










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383
Mastozool. neotrop. vol.21 no.2 Mendoza dic. 2014
NOTA
Hábitos alimentarios del puma (Puma concolor) e incidencia de la depredación en la mortandad de guanacos (Lama guanicoe) en el noreste de la Patagonia
Cynthia S. Fernández1 y Ricardo Baldi2,3
1 Facultad de Ciencias Naturales, Universidad Nacional de la Patagonia San Juan Bosco, Boulevard Brown s/n, 9120 Puerto Madryn, Chubut, Argentina.
2 Centro Nacional Patagónico (CENPAT-CONICET), Boulevard Brown 2915, 9120 Puerto Madryn, Chubut, Argentina [correspondencia: <rbaldi@cenpat-conicet.gob.ar>].
3 Patagonian and Andean Steppe Program, Wildlife Conservation Society, Argentina.
Recibido 10 julio 2014.
Aceptado 13 noviembre 2014.
Editor asociado: J Pereira
RESUMEN.
En la Patagonia, las relaciones funcionales entre el puma y sus presas nativas se han modificado como consecuencia de la introducción de especies exóticas. Estudiamos los hábitos alimentarios de Puma concolor en el nordeste del Chubut durante un período de 3 años y encontramos que: (1) el guanaco, Lama guanicoe, fue la presa más importante en su dieta; (2) ante una reducción sustancial en la abundancia de ovinos, el puma respondió con una ampliación de su nicho trófico mediante la incorporación de presas de menor tamaño; y (3) la depredación por pumas fue la principal causa de muerte de guanacos.
ABSTRACT.
Feeding habits of the puma (Puma concolor) and the incidence of predation on guanaco (Lama guanicoe) mortality in northeastern Patagonia.
In Patagonia, the functional relationships between pumas and their native prey species have been modified as a result of the introduction of exotics. We studied the feeding habits of Puma concolor in northeastern Chubut province during 3 years and found that (1) the native guanaco, Lama guanicoe, was the most important prey in the diet of pumas; (2) pumas expanded their trophic niche by incorporating smaller prey species to their diets after a substantial reduction in the abundance of the introduced sheep; and (3) puma predation was the main cause of guanaco mortality.
Palabras clave: Abundancia de presas; Argentina; Hábitos alimentarios; Mortalidad de guanacos; Predador-presa.
Key words: Argentina; Feeding habits; Guanaco mortality; Predator-prey; Prey abundance.
El puma, Puma concolor, presenta el mayor rango de distribución entre los mamíferos terrestres silvestres en el hemisferio occidental (Logan y Sweanor, 2001), lo cual lo expone a una gran variedad de ensambles de presas desde el oeste de Canadá hasta el sur de Argentina y Chile. El tamaño corporal del puma se correlaciona positivamente con la latitud y con el tamaño promedio de las presas capturadas (Iriarte et al., 1990). Los pumas más pequeños, que habitan regiones ecuatoriales, consumen presas más pequeñas -de peso menor a los 15 kg- y taxonómicamente diversas (Taber et al., 1997) que se encuentran en mayor disponibilidad, lo cual les permite además reducir la competencia interespecífica con el jaguar (Panthera onca; Iriarte et al., 1990; Iriarte et al., 1991). Por el contrario, las presas de mayor tamaño son más abundantes en su dieta a medida que aumenta la latitud, siendo los ungulados los ítems principales hacia las regiones templadas de América del Norte y América del Sur (Logan y Sweanor, 2001).
El puma es el principal predador tope terrestre en la Patagonia y sus presas nativas incluyen varias especies por sobre los 15 kg de peso, como el guanaco (Lama guanicoe), el choique (Rhea pennata) y el huemul (Hippocamelus bisulcus; Courtin et al., 1980). Sin embargo, sus relaciones funcionales han sido modificadas por la introducción de especies exóticas (domésticas y silvestres), la degradación del hábitat y la caza de herbívoros nativos, entre otros factores (Ballard et al., 2001; Rau y Jiménez, 2002; Novaro y Walker, 2005). Novaro et al. (2000) han propuesto que varias especies nativas se encuentran ecológicamente extintas en diversos sitios del noroeste de la Patagonia; esto se verifica en particular para el guanaco, que representa < 1% de la dieta de los carnívoros nativos en esos sitios, los cuales se alimentan en su lugar principalmente de presas introducidas. Adicionalmente, la depredación por pumas sobre algunas poblaciones de guanacos que ocurren a densidades muy bajas podría limitar su crecimiento e incluso provocar su erradicación local o regional (Novaro y Walker, 2005).
