Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. vol.22 no.1 Mendoza jun. 2015
ARTÍCULO
Mamíferos en relictos de bosque seco tropical del Tolima, Colombia
Leidy V. García-Herrera1, Leidy A. Ramírez-Fráncel1 y Gladys Reinoso Flórez2
1 Facultad de Ciencias, Maestría en Ciencias Biológicas, Grupo de Investigación en Zoología, Universidad del Tolima, Colombia [correspondencia: Leidy V. García-Herrera <lviviana0427@hotmail.com>].
2 Grupo de Investigación en Zoología, Universidad del Tolima, Colombia.
Recibido 29 mayo 2014.
Aceptado 14 octubre 2014.
Editor asociado: S Solari
RESUMEN.
El norte del departamento del Tolima presenta remanentes importantes de Bosque Seco Tropical (BST), ecosistema estratégico ya que alberga una alta diversidad y endemismo. Sin embargo, las actividades humanas han generado alteraciones considerables, incluida su disminución y transformación; por ello es importante documentar la diversidad de estos ambientes así como la respuesta de la fauna a las transformaciones antropogénicas. Este trabajo tiene como objetivo estimar la diversidad de mamíferos en un relicto de BST. Se establecieron 3 hábitats con diferencias estructurales a nivel de vegetación: bosque natural denso-bosque natural fragmentado, bosque de galería y ripario y mosaico de cultivos, pastos y espacios abiertos. Para el registro de mamíferos se usaron técnicas de captura (redes de niebla, trampas tipo Sherman) y técnicas de detección directa e indirecta (recorridos, búsqueda de rastros, trampas de huella). Se registraron 8 órdenes, 16 familias, 35 géneros y 39 especies. El mejor representado fue Chiroptera con 24 especies, seguido de Carnivora con 5. Las especies más abundantes fueron Heteromys anomalus (26 individuos), Carollia perspicillata (45) y Aotus griseimembra (7). El registro de Lontra longicaudis y A. griseimembra, especies consideradas en riego por la legislación colombiana, hacen de estos ambientes lugares de interés para la conservación de la biodiversidad de la región.
ABSTRACT.
Mammals in tropical dry forest fragments of Tolima, Colombia.
The northern Tolima has significant remnants of Tropical Dry Forest (TDF), strategic ecosystems that harbor a high diversity and endemism. However, human activities have produced substantial alterations, including reduction and transformation; therefore it is important to document the diversity of this environment and the response of wildlife to anthropogenic changes. Therefore, this study aimed to estimate the diversity of mammals in a TDF fragment. Three habitats with structural differences at the level of vegetation were sampled: dense natural forest-fragmented natural forest, gallery and riparian forest and mosaic of crops, pastures and open spaces. Mammal species were recorded using capture techniques (mist nets, Sherman traps) as well as direct and indirect signs (walks, hunting trails, traps trace). We recorded 8 orders, 16 families, 35 genera and 39 species. Chiroptera was best represented with 24 species, followed by Carnivora with 5. The most abundant species were Heteromys anomalus (26 individuals), Carollia perspicillata (45) and Aotus griseimembra (7). The presence of Lontra longicaudis and A. griseimembra species considered at risk by Colombian law, makes these environments interesting sites for the region's biodiversity conservation.
Palabras clave: Aotus griseimembra; Bosque seco tropical; Lontra longicaudis.
Key words: Aotus griseimembra; Lontra longicaudis; Tropical dry forest.
INTRODUCCIÓN
En Colombia se han registrado 492 especies de mamíferos (Solari et al., 2013). Sin embargo, hasta la fecha los estudios sobre la mastofauna resultan aún incipientes, dada la complejidad de sus ecosistemas (Hernández-Camacho et al., 1992) y la falta de políticas y acciones que permitan desarrollar investigaciones, inventarios o evaluaciones a nivel regional y nacional (Fernández, 2011). Es así como el conocimiento de la distribución geográfica de muchas especies está basado en algunas localidades con especímenes depositados en las colecciones de mayor trascendencia en el país, existiendo vacíos de información en importantes porciones del territorio colombiano.
