pterigoid muscle inserts. Phaenomys has 12 ribs, an unilocular-hemiglandular stomach, lacks gall bladder and has a large blind gut. A phylogeny based on the combined analysis of 2 molecular markers retrieves Phaenomys as sister to the genus Wiedomys. The plausible existence of a clade including endemic genera of the southeastern portion of the subcontinent, is discussed.
Palabras clave: Brasil; Juliomys; Thomasomyini; Wiedomys; Wilfredomys.
Key words: Brazil; Juliomys; Thomasomyini; Wiedomys; Wilfredomys.
INTRODUCCIÓN
El género monotípico Phaenomys, endémico de la Selva Atlántica en las sierras de Mantiqueira y Bocaina (Brasil; Passamani et al., 2011), contiene una especie de sigmodontino arborícola de tamaño grande y coloración dorsal rojiza brillante. Su conocimiento, pese a la temprana descripción original (Thomas, 1894, 1917), ha estado limitado por el escaso número de ejemplares colectados. Bonvicino et al. (2001) lo sintetizaron algo más de una década atrás y reportaron el cariotipo de la única especie conocida, P. ferrugineus (Thomas, 1894). Estos autores también discutieron -sobre las evidencias morfológicas reunidas- la posición filogenética de Phaenomys, señalando su condición de línea singular dentro de la radiación sigmodontina. Por su parte Pacheco (2003), en el único estudio filogenético que incluyó al género, basado en morfología, lo recuperó como un integrante de Thomasomyini. Otros autores han optado por mantenerlo -ante la carencia de información molecular para Phaenomys y debido a que la hipótesis de Pacheco (2003) ha sido contrastada negativamente por filogenias moleculares- como un Sigmodontinae incertae sedis (Smith y Patton, 1999; Musser y Carleton, 2005; D'Elía et al., 2007; D'Elía, en prensa).
Dos ejemplares de P. ferrugineus obtenidos recientemente permiten aportar información novedosa sobre la morfología interna y externa de la especie. Además, datos moleculares de uno de estos, en el contexto de un muestreo taxonómico denso, brindan una nueva hipótesis filogenética para este roedor. Ambos aspectos se desarrollan en esta contribución.
MATERIALES Y MÉTODOS
Los materiales estudiados consisten en un macho adulto, del cual se inspeccionó su cuerpo sin piel en fluido con el sistema digestivo completo, que pertenece a la Colección de Mamíferos de la Universidade Federal de Lavras, Minas Gerais (CMUFLA 904) y una hembra subadulta, conservada como piel disecada, esqueleto parcial y muestras de tejidos en alcohol y depositada en la Colección de Mamíferos del Museu de Zoologia, Departamento de Biologia Animal, Universidade Federal de Viçosa, Viçosa, Minas Gerais (con el número MZUFV 3400), ambos repositorios en Brasil. Mientras que el registro del CMUFLA 904, procedente de Hotel Casa Alpina, Itamonte, Minas Gerais, fue publicado previamente (Passamani et al., 2011), el MZUFV 3400 representa un dato novedoso. Este espécimen fue colectado en el Parque Estadual da Serra do Brigadeiro (20°43'01" S, 42°28'46" W, 1350 m) por uno de los autores (número de campo GL 625, fecha de colecta 14/11/2010) mediante una trampa de caída. El registro permite extender unos 170 km hacia el norte la distribución de Phaenomys y constituye la segunda localidad conocida para el género en el Estado de Minas Gerais (cf. Passamani et al., 2011). Todos los ejemplares estudiados en esta contribución se detallan en el Apéndice 1; en la descripción de la morfología dentaria se han seguido los conceptos y nomenclatura de Reig (1977).
Las secuencias del gen mitocondrial que codifica para el citocromo b (cit b) y del gen nuclear que codifica para la proteína de fijación del interfotorreceptor retinoide (IRBP) del ejemplar de P. ferrugineus MZUFV 3400 se obtuvieron siguiendo los protocolos detallados en Pardiñas et al. (2003) y D'Elía et al. (2006a), respectivamente. Las secuencias de ADN generadas fueron depositadas en Genbank (KM065876-7) e integradas a las matrices analizadas por Ventura et al. (2013), mediante un alineamiento realizado en Clustal X (Thompson et al., 1997) usando valores de defecto en los parámetros de alineación. Dichas matrices se analizaron independientemente mediante Máxima Verosimilitud (MV) e Inferencia Bayesiana (IB), usando los modelos de sustitución nucleotídica y protocolos analíticos detallados en Ventura et al. (2013). Además, se creó una tercera matriz donde se concatenaron los 2 genes; esta matriz fue analizada de igual manera que las matrices independientes, con la salvedad de que para cada partición se usó el modelo de evolución molecular correspondiente (ver detalles en Ventura et al., 2013).