En este trabajo examinamos los hábitos alimentarios del puma durante un período de 3 años en una localidad del nordeste de la provincia del Chubut, Argentina, y evaluamos en particular la selección de guanacos y ovinos según su abundancia, que para el caso de los ovinos se redujo -por decisiones de manejo- en un 30% durante el período cubierto por este estudio. También evaluamos la importancia de la depredación por pumas como causa de muerte del guanaco. La hipótesis general de este trabajo propone que, en ausencia de competidores, la selección de presas por el puma está influenciada por el tamaño y la abundancia de las mismas (Gitlleman y Harvey, 1982; Carbone et al., 1999). De ser así, esperamos que: (1) guanacos y ovinos sean las presas más importantes en la dieta del puma; (2) ambas especies estén representadas de acuerdo a su abundancia relativa en el área de estudio; y (3) la disminución del número de ovinos resulte en que las especies más pequeñas incrementen su representatividad en la dieta del puma.
Este estudio se llevó a cabo en el Refugio de Vida Silvestre "La Esperanza" (42°11'S, 64°57'W), en el nordeste de la provincia del Chubut, Argentina, entre los años 2002 y 2004. Este establecimiento de 6700 ha pertenece, desde el año 2000, a la Organización No Gubernamental Fundación Patagonia Natural y cuenta desde esa fecha con un guardafauna permanente. La carga ovina se redujo por decisiones de manejo, de 651 a 455 cabezas durante el período de estudio, obteniéndose su número total anual durante cada evento de esquila. La abundancia de guanacos en el área fue estimada por Baldi (2004) utilizando el método Distance Sampling (Buckland et al., 1993), permaneciendo relativamente estable entre 2002 (5.92 guanacos.km-2; %CV = 14.20) y 2004 (5.30 guanacos.km-2; %CV = 14.18). Durante los relevamientos se estimó, en base al tamaño corporal, que el 66.9% de 329 guanacos observados fueron adultos, el 19.8% chulengos (crías < 1 año de edad) y el 13.3% juveniles de entre 1 y 2 años de edad (Baldi, 2004).
La vegetación en el área de estudio pertenece a la Provincia Fitogeográfica del Monte, influenciada por la Península Valdés con comunidades de la Provincia Patagónica (León et al., 1998). La fisonomía dominante es la de estepa arbustiva, con especies como jarilla (Larrea divaricata y Larrea nitida), quilembay (Chuquiraga avellanedae) y halófitas como zampa (Atriplex lampa), mata azul (Cyclolepis genistoides) y jume (Suaeda divaricata). La estructura de la vegetación ofrece sitios que favorecen el acecho del puma a sus presas, según lo descrito en estudios para este (Marino y Baldi, 2008) y otros sitios (Franklin et al., 1999; Bank et al., 2002).
Se relevó el área con una frecuencia quincenal, recolectándose todas las fecas de puma encontradas en los alrededores de senderos que totalizan 46 km y varios cañadones. Se registró la ocurrencia de cadáveres de guanacos y ovinos, determinándose cuando fue posible la causa de muerte; para el guanaco también se verificó el sexo por observación de los genitales o bien por el dimorfismo en los caninos y la categoría de edad de acuerdo al reemplazo y desgaste dentario (Raedeke, 1979).
Las fecas obtenidas se esterilizaron en estufa a 90°C durante 48 horas y luego se lavaron para separar restos identificables de animales. Los restos óseos y dentarios presentaron un alto grado de fragmentación, por lo tanto la determinación de las presas se basó principalmente en la identificación de los pelos de mamíferos, ya que la estructura microscópica de los mismos se mantiene inalterada aun luego de ser digeridos (Chehébar y Martín, 1989). Los restos identificables fueron comparados macroscópica y/o microscópicamente con material de referencia del Laboratorio de Fauna Silvestre (Unidad de Ecología Terrestre, CENPAT), utilizando un microscopio óptico con oculares de 40x y 100x.