Sin embargo, como ocurre en otros países neotropicales, las presiones antropogénicas han originado una serie de amenazas para la mastofauna. Entre las zonas de vida más afectadas se encuentra el Bosque Seco Tropical (BST). Este ecosistema se encuentra en proceso de deterioro, debido a la deforestación y el uso extensivo de las áreas boscosas para dar paso a la agricultura de pastos y cultivos migratorios (Maass, 1995). El BST reviste interés para las comunidades de mamíferos; en su ensamble convergen especies endémicas, cuyos individuos han desarrollado características fisiológicas y ecológicas para hacer frente a la estacionalidad climática típica de este bioma (Ceballos, 1995).
Los estudios para el BST colombiano se han centrado en pequeñas regiones geográficas o zonas muy intervenidas. Por lo tanto, actualmente no existen inventarios completos de mamíferos para todas las ecorregiones de BST del país (Pizanoy García, 2014). Dada la importancia del BST como ecosistema estratégico para su conservación, sumado a la poca información taxonómica y ecológica sobre la mastofauna que lo habita, en este trabajo se documenta la diversidad y abundancia de mamíferos asociados a 3 áreas con diferente grado de perturbación en el norte del departamento del Tolima, Colombia.
MATERIALES Y METODOS
Área de estudio
El sector muestreado corresponde a la vereda Chorrillo, municipio de Ambalema, cuya superficie es de 3.28 ha y se encuentra al norte del departamento del Tolima, a 62.3 km de Ibagué, ubicado a 4°50'27.78”N y 74º48'15.00”W, altura promedio de 253 m. Esta vereda parte de la cuenca del río Lagunillas —uno de los afluentes principales del río Magdalena— y está ubicada en el flanco derecho de la Cordillera Central, con una extensión de 795 km2. El estudio aquí implementado abarcó 3 coberturas con vegetación contrastante. De acuerdo con el sistema de coberturas IGAC-COLOMBIA, estas corresponden a: 1) Bosque con vegetación secundaria; 2) Bosque de galería; y 3) Agroecosistema. En el sistema Corine Land Cover —que aquí se sigue— equivalen a: 1) Bosque natural denso-bosque natural fragmentado (BS); 2) Bosque de galería y ripario (BG); y 3) Mosaico de cultivos, pastos y espacios abiertos (MC) (Fig. 1). El BS presenta una estructura compleja compuesta por 2 estratos: arbóreo (aproximadamente 16 m) y arbustivo (<10 m); se encuentra ubicado a 4°50'27.78”N y 74º48'15.00”W a una altitud de 249 m, distanciado del bosque de galería y ripario por 1.83 km y por 0.89 km del mosaico de cultivos, pastos y espacios abiertos. El BG cuenta con 3 estratos: arbóreo (aproximadamente 13 m), arbustivo alto (10-12 m) y bajo (<10 m), ubicado a 4°50'54.48”N y 74º48'52.57”W, a 242 m de altitud y distanciado por 0.86 km del MC. Por su parte, este último se encuentra constituido por un cultivo transitorio no tecnificado de maíz y plátano rodeado de rastrojo y se ubica a 4°51'15.01”N y 74º48'24.48”W,232 m.
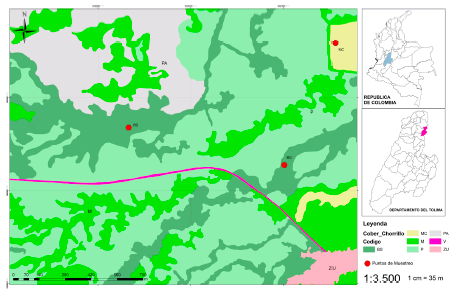
Fig. 1. Localización geográfica de la vereda Chorrillo, municipio de Ambalema, Tolima-Colombia. BS = bosque natural denso-bosque natural fragmentado; BG = bosque de galería y ripario; MC = mosaico de cultivos, pastos y espacios abiertos; C = cultivo; P = pasto; PA = pastizales arbolados; V = vías; ZU = zonas urbanas.