RESULTADOS Y DISCUSIÓN
El espécimen CMUFLA 904 aporta información novedosa sobre la anatomía interna de Phaenomys. La disección de su estómago revela una condición unilocular-hemiglandular (Fig. 1) similar a la descripta para Thomasomys aureus por Carleton (1973:fig. 4C); sin embargo, en Phaenomys no se verifica una extensión de la pared glandular hacia la porción superior de la cavidad estomacal. El intestino grueso es proporcionalmente corto pero hay un ciego de enorme desarrollo, que recuerda a aquel exhibido por Reithrodon (U. Pardiñas, obs. pers.). No se detecta en Phaenomys vesícula biliar.

Fig. 1. Morfología del estómago de Phaenomys ferrugineus basada en el ejemplar CMUFLA 904: ambas mitades en vistas internas.
El espécimen MZUFV 3400 suma datos al conocimiento de Phaenomys tanto en aspectos de la morfología externa como así también de la anatomía craneana, dentaria y, por primera vez, del postcráneo. A fin de evitar redundancias con respecto a las descripciones aportadas por Bonvicino et al. (2001), aquí detallaremos solo los resultados novedosos (las medidas de este ejemplar se proveen en la Tabla 1). El pelaje dorsal presenta el rojizo brillante, característico de varios de los ejemplares disponibles, mientras que el vientre es de color crema claro, con una mancha pectoral ocre, generando un fuerte contraste dorso-ventral (Fig. 2, material suplementario). La cola es enteramente marrón oscura, cubierta de pelos híspidos y cortos, en grupos de 3 por cada escama y no más largos que 2 hileras de escamas, estas subrectangulares. La diferencia de coloración entre la cola y el resto del animal produce un contraste marcado. El pie es dorsalmente tricolor, rojizo en su parte más proximal, marrón oscuro en su porción metacarpal y progresivamente blanquecino hacia los dedos, para rematar en pelos ungueales blancos que cubren enteramente (aunque no sobrepasan) unas cortas garras no aquilladas. La morfología plantar -que describimos solo someramente por tratarse de una piel seca- revela un pie moderadamente ancho, con normal desarrollo de tenar e hipotenar y especial abultamiento de las 4 interdigitales y también de las callosidades, en particular los pulpejos. Este pie recuerda, tanto en su morfología como en color, a aquel de Juliomys (cf. Pardiñas et al., 2008:fig. 4).
Tabla 1. Medidas (en mm) externas y cráneo-dentarias (siguiendo las definiciones de Bonvicino et al., 2001) del ejemplar de Phaenomys ferrugineus MZUFV 3400.

En el cráneo (Fig. 3, material suplementario y Figs. 4A-4H) en vista dorsal llaman la atención unas crestas frontales que invaden la región supraorbitaria pero acaban antes de alcanzar la sutura coronal. Las mismas no coinciden, como es típico en otros sigmodontinos, con el borde superior de la órbita, sino que discurren dejando un espacio en su lateral externo y contribuyendo a acentuar una ya marcada depresión del frontal en su línea media (Figs. 4B-4C). La región interorbitaria y las órbitas adquieren, en conjunción con unos lagrimales bien desarrollados (Fig. 4B), una preponderancia en la norma dorsal que apenas supera el basicráneo. La caja craneana tiene una apariencia subrectangular generada por unos parietales alargados y el gran tamaño del interparietal. En la región rostral (Fig. 4A) destacan unas importantes fisuras nasolagrimales e, internamente, acompañan turbinales de buen desarrollo (Fig. 4D); el conjunto concluye con forámenes incisivos que cubren buena parte del paladar diastémico hasta alcanzar el plano de las raíces anteriores de los M1. Los arcos cigomáticos revelan jugales grandes y, en el escamoso, el proceso hamular es tan masivo que oblitera la fenestra subescamosal y casi también hace lo propio con el foramen postglenoideo (Fig. 4E). Dorsal al escamoso, la expresión lateral del parietal es mayúscula, generando hacia atrás una cresta lambdoidea prominente, que se continúa entre el escamoso y unas conspicuas cápsulas mastoideas. El paladar palatino es liso aunque muestra una línea de forámenes pareados hacia la sutura media, mientras que los molares se implantan en sendos procesos (Fig. 4G). Una gran fosa mesopterigoidea, con forma de lira y proceso medio acuminado, penetra el plano posterior de los M3, determinando un paladar corto (Fig. 4G). Mientras que su techo está completamente osificado, el foramen lacerado deja una importante abertura entre cada placa pterigoidea y su cápsula timpánica (Fig. 4H). Estas últimas son pequeñas, con importantes desarrollos del canal carotídeo y foramen estapedial, exposición del petrotimpánico y separadas por un basioccipital ancho y aquillado (Fig. 4H).