Se definió como ítem-presa a cada tipo de presa diferente identificado en la materia fecal del puma. El material de cada muestra se distribuyó en una grilla de 18 celdas y se analizaron celdas al azar hasta que no se reconocieran nuevos ítems-presa. Se calculó el porcentaje de ocurrencia como el número de veces que un ítem-presa particular fue encontrado en las fecas, sobre el número total de ítems-presa ×100. Se calculó la frecuencia de ocurrencia como el número de fecas en que un ítem es determinado sobre el número total de fecas, para así estimar la biomasa relativa consumida aplicando la corrección de Ackerman et al. (1984); esta última da cuenta de que las presas pequeñas poseen más pelo por unidad de peso corporal consumido y, por lo tanto, pueden ser sobreestimadas en las dietas de los carnívoros (Floyd et al., 1978). La ecuación se expresa como: Y = 1.980+0.035*X, donde Y = kg de presa consumida por feca colectada, y X = peso promedio de un individuo adulto de cada presa (los pesos fueron tomados de Iriarte et al., 1990 y Redford y Eisemberg, 1992). Para evaluar el carácter generalista o especialista de la dieta del puma se calculó la amplitud de nicho trófico mediante la expresión de Levins (1968): B = 1/ Σ pi2 donde pi es la proporción relativa de presas de la categoría i en la dieta. Para caracterizar la selección de presas según su tamaño corporal, se calculó el peso promedio de las presas de vertebrados consumidas por el puma, sumando los productos de las frecuencias de cada tipo de presa por el logaritmo natural de su peso y dividiéndolo luego por el número total de tipos de presa en la muestra (Iriarte et al., 1990). Para evaluar las diferencias temporales en la dieta, las especies de mamíferos determinadas como presa fueron clasificadas en 4 grupos de acuerdo a su tamaño corporal: guanacos, ovinos, presas medianas (aquellas con un peso promedio entre 1 y 15 kg) y presas pequeñas (aquellas con un peso promedio < 1 kg). Para evaluar la preferencia del puma por guanacos u ovinos se utilizaron 2 aproximaciones. En primer lugar la frecuencia de ocurrencia de guanacos y ovinos en las fecas de puma fue comparada con las frecuencias esperadas derivadas de las estimaciones de abundancia mediante una prueba de chi cuadrado (X2) y bajo la hipótesis de un depredador no selectivo (Manly et al., 1972). El estimador usado para calcular las frecuencias de presas esperadas en las fecas fue: πi = diλi/Σi diλi (Link y Karanth, 1994), en donde la presa i tiene una densidad de población di, y λi (derivado de la ecuación de Ackerman et al., 1984) es el número de fecas producidas por cada animal consumido de la especie i. Las frecuencias esperadas de las especies-presa en las fecas del puma se obtuvieron utilizando del software Scatman (Link y Karanth, 1994). Complementariamente, el número de ovinos y guanacos encontrados muertos en el campo como consecuencia de la depredación por pumas fue comparado con los valores esperados mediante una prueba de bondad de ajuste de X2. Las frecuencias esperadas se calcularon a partir de las proporciones relativas de guanacos y ovinos en la población total de ungulados, bajo la hipótesis de un depredador no selectivo. Finalmente, se registró la incidencia de la depredación como causa de mortalidad de guanacos y la posible selección de categorías etarias (adultos, chulengos y juveniles) por parte del puma, comparándose mediante pruebas de bondad de ajuste de X2 el número de muertes observadas para cada categoría con los valores esperados calculados a partir de las proporciones de la población de acuerdo a los censos realizados en el área de estudio.
Se determinaron taxonómicamente 228 restos provenientes de 71 fecas. El 96.5% correspondieron a mamíferos, clasificados en 18 ítems-presa y el 3.5% restante fueron aves que se agruparon en una única categoría (Tabla 1). Guanacos y ovinos (ungulados) fueron las presas más importantes en términos de biomasa, en particular el guanaco que constituyó más de la mitad de la biomasa total consumida. En cambio, las presas medianas fueron las más frecuentemente encontradas en las fecas (36.7% para todo el período de estudio), mientras que los ungulados presentaron en total una frecuencia de ocurrencia similar a la de las presas pequeñas (ca. 30%, Tabla 1).