Colecta de datos
Se realizaron campañas en los meses de abril, julio, octubre y noviembre de 2013, durante 3 días consecutivos. Se emplearon técnicas de captura, observación directa e indirecta y se clasificaron las especies registradas de acuerdo a su masa corporal, siguiendo la propuesta de Sánchez et al. (2004). Se consideraron como pequeños mamíferos terrestres aquellos con masa <150 g, medianos entre 150 g y 5 kg y grandes con una masa >5 kg. Para el registro de pequeños mamíferos voladores se instalaron 5 redes de niebla de 12 x 2.5 m, calibre de 36 mm y ojo de 1”1/2, que se ubicaron ad libitum teniendo en cuenta las características del área de estudio y la composición vegetal de la misma. Debido a la heterogeneidad del paisaje, el muestreo se realizó siguiendo un diseño por conglomerados con el fin de abarcar la mayor área posible. Las redes permanecieron abiertas entre las 18:00 y las 24:00, con una intensidad de una jornada de 120 horas/red/ cobertura/muestreo. Para los pequeños mamíferos no voladores se emplearon un total de 150 trampas tipo Sherman. En cada cobertura se instalaron 50 trampas dispuestas en un cuadrante de 72 x 20 m, distanciadas por 8 m y con 2 trampas por estación de trampeo. Las trampas se dispusieron a nivel del suelo, en la hojarasca y sobre troncos caídos. Se cebaron diariamente empleando un atrayente generalista, a base de maní tostado, molido e impregnado con esencia de vainilla y banano. Para el registro de los medianos y grandes mamíferos se emplearon 9 cámaras trampa Bushnell Trophy, desplegándose una cámara por cada 2 km2, durante 3 días consecutivos; no se empleó ningún tipo de cebo con el fin de minimizar la atracción de una especie en particular. Para el rastreo de indicios en el BG y BS se empleó un transecto lineal, de longitud de 2 km; para el caso particular del MC la longitud fue de 0.32 km, realizándose observaciones entre las 07:00 y las 17:00, a una velocidad promedio de 1 km/hora. Los avistamientos, huellas, heces, restos y madrigueras fueron tomados como indicios de acuerdo a lo propuesto por Wilson et al. (1996); se registraron datos como especie, número de indicio, hora de encuentro, ubicación y tipo de formación vegetal, siguiendo la metodología de Rumiz et al. (1998). Para la identificación de huellas se elaboró un molde con yeso odontológico (polvo de fraguado rápido tipo “III”), siguiendo lo propuesto por Aranda (2000); adicionalmente, a todas las huellas se les realizó un dibujo en acetato transparente, colocándolo por encima de la huella y trazando la silueta de la misma con ayuda de un marcador de punta fina y tinta indeleble (Smallwood y Fitzhugh, 1995). Los moldes en yeso y los dibujos en acetato fueron confrontados contra el catálogo de huellas de los mamíferos de México (Aranda, 2000). Los especímenes e impresiones de huellas colectados fueron depositadas en la Colección Zoológica de la Universidad del Tolima – Área mastofauna (M), bajo los números de catálogo 1304 al 1341. El permiso ambiental fue suministrado por CORTOLIMA N°030 del 31 de julio 2013.