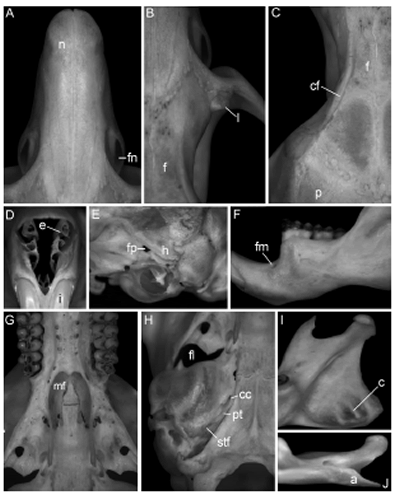
Fig. 4. Detalles de la anatomía craneana y mandibular de Phaenomys ferrugineus basados en el ejemplar MZUFV 3400: A. Rostro en vista dorsal; B. Sector de la muesca cigomática derecha en vista dorsal; C. Región interorbitaria en vista dorsal; D. Rostro en vista frontal; E. Región temporal en vista lateral; F. Rama horizontal del dentario izquierdo en vista labial; G. Sector palatino posterior en vista ventral; H. Bulla auditiva; I. Rama ascendente del dentario derecho en vista labial; J. Proceso angular del dentario derecho en vista ventral. Abreviaturas: a = proceso angular; c = "costillas" en la cara interna del proceso angular; cc = canal carotídeo; cf = cresta del frontal; e = etmoturbinales; f = frontal; fl = foramen lacerado; fm = foramen mentoniano; fn = fisura nasolagrimal; fp = foramen postglenoideo; h = proceso hamular del escamoso; l = lagrimal; mf = fosa mesopterigoidea; n = nasal, pt = petrotimpánico; stf = foramen estapedial.
En la mandíbula (Figs. 4F, 4I y 4J) destaca, además del contraste que producen los robustos molares e incisivo, el gran desarrollo del sector que corresponde a la inserción del músculo pterigoideo interno (Fig. 4I). No solo se observa una triple "costilla" o rugosidad en la parte interna de la lámina que forma el proceso angular -que visto ventralmente presenta un singular recorte (Fig. 4J)- sino que el foramen mandibular prácticamente desaparece de la visual en norma lateral y queda relegado a la cara ventral del estuche generado por el incisivo.
Los incisivos son muy robustos, opistodontes y con fisura en la dentina recta y corta (cf. Steppan, 1995). Los molares (Fig. 5) son braquiodontes y crestados, al menos en la condición de subadulto del MZUFV 3400 (contra la tipificación de aterrazados que dan Bonvicino et al., 2001). Las cúspides principales son opuestas, tanto en el M1 como en el m1, con una alternancia creciente en los M2/m2 y M3/m3. Tanto los molares superiores como los inferiores muestran sus flexos/idos linguales/ labiales cerrados por un cíngulo filoso de esmalte; mientras que en los superiores este cordón perimetral no parece tener engrosamientos, eliminando la posibilidad de confusión con enterostilos o protostilos, en los inferiores hay un leve abultamiento a nivel del hipofléxido, que podría resultar en la homología con un ectostílido (evidente en el m1, no así en los m2-m3). En el M1 el procíngulo es casi tan ancho como el resto del diente pero comprimido en sentido ántero-posterior. Previamente se ha señalado para Phaenomys la existencia de un fuerte flexo anteromediano y la formación de dos cónulos, el lingual de menor tamaño (cf. Bonvicino et al., 2001:6). La tinción de la dentina revela en el MZUFV 3400 una compleja morfología. Por un lado, se verifica una estructura que podría homologarse con un flexo anteromediano pero que se presenta como una entalladura en la cara anterior del procíngulo, dividida internamente por una proyección de esmalte, con leve desplazamiento lingual. Por el otro, los cónulos se forman a expensas de una gran foseta central, siendo el lingual apenas menor que el labial. Las cúspides principales del diente son subiguales y opuestas; antero y mesolofo se presentan como estructuras filiformes, casi sin exposición de dentina al menos en este estado de desgaste; opuesto al anterolofo se verifica un protolofo como un cordón de esmalte. El mesolofo muestra puntos de contacto con el paracono y con el metacono. Mientras que los flexos linguales son amplios y transversos, los labiales aparecen profundamente recurvados. El posterolofo resulta discernible pero corto y de orientación transversa. El M2 es de contorno cuadrangular, con las cúspides principales apenas alternas y en todo coincidente con la morfología de la porción comparable del M1. El M3 es un diente grande y complejo, con cúspides principales con leve alternancia; de estas, el metacono muestra un pequeño tamaño pero está claramente presente. Destacan en la morfología oclusal de este diente 2 enormes fosetas centrales y subcirculares que corresponden a los fondos de los flexos labiales cerrados por conexiones entre las cúspides principales. El m1 es de contorno subrectangular y patrón de cúspides opuesto; su procíngulo aparece ocupado por una enorme fosétida central que determina 