Tabla 1. Composición dietaria anual y total del puma por ítem y categoría de presa expresada en porcentaje de ocurrencia (% Oc.) y de biomasa (% Bi.) y número de presas, peso, amplitud del nicho trófico (ANT) y promedio de ítems presa por feca (N prom. I-P por feca). Otras abreviaturas: n = número de fecas de puma recolectadas, nd = no disponible, *especie introducida).

La dieta del puma estuvo dominada por la presencia de 14 ítems-presa nativos que constituyeron el 76.6% de la biomasa total consumida y se registraron 3 especies introducidas. De estas últimas, la oveja y la liebre europea fueron las más frecuentes (18% de los ítems-presa) y aportaron un 23.1% de la biomasa relativa consumida.
Durante el período de estudio, la abundancia de guanacos se mantuvo relativamente estable (Fig. 1). La disminución en el número de ovinos se asoció a un incremento en el número de ítems-presa consumidos por el puma (Fig. 1). Sin embargo, el guanaco continuó siendo la presa más importante durante los 3 años de estudio, tanto en ocurrencia como en biomasa (Tabla 1). Los ovinos, en cambio, disminuyeron su importancia en dos tercios tanto en porcentaje de ocurrencia como en biomasa relativa en el año 2004, en comparación al año 2002 (Tabla 1). Consecuentemente, la amplitud de nicho trófico aumentó y disminuyó el peso promedio de las presas consumidas por el puma, ya que las presas medianas, en particular el peludo (Chaetophractus villosus), aumentaron su ocurrencia y biomasa en la dieta hacia el año 2004 (Tabla 1). Además, se identificaron presas no registradas durante 2002 y 2003, entre ellas una especie de marsupial y una de quiróptero (Tabla 1).
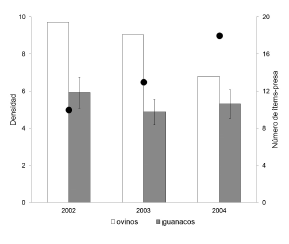
Fig. 1. Densidad anual de guanacos (individuos.km-2 ± E.E.) y ovinos en el el Refugio de Vida Silvestre "La Esperanza", nordeste del Chubut, Argentina (columnas) y número de ítems-presa en la dieta del puma (círculos llenos). La densidad de ovinos se calculó en base al conteo total anual durante la esquila, dividido por la superficie del área de estudio.
Si bien el número de fecas recolectadas por año aumentó, también se incrementó el número promedio de ítems-presa encontrados por feca en 2004 (Tabla 1). Solo el 16% de las fecas recolectadas durante 2002 y 2003 presentaron > 3 ítems-presa; en contraste, más del 55% de las fecas encontradas durante 2004 presentaron > 3 ítems-presa, registrándose muestras con hasta 6 y ninguna con < 2 ítems-presa.
La frecuencia de guanacos y ovinos representados en las fecas del puma (año 2002 = 10 guanacos, 8 ovinos; año 2003 = 10 guanacos, 10 ovinos; año 2004 = 23 guanacos, 7 ovinos) y lo esperado de acuerdo a sus abundancias relativas (año 2002 = 8 guanacos, 10 ovinos; año 2003 = 8 guanacos, 12 ovinos; año 2004 = 21 guanacos, 9 ovinos) no difirieron significativamente en ninguno de los años considerados (2002: X2 = 0.90, p = 0.343; 2003: X2 = 0.83, p = 0.361; 2004: X2 = 0.50, p = 0.480; g.l. = 1, en todos los casos). Asimismo, el número de carcasas de individuos de ambas especies muertos por puma (año 2002 = 8 guanacos, 11 ovinos; año 2003 = 10 guanacos, 25 ovinos; año 2004 = 14 guanacos, 14 ovinos) y lo esperado de acuerdo a su abundancia relativa en el campo (año 2002 = 7 guanacos, 12 ovinos; año 2003 = 12 guanacos, 23 ovinos; año 2004 = 12 guanacos, 16 ovinos) no difirieron significativamente en ninguno de los años considerados (2002: X2 = 0.14, p = 0.705; 2003: X2 = 0.61, p = 0.435; 2004: X2 = 0.42, p = 0.517; g.l. = 1, en todos los casos). Ambas líneas de evidencia sugieren que guanacos y ovinos fueron depredados por el puma de acuerdo a su abundancia.