2 conúlidos, el labial mayor que el lingual; un fléxido anteromediano poco penetrante aparece entallado en la cara anterior con leve desplazamiento labial. Mientras que los proto e hipocónido son levente triangulares, meta y entocónidos aparecen como bulbosos. Un anterolófido corto y fusionado al metacónido encierra una fosétida que corresponde al metafléxido. En el lado opuesto aparece un largo protolófido que alcanza el margen dentario. El mesolófido es una estructura de buen desarrollo que hacia su extremo lingual coalesce tanto con el meta como con el entocónido. Sendos proto y ectostílido cierran los fléxidos labiales. Un hipolofúlido de buen desarrollo genera la aparición de una fosétida subcentral. Finalmente, un posterolófido estrecho flanquea el margen posterior del diente. En el m2 se repite el patrón morfológico del m1. Como particularidad -algo inusual en sigmodontinos- proto e hipolofúlidos son más que evidentes. Además, los puntos de coalescencia del mesolófido quedan revelados, en lo que hace al entocónido, por una suerte de entolofúlido doble. El m3 es sigmoide, con cúspides alternas y con menos estructuras que el m1 y m2. Un importante cíngulo anterolabial flanquea el ángulo del diente a la par que sendas fosétidas, correspondientes a los fondos de los meso y posterofléxido, ocupan la mitad lingual. El entocónido, si bien pequeño, es discernible.

Fig. 5. Morfología oclusal de los molares de Phaenomys ferrugineus basada en el ejemplar MZUFV 3400, de izquierda a derecha: serie molar superior y serie molar inferior, ambas izquierdas. Escala = 2 mm.
El postcráneo parcial disponible para el espécimen MZUFV 3400 revela la existencia de 12 vértebras torácicas (e igual número de costillas), 7 lumbares, 4 sacras (3 + 1) y 6 esternebras. El axis presenta una espina neural baja aunque expandida en sentido ántero-posterior, si bien no alcanza a contactar con la espina de la 3ra. cervical. Por su parte, la espina neural de la 2da. torácica está alargada, algo usual en sigmodontinos (Voss, 1988), con una longitud 2 veces y media la de su homóloga de la 3ra. torácica y destacando por una orientación de su porción más distal hacia arriba, en lugar de caudal. Las últimas lumbares presentan los procesos transversos cortos y claramente expandidos en forma de ala y los procesos neurales bajos, anchos y anteriormente inclinados (Fig. 6, material suplementario). En conjunto, la arquitectura para esta última porción axial es consistente con una mayor rigidez y estabilidad de la cintura pélvica, con reducción de los movimientos laterales en detrimento de los dorsoventrales, tal como se verifica en otros mamíferos de vida arborícola (Argot, 2003). Al menos entre las caudales 2da., 3ra. y 4ta. se verifican arcos hemales.
Los resultados obtenidos con cada gen por separado son inconclusivos con relación a la posición filogenética de Phaenomys (Figs. 7 y 8, material suplementario); además de variar en su posición, con la excepción del análisis Bayesiano basado en cit b, todas las relaciones recobradas para Phaenomys tienen poco apoyo. La topología derivada de la matriz de IRBP analizada con MV e IB ubica a Phaenomys como hermano de Juliomys (Boostrap < 50, probabilidad a posteriori = 0.76). La topología obtenida analizando la matriz de cit b mediante análisis de MV ubica a Phaenomys como hermano de Delomys (Bootstrap < 50) y, cuando se analiza con IB, lo recobra como hermano de Wiedomys (probabilidad a posteriori = 1). Por su parte, la matriz combinada analizada con IB muestra, con buen soporte (probabilidad a posteriori = 1), una topología donde Phaenomys es recuperado como hermano de Wiedomys (Fig. 9); la misma relación es recuperada en el análisis MV de esta matriz, aunque con apoyo no significativo (Bootstrap < 50; resultados no mostrados).

Fig. 9. Resultados del análisis filogenético de la matriz combinada de secuencias del gen del citocromo b e IRBP. Árbol de consenso por la mayoría obtenido en el análisis Bayesiano. Los números indican la probabilidad a posteriori (izquierda de la diagonal) y valores de Bootstrap del análisis de máxima verosimilitud (derecha de la diagonal) de los nodos adyacentes. Solo se muestran valores de Bootstrap mayores a 50 %. Un "-" indica que el nodo en cuestión no fue recobrado en el análisis de máxima verosimilitud (ln=-44011.5).
Numerosos autores han destacado similitudes morfológicas -y también algunos han propuesto vinculaciones filogenéticas- entre los géneros Delomys, Juliomys, Phaenomys, Rhipidomys, Thomasomys y Wilfredomys y, con menor extensión, Abrawayaomys, Rhagomys y Wiedomys (e.g., Thomas, 1917; Avila Pires, 1960; Voss, 1993; Bonvicino et al., 2001; Oliveira y Bonvicino, 2002; Pacheco, 2003). Además, los datos moleculares, disponibles para algunos de estos taxones, han permitido elaborar diversas hipótesis filogenéticas parciales (e.g., Smith y Patton, 1999; Oliveira y Bonvicino, 2002; D'Elía et al., 2006b; Salazar-Bravo et al., 2013; Ventura et al., 2013). Bonvicino et al. (2001:10) concluyeron su revisión de Phaenomys indicando que "the systematic position of Phaenomys is nevertheless not clarified by the analysis of morphological information, although karyological data suggest a possible affinity with the forms included in the genus Delomys". Pacheco (2003), en un análisis filogenético basado en caracteres morfológicos con importante muestreo taxonómico, recuperó a Phaenomys como un linaje dentro de un clado correspondiente a la tribu Thomasomyini lato sensu. Este autor sólo pudo estudiar material fragmentario de Phaenomys; esta circunstancia seguramente explica por qué tipificó (Pacheco, 2003:54) como pequeños a los etmoturbinales de Phaenomys cuando, en realidad, muestran una expresión conspicua en la pared lateral del cráneo (Fig. 10, material suplementario; cabe aclarar que este carácter fue discutido y descartado por Weksler [2006:110]).
Las aproximaciones filogenéticas realizadas para los géneros históricamente considerados como parte de Thomasomyini presentan varias concordancias y no pocas diferencias (ver revisión en Pacheco et al., en prensa). Los datos genéticos (cit b + IRBP) con los muestreos taxonómicos más integrales (e.g., Ventura et al., 2013) no recuperan grupos naturales identificables como "tomasominos andinos" (e.g., Rhipidomys, Thomasomys) y "tomasominos atlánticos" (e.g., Delomys, Juliomys, Phaenomys), una hipótesis discutida por algunos autores (e.g., Oliveira y Bonvicino, 2002). Aún así, es remarcable que en análisis basados en IRBP los géneros andinos Aepeomys, Rhipidomys y Thomasomys forman un clado con el ampliamente distribuido -Andes, Amazonia, Selva Atlántica- Rhagomys (véase D'Elía et al., 2006b), mientras que en matrices basadas en secuencias de cit b el género andino Chilomys, para el cual no hay secuencias de IRBP disponibles, forma un clado con Rhipidomys y Thomasomys (Smith y Patton, 1999). Por su parte, una serie de géneros endémicos de la Selva Atlántica e históricamente considerados parte de Thomasomyini ocupan posiciones de grupo hermano -si bien con soporte bajo y sólo en análisis basados en secuencias del gen IRBP- de clados que corresponden a tribus bien diversificadas. Tales son los casos de Abrawayaomys y Akodontini (cf. Ventura et al., 2013), Delomys y Phyllotini (cf. Salazar-Bravo et al., 2013) y Wiedomys y Abrotrichini (cf. Parada et al., 2013).
Las hipótesis filogenéticas generadas en este trabajo (Figs. 7, 8, material suplementario, y Fig. 9) son de interés porque relacionan a Phaenomys con Juliomys (IRBP, MV e IB), o Delomys (cit b, MV) o Wiedomys (cit b, IB; combinado, MV e IB), géneros también endé micos del sudeste de Brasil. Sin embargo, estas hipótesis deben tomarse con cautela dada su incongruencia y debido a que generalmente carecen de apoyo significativo.