La depredación por pumas fue la principal causa de muerte de guanacos (50.8% de 63 carcasas examinadas), seguida por los enganches en alambrados (14.3%), caza ilegal desde campos linderos (4.8%), inanición de chulengos separados de sus madres (6.3%) y heridas punzantes en la cabeza y cuello de un macho adulto, compatibles con agresiones intraespecíficas (1.6%); no pudo determinarse la causa de muerte en el 22.2% restante. De los 32 guanacos muertos por pumas, 15 fueron adultos (53.3% hembras, 33.3% machos, 13.3% no identificados), 15 chulengos (20% hembras, 40% machos, 40% no determinado) y 2 juveniles de entre 1 y 2 años de edad. El número de guanacos depredados según clases de edad fue diferente a los valores esperados (X2 = 17.01, g.l. = 2, p = 0.0002). Los chulengos fueron seleccionados positivamente (X2 = 16.61, g.l. = 1, p = 0.0001), los adultos fueron evitados (X2 = 4.98, g.l. = 1, p = 0.025) y los juveniles fueron depredados de acuerdo a lo esperado (X2 = 2.13, g.l. = 1, p = 0.144).
El guanaco fue la especie predominante en la dieta del puma, tanto en su frecuencia de representatividad fue similar en los diferentes años. En cambio, la reducción en el número de ovinos, segunda presa en importancia en términos de biomasa, se reflejó en la disminución de esta especie en la dieta del puma. Además, la frecuencia de ocurrencia de algunas presas medianas aumentó y otras especies no registradas fueron incorporadas a la dieta, la cual fue más diversa al disminuir la importancia de los ovinos. Por lo tanto, los principales resultados apoyan las hipótesis planteadas sobre el predomino de presas de tamaño grande en la dieta del puma en la región, sus hábitos alimentarios en relación con la abundancia relativa de ungulados y el cambio de estrategia de alimentación al disminuir la abundancia de alguna de sus presas principales.
Existe evidencia de la preferencia por parte del puma de especies presa de tamaño grande en distintos sitios (Iriarte et al., 1991; Polisar et al., 2003; Murphy y Ruth, 2010), como así también del empleo de presas alternativas cuando las presas preferidas son escasas (Iriarte et al., 1990; Wolff, 2001; Novack, 2003). Por ejemplo, en el sur de Chile, los pumas consumieron más liebres europeas cuando las densidades de guanacos fueron bajas. En cambio, cuando las poblaciones de guanacos se incrementaron, tambien aumentaron su representación en la dieta del puma (Yáñez et al., 1986; Iriarte et al., 1990). Fryxell y Lundberg (1994) han demostrado que un depredador captura presas de menor calidad sólo cuando se ha reducido la disponibilidad de presas energéticamente más rentables. El aumento en la amplitud del nicho trófico observado en este estudio estaría asociado a una disminución numérica de una de las presas principales y sugiere que la selección por parte del puma está influenciada por la abundancia de las presas más grandes y energéticamente más rentables, pero maximizando el número de individuos consumidos ante una disminución de presas de mayor tamaño.
Si bien el aporte de los ovinos a la dieta del puma fue importante, esta estuvo mayormente compuesta por especies nativas, en coincidencia con lo encontrado en sitios de Santa Cruz (Zanón Martínez et al., 2012) y en contraste con sitios de Neuquén y el oeste de Río Negro, donde el guanaco fue considerado ecológicamente extinto con respecto a su rol como presa, representando < 1% de la biomasa consumida (Novaro et al., 2000). El ganado ovino, el ciervo colorado y la liebre europea han desplazado a las presas nativas en diferentes áreas de la Patagonia, a la vez que subsidiaron a los depredadores nativos como el puma (Novaro y Walker, 2005; Zanón Martínez et al., 2012). En nuestra área de estudio, la depredación por pumas fue la principal causa de mortalidad de guanacos. Los chulengos fueron fuertemente seleccionados, en coincidencia con otros estudios que describen la relación entre pumas y guanacos (Iriarte et al., 1991; Franklin et al., 1999). Nuestro estudio muestra que el puma se comportó como un depredador oportunista, cuyos hábitos alimentarios fueron flexibles en el tiempo. El guanaco es ecológicamente funcional como presa y el ensamble de presas nativas cobra mayor importancia al disminuir la abundancia de ganado ovino. La Patagonia árida es una región extensa donde el impacto de actividades humanas, como la ganadería, ha sido históricamente significativo, manifestándose sobre las poblaciones silvestres y sus interacciones ecológicas. Los estudios de las estrategias de alimentación de los carnívoros en función de factores temporales y espaciales contribuyen a la comprensión de las relaciones funcionales entre los depredadores y sus presas nativas, resultando fundamentales para la planificación de la conservación de procesos ecológicos en la región.
Agradecimientos.
A la Fundación Patagonia Natural por permitirnos trabajar en el Refugio de Vida Silvestre "La Esperanza". A Victoria Rodríguez, Gustavo Zamora, Andrea Marino, Sebastián Cévoli, Kerry Lock y Romina D'Agostino por la asistencia en el campo. A Ulyses Pardiñas y Alicia Toyos (CENPAT - CONICET) por la ayuda en la determinación taxonómica de los restos de presas y a 2 revisores anónimos por sus comentarios y sugerencias. Este trabajo fue financiado por Wildlife Conservation Society.
LITERATURA CITADA
1. ACKERMAN BB, FG LINDZEY y TP HEMKER. 1984. Cougar food habits in southern Utah. Journal of Wildlife Management 48:147-155. [ Links ]
2. BALDI R. 2004. Procesos ecológicos que afectan la conservación de especies-clave en la Patagonia árida. Investigación en el Establecimiento La Esperanza. Informe Técnico presentado a la Fundación Patagonia Natural, Wildlife Conservation Society, Argentina. [ Links ]
3. BALLARD WB, D LUTZ, TW KEEGAN, LH CARPENTER y JR DEVOS. 2001. Deer-predator relationships: A review of recent North American studies with emphasis on mule and black-tailed deer. Wildlife Society Bulletin 29:99-115. [ Links ]
4. BANK MS, RJ SARNO, NK CAMPBELL y WL FRANKLIN. 2002. Predation on guanacos Lama guanicoe by southernmost mountain lions Puma concolor during a historically severe winter in Torres del Paine National Park, Chile. Journal of Zoology (London) 258:215-222. [ Links ]
5. BUCKLAND ST, DR ANDERSON, KP BURNHAM y JL LAAKE. 1993. Distance Sampling: Estimating abundance of biological populations. Chapman & Hall, London. [ Links ]
6. CARBONE C, GM MACE, SC ROBERTS y DW MACDONALD. 1999. Energetic constraints on the diet of terrestrial carnivores. Nature 402:286-288. [ Links ]
7. CHEHÉBAR C y S MARTÍN. 1989. Guía para el reconocimiento microscópico de los pelos de los mamíferos de la Patagonia. Doñana, Acta Vertebrata 16:247-291. [ Links ]
8. COURTIN SL, NV PACHECO y WD ELDRIDGE. 1980. Observaciones de alimentación, movimientos y preferencias de hábitat del puma en el Islote Rupanco. Medio ambiente 4:50-55. [ Links ]
9. FLOYD TJ, LD MECH y PJ JORDAN. 1978. Relating wolf scat contents to prey consumed. Journal of Wildlife Management 42:528-532. [ Links ]
10. FRANKLIN WL, WE JOHNSON, RJ SARNO y JA IRIARTE. 1999. Ecology of the Patagonian puma Felis concolor patagonica in southern Chile. Biological Conservation 90:33-40. [ Links ]
11. FRYXELL JM y P LUNDBERG. 1994. Diet choice and predator-prey dynamics. Evolutionary Ecology 8:407-421. [ Links ]
12. GITLLEMAN JL y PH HARVEY. 1982. Carnivore home range size, metabolic needs and ecology. Behavioral Ecology and Sociobiology 10:57-63. [ Links ]
13. IRIARTE JA, WL FRANKLIN, WE JOHNSON y KH REDFORD. 1990. Biogeographic variation of food habits and body size of the American puma. Oecologia 85:185-190 [ Links ]
14. IRIARTE JA, WE JOHNSON y WL FRANKLIN. 1991. Feeding ecology of the Patagonian puma in southernmost Chile. Revista Chilena de Historia Natural 64:145-156. [ Links ]
15. LEÓN RJC, D BRAN, M COLLANTES, JM PARUELO y A SORIANO. 1998. Grandes unidades de vegetación de la Patagonia extra andina. Ecología Austral 8:125-144. [ Links ]