La hipótesis de Juliomys y Phaenomys como hermanos es interesante ya que ambos son arborícolas y endémicos de la Selva Atlántica. Si bien Delomys también está restringido a este bioma, se trata de una forma cursorial típica del sotobosque denso (Voss, 1993). Por su parte Wiedomys, género tipo de la tribu Wiedomyini (cf. Reig, 1980), cursorial, es característico en el nordeste de Brasil del Cerrado y la Caatinga (Gonçalves et al., 2005). Diversos autores (e.g., Voss, 1993; Steppan, 1995; Oliveira y Bonvicino, 2002; Pacheco, 2003) han destacado los varios rasgos anatómicos generales que estos géneros comparten. En esta misma situación se presenta Wilfredomys, un género monotípico de los montes riparios y serranos de Uruguay y sudeste de Brasil (Musser y Carleton, 2005), parcialmente alopátrico respecto de Juliomys y Phaenomys. Wilfredomys coincide con estos últimos, además de compartir el nicho arborícola, en varias características anatómicas (cf. Pacheco, 2003). Entre otros rasgos, Phaenomys y Wilfredomys presentan molares superiores con cíngulos linguales bien desarrollados, anterolofos y mesolofos filiformes, crestas supraorbitarias en forma de cordón, sutura coronal en V, jugales grandes, extensión lateral del parietal de gran desarrollo y una fuerte expresión del área de inserción del músculo pterigoideo interno en la mandíbula. Es interesante señalar que Wilfredomys y Wiedomys se recobran como géneros hermanos en la filogenia de Pacheco (2003) y que algunos autores (R. Voss, com. pers.; J. Salazar-Bravo, com. pers.) consideran -sobre la base de su similitud morfológica- que podrían ser congenéricos (véase además Pine, 1980).
La inclusión de Wilfredomys en análisis basados en secuencias de ADN es necesaria para explorar si la posición del resto de los elementos "atlánticos" tradicionalmente asociados a Thomasomyini (cf. Voss, 1993; Steppan, 1995; Pacheco, 2003), en particular Abrawayaomys, Juliomys y Phaenomys, se estabiliza en las inferencias filogenéticas de Sigmodontinae. También resulta crucial el análisis de un mayor número de genes (Lessa et al., 2014). Estos estudios indicarán si es pertinente expandir (y, entonces, rediagnosticar) la tribu Wiedomyini para incluir a alguno de estos géneros, a la par que contribuirán a detallar el papel de la región sudoriental del subcontinente en la diversificación de los sigmodontinos (Smith y Patton, 1999; Pardiñas et al., 2014).
AGRADECIMIENTOS
A los biólogos Mario Ribeiro Moura y Rodolfo Stumpp por la captura e identificación a campo del MZUFV 3400. Este trabajo se ha nutrido de años de intercambios con numerosos colegas, entre otros Jorge Salazar-Bravo, Carlos Galliari, Robert Voss, a quienes agradecemos profundamente. Del mismo modo, a los curadores David Flores y Enrique González, que facilitaron o donaron materiales comparativos, y a este último por la lectura crítica del manuscrito. Los estudios morfológicos se hicieron con equipo adquirido en el marco del PICT 2008-547 (a UFJP), mientras que las secuencias de ADN y análisis filogenéticos se realizaron con el apoyo financiero de FONDECYT 1141055 (a GD).
LITERATURA CITADA
1. ARGOT C. 2003. Functional-adaptative anatomy of the axial skeleton of some extant marsupials and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. Journal of Morphology 255:279-300. [ Links ]
2. AVILA-PIRES F. 1960. Um novo gênero de roedor sul-americano. Boletim do Museu Nacional, Nóva Serie, Zoologia 220:1-6. [ Links ]
3. BONVICINO CR, JA OLIVEIRA, PC D'ANDREA y RW CARVALHO. 2001. The endemic Atlantic forest rodent Phaenomys ferrugineus (Thomas, 1894) (Sigmodontinae): New data on its morphology and karyology. Boletim do Museu Nacional 467:1-12. [ Links ]
4. CARLETON MD. 1973. A survey of gross stomach morphology in New World Cricetinae (Rodentia, Muroidea), with comments on functional interpretations. Miscellaneous Publications, Museum of Zoology, University of Michigan 146:1-43. [ Links ]
5. D'ELÍA G. En prensa. Sigmodontinae incertae sedis. En: Mammals of South America. Volume 2, Rodents (JL Patton, UFJ Pardiñas y G D'Elía, eds.). The University of Chicago Press, Chicago, Illinois. [ Links ]
6. D'ELÍA G, AA OJEDA, F MONDACA y MH GALLARDO. 2006a. New data of the long-clawed mouse Pearsonomys annectens (Cricetidae, Sigmodontinae) and additional comments on the distinctiveness of Pearsonomys. Mammalian Biology 71:39-51. [ Links ]
7. D'ELÍA G, L LUNA, EM GONZÁLEZ y BD PATTERSON. 2006b. On the Sigmodontinae radiation (Rodentia, Cricetidae): An appraisal of the phylogenetic position of Rhagomys. Molecular Phylogenetics and Evolution 38:558-564. [ Links ]
8. D'ELÍA G, UFJ PARDIÑAS, P TETA y JL PATTON. 2007. Definition and diagnosis of a new tribe of sigmodontine rodents (Cricetidae: Sigmodontinae), and a revised classification of the subfamily. Gayana 71:187-194. [ Links ]
9. GONÇALVES PR, F ALMEIDA y CR BONVICINO. 2005. A new species of Wiedomys (Rodentia:Sigmodontinae) from Brazilian Cerrado. Mammalian Biology 70:46-60. [ Links ]
10. LESSA EP, JA COOK, G D'ELÍA y JC OPAZO. 2014. Rodent diversity in South America: Transitioning into the genomics era. Fronteirs in Ecology and Evolution 2:39. DOI: 10.3389/fevo.2014.00039. [ Links ]
11. MUSSER GM y MD CARLETON. 2005. Superfamily Muroidea. Pp. 894-1531 en Mammal species of the world: A taxonomic and geographic reference (DE Wilson and DM Reeder, eds.). 3rd ed, Johns Hopkins University Press, Baltimore, Maryland. [ Links ]
12. OLIVEIRA JA de y CR BONVICINO. 2002. A new species of sigmodontine rodent from the Atlantic forest of eastern Brazil. Acta Theriologica 47:307-322. [ Links ]
13. PACHECO V. 2003. Phylogenetic analyses of the Thomasomyini (Muroidea: Sigmodontinae) based on morphological data. Tesis doctoral, City University of New York, EE. UU. [ Links ]
14. PACHECO V, JL PATTON y G D'ELÍA. En prensa. Tribe Thomasomyini Steadman and Ray, 1982. En: Mammals of South America. Volume 2, Rodents (JL Patton, UFJ Pardiñas y G D'Elía, eds.). The University of Chicago Press, Chicago, Illinois. [ Links ]
15. PARADA A, UFJ PARDIÑAS, J SALAZAR-BRAVO, G D'ELÍA y E PALMA. 2013. Dating an impressive Neotropical radiation: Molecular time estimates for the Sigmodontinae (Rodentia) provide insights into its historical biogeography. Molecular Phylogenetics and Evolution 66:960-968. [ Links ]
16. PARDIÑAS UFJ, G D'ELÍA y CIRIGNOLI S. 2003. The genus Akodon (Muroidea: Sigmodontinae) in Misiones, Argentina. Mammalian Biology 68:129-143. [ Links ]
17. PARDIÑAS UFJ, G LESSA, P TETA, J SALAZAR-BRAVO y EM CÂMARA. 2014. A new genus of sigmodontine rodent from Eastern Brazil and the origin of the tribe Phyllotini. Journal of Mammalogy 95:201-215. [ Links ]
18. PARDIÑAS UFJ, P TETA, G D'ELÍA y C GALLIARI. 2008. Rediscovery of Juliomys pictipes (Rodentia: Cricetidae) in Argentina: Emended diagnosis, geographic distribution, and insights on genetic structure. Zootaxa 1758:29-44. [ Links ]
19. PASSAMANI M, RA SERPA CERBONCINI y J EVERALDO DE OLIVEIRA. 2011. Distribution extension of Phaenomys ferrugineus (Thomas, 1894), and new data on Abrawayaomys ruschii Cunha and Cruz, 1979 and Rhagomys rufescens (Thomas, 1886), three rare species of rodents (Rodentia: Cricetidae) in Minas Gerais, Brazil. Check List, Journal of Species Lists and Distribution 7:827-831. [ Links ]
20. PINE RH. 1980. Notes on rodents of the genera Wiedomys and Thomasomys (including Wilfredomys). Mammalia 44:195-202. [ Links ]
21. REIG OA. 1977. A proposed unified nomenclature for the enamelled components of the molar teeth of the Cricetidae (Rodentia). Journal of Zoology (London) 181:227-241. [ Links ]
22. REIG OA. 1980. A new fossil genus of South American cricetid rodents allied to Wiedomys, with an assessment of the Sigmodontinae. Journal of Zoology (London) 192:257-281. [ Links ]
23. SALAZAR-BRAVO J, UFJ PARDIÑAS y G D'ELÍA. 2013. A phylogenetic appraisal of Sigmodontinae (Rodentia, Cricetidae) with emphasis on phyllotine genera: Systematics and biogeography. Zoologica Scripta 42:250-261. [ Links ]
24. SMITH MF y JL PATTON. 1999. Phylogenetic relationships and the radiation of sigmodontine rodents in South America: Evidence from cytochrome b. Journal of Mammalian Evolution 6:89-128. [ Links ]