16. LEVINS R. 1968. Evolution in changing environments. Princeton University Press, Princeton, NJ. [ Links ]
17. LINK WA y KU KARANTH .1994. Correcting for overdispersion in test of prey selectivity. Ecology 75:2456-2459. [ Links ]
18. LOGAN KA y LL SWEANOR. 2001. Desert puma. Evolutionary Ecology and Conservation of an Enduring Carnivore. Island Press, Washington. [ Links ]
19. MANLY BFJ, P MILLER y LM COOK. 1972. Analysis of selective predation experiment. American Naturalist 106:719-736. [ Links ]
20. MARINO A y R BALDI. 2008. Vigilance patterns of territorial guanacos Lama guanicoe: The role of reproductive interests and predation risk. Ethology 114:413-423. [ Links ]
21. MURPHY K y TK RUTH. 2010. Diet and prey selection of a perfect predator. Pp 118-137, en: Cougar ecology and conservation (M Hornocker y S Negri, eds.). The University of Chicago Press, Chicago. [ Links ]
22. NOVACK AJ. 2003. Impacts of subsistence hunting on the foraging ecology of jaguar and puma in the Maya Biosphere Reserve, Guatemala. Tesis de Maestría, University of Florida, EE.UU. [ Links ]
23. NOVARO AJ, MC FUNES y RS WALKER. 2000. Ecological extinction of native prey of a carnivore assemblage in Argentine Patagonia. Biological Conservation 92:25-33. [ Links ]
24. NOVARO AJ y RS WALKER. 2005. Human-induced changes in the effect of top carnivores on biodiversity in the Patagonian Steppe. Pp 268-288, en: Large carnivores and biodiversity conservation (JC Ray, KH Redford, RS Steneck y J Berger, eds.). Island Press, Washington. [ Links ]
25. POLISAR J, I MAXIT, D SCOGNAMILLO, L FARRELL, ME SUNQUIST y JF EISEMBERG. 2003. Jaguars, pumas, their prey base, and cattle ranching: Ecological interpretations of a management problem. Biological Conservation 109:297-310. [ Links ]
26. RAEDEKE K. 1979. Population dynamics and socioecology of the guanaco (Lama guanicoe) of Magallanes, Chile. Tesis de Doctorado, University of Washington, Seattle, EE.UU. [ Links ]
27. RAU JM y JE JIMÉNEZ. 2002. Diet of puma (Puma concolor, Carnivora: Felidae) in coastal and Andean ranges of southern Chile. Studies on Neotropical Fauna and Environment 37:201-205. [ Links ]
28. REDFORD K y JF EISENBERG. 1992. Mammals of the Neotropics: The Southern Cone. Vol. 2. University of Chicago Press, Chicago. [ Links ]
29. TABER AB, AJ NOVARO, N NERIS y FH COLMAN. 1997. The food habits of sympatric jaguar and puma in the Paraguayan Chaco. Biotropica 29:204-213. [ Links ]
30. WOLFF F. 2001. Vertebrate ecology in Caatinga: A. Distribution of wildlife in relation to water. B. Diet of pumas Puma concolor and relative abundance of felids. Tesis de Maestría, University of Missouri, St. Louis, EE.UU. [ Links ]
31. YÁÑEZ JL, JC CÁRDENAS, P GEZELLE y FM JAKSIC. 1986. Food habits of the southernmost mountain lions Felis concolor in South America: Natural versus livestocked ranges. Journal of Mammalogy 67:604-606. [ Links ]
32. ZANÓN MARTÍNEZ JI, A TRAVAINI, S ZAPATA, D PROCOPIO y MA SANTILLÁN. 2012. The ecological role of native and introduced species in the diet of the puma Puma concolor in southern Patagonia. Oryx 46:106-111. [ Links ]