25. STEPPAN SJ. 1995. Revision of the tribe Phyllotini (Rodentia: Sigmodontinae), with a phylogenetic hypothesis for the Sigmodontinae. Fieldiana, Zoology, n.s. 80:1-112. [ Links ]
26. THOMAS O. 1894. Description of some new Neotropical Muridae. Annals and Magazine of Natural History (London) 6(14):345-366. [ Links ]
27. THOMAS O. 1917. On the arrangement of the South American rats allied to Oryzomys and Rhipidomys. Annals and Magazine of Natural History (London) 8(20):192-198. [ Links ]
28. THOMPSON JD, TJ GIBSON, F PLEWNIAK, F JEANMOUGIN y HG HIGGINS. 1997. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 24:4876-4882. [ Links ]
29. VENTURA K, MJ SILVA, L GEISE, Y LEITE, UFJ PARDIÑAS, Y YONENAGA-YASSUDA y G D'ELÍA. 2013. The phylogenetic position of the enigmatic Atlantic forest-endemic spiny mouse Abrawayaomys (Rodentia: Sigmodontinae). Zoological Studies 52:55 doi:10.1186/1810-522X-52-55. [ Links ]
30. VOSS RS. 1988. Systematics and ecology of Ichtyomtine rodents (Muroidea): Patterns of morphological evolution in small adaptive radiation. Bulletin of the American Museum of Natural History 188:262-493. [ Links ]
31. VOSS RS. 1993. A revision of the Brazilian muroid rodent genus Delomys with remarks on "Thomasomyine" characters. American Museum Novitates 3073:1-44. [ Links ]
32. WEKSLER M. 2006. Phylogenetic relationships of oryzomine rodents (Muroidea: Sigmodontinae): Separate and combined analyses of morphological and molecular data. Bulletin of the American Museum of Natural History 296:1-149. [ Links ]
Los ejemplares estudiados en esta contribución pertenecen a las siguientes colecciones: Colección de Mamíferos de la Universidade Federal de Lavras (CMUFLA; Minas Gerais, Brasil), Colección de Mamíferos del Centro Nacional Patagónico (CNP; Puerto Madryn, Chubut, Argentina), Colección de Mamíferos del Museu de Zoologia, Departamento de Biologia Animal, Universidade Federal de Viçosa (MZUFV; Minas Gerais, Brasil), Colección Nacional de Mastozoología, Museo Argentino de Ciencias Naturales "B. Rivadavia" (MACN; Buenos Aires, Argentina).
Delomys dorsalis (n = 3). Argentina: Misiones, Reserva Vida Silvestre Urugua-í (MACN 22253-22255).
Juliomys pictipes (n = 3). Argentina: Misiones, Arroyo de Salamanca (CNP 895), Sendero Macuco (MACN 23385); Brazil: Minas Gerais, Itamarati de Minas (MZUFV 2762).
Phaenomys ferrugineus (n = 2). Brasil: Minas Gerais, Itamonte, Hotel Casa Alpina (CMUFLA 904), Parque Estadual da Serra do Brigadeiro (MZUFV 3400).
Rhipidomys austrinus (n = 1). Argentina: Salta, Orán, Santa María (MACN 51.158).
Thomasomys baeops (n = 1). Ecuador (MACN 3.20).
Thomasomys cinereus (n = 1). Ecuador (MACN 31.155).
Thomasomys paramorum (n = 1). Ecuador (MACN 31.147).
Wiedomys pyrrhorhinos (n = 2). Brasil: Bahía (CNP 3643, CNP 4278).
Wilfredomys oenax (n = 2). Uruguay: Florida, Cerro Colorado (CNP 2378, CNP 2379).
Fig. 2. Morfología externa dorsal (arriba) y ventral (abajo) de Phaenomys ferrugineus basada en la piel disecada del ejemplar MZUFV 3400.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup1.pdf
Fig. 3. Morfología general del cráneo y mandíbula de Phaenomys ferrugineus basada en el ejemplar MZUFV 3400.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup2.pdf
Fig. 6. Región lumbo-sacra y pélvica (en vista dorsal) de Phaenomys ferrugineus basada en el ejemplar MZUFV 3400.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup3.pdf
Fig. 7. Resultados del análisis filogenético de secuencias del gen IRBP. Árbol de consenso por la mayoría obtenido en el análisis Bayesiano.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup4.pdf
Fig. 8. Resultados del análisis filogenético de secuencias del gen del citocromo b. Árbol de consenso por la mayoría obtenido en el análisis Bayesiano.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup5.pdf
Fig. 10. Vista lateral de la región orbitaria izquierda en Phaenomys ferrugineus (MZUFV 3400) donde se aprecia el desarrollo del etmoturbinal (e). Abreviaturas: ef = foramen etmoide; max = maxilar; of: foramen óptico; os = órbitoesfenoides; pal = palatino; spl = foramen esfenopalatino.
http://www.sarem.org.ar/wp-content/uploads/2014/12/SAREM_MastNeotrop_21-2_Pardinas-sup6.pdf