Análisis de datos
Para evitar sobreestimación en el número total de mamíferos capturados, los pequeños mamíferos no voladores y voladores se marcaron individualmente mediante elastómeros de color verde y rosado, siguiendo una clave de marcaje. Las huellas se contaron como individuos independientes en función de la cercanía o lejanía en las medidas de cada registro, los indicios se cuantificaron con un solo individuo en todo el muestreo, al igual que las tropas de primates, por lo que solo se consideraron los organismos de la primera tropa avistada. Entre los pequeños mamíferos no voladores se consideraron los roedores y los marsupiales que fueron capturados. Se realizó el análisis de riqueza y diversidad y se empleó el éxito de captura expresado como el número de individuos capturados por 100 noches-trampa de acuerdo con Ochoa (2000), con el fin de establecer la abundancia relativa. El esfuerzo de captura para los mamíferos voladores se analizó a través de las horas-red totales trabajadas y el éxito de captura, teniendo en cuenta el número de individuos totales capturados respecto al esfuerzo de muestro (individuos/horas-red totales) x 100. Se calculó el porcentaje de abundancia relativa para las especies encontradas; la abundancia fue tomada como el número de individuos en cada una de las coberturas evaluadas. La representatividad del inventario se calculó a través de la curva de acumulación de especies, representada por el número de especies acumuladas a través del tiempo o unidad de esfuerzo (meses evaluados). Considerando que la riqueza observada se diferencia de las reales con una magnitud que no se conoce pero basados en el supuesto de que se aplicaron los mismos métodos, se puede suponer que dichas diferencias son proporcionales entre las 3 coberturas evaluadas, permitiendo realizar comparaciones entre ellas. Para estimar la riqueza de especies, se empleó la ecuación de Clench, modelo que permite encontrar el número esperado de especies, así como la proporción de la fauna muestreada. La diversidad en cada área evaluada se determinó a través del exponencial del índice de Shannon (eH'), cuyas unidades son el número de especies efectivas en las comunidades (Jost, 2006) y el índice invertido de Simpson (Cinv). Las curva de acumulación de especies, así como el exponencial de Shannon e inverso de Simpson fueron calculados con el programa EstimateS, version 9 (Colwell, 2013). Finalmente, para calcular la diversidad β entre las áreas evaluadas se calculó el índice Morisita-Horn, el cual es influenciado por la riqueza de especies y el tamaño de la muestra (Moreno, 2000). Para medianos y grandes mamíferos se analizó la riqueza, abundancia relativa y diversidad en cada una de las coberturas evaluadas, se realizó la curva de rarificación y se calculó la diversidad inversa de Simpson y el exponencial de Shannon.
RESULTADOS
En 5 meses de muestreo se registraron 8 órdenes, 16 familias, 35 géneros y 39 especies, lo que corresponde al 7.92% de la diversidad de la mastofauna presente a nivel nacional. Entre los pequeños mamíferos no voladores se registraron únicamente 2 especies (Marmosa robinsoni y Heteromys anomalus), con un éxito de captura total de 0.02 individuos/trampas-noche. M. robinsoni registró 19 capturas y H. anomalus, 17. El éxito de captura para cada cobertura varió; en el BS se obtuvo la mayor representación con el 44.44% (16 individuos), seguido por el BG con 36.11% (13 individuos)mientras en el MC solo se registró el 19.44% (7 individuos). Con un esfuerzo de 1920 horas-red totales se capturaron 223 quirópteros, correspondientes a 24 especies, con un éxito de captura de 0.44 individuos/horas-red. La familia Phyllostomidae fue la mejor representada con el 66.7% (14 especies), con 6 especies de la subfamilia Stenodermatinae, 5 de la Phyllostominae, 2 de Carolliinae y Glossophaginae, y 1 de Desmodontinae. La segunda mayor representación la obtuvo Vespertilionidae con el 16.63% (4 spp.) con un solo miembro en la subfamilia Myotinae y 3 en Vespertilioninae. Por su parte, la familia Emballonuridae y Molossidae aportaron 2 y 1 especie, respectivamente. La especie más abundante fue Carollia perspicillata con el 20.17% de los registros (45 individuos) seguido de Phyllostomus discolor con el 14.34% (32 individuos); Peropteryx macrotis, Phylloderma stenops, Sturnira tildae, Molossus molossus y Eptesicus fuscus fueron registradas solo una vez.
La curva de acumulación de especies obtenida se ajustó a los datos del modelo de Clench (r = 0.98), y mediante los estimados a = 15.12 y b = 0.38 se predijo el número de especies esperadas (n = 39). Las especies observadas frente a las esperadas equivalen al 61.53% (Fig. 2). Al evaluar la calidad del inventario calculando la pendiente al final de la curva, esta fue de 0.51, indicando que se logró un inventario bastante completo y altamente fiable.

Fig. 2. Curva de acumulación de especies de mamíferos voladores para todos los ambientes muestreados, de acuerdo en lo observado (Sobs) con losestimadores paramétricos (Modelo de Clench) e intervalos de confianza del 95% (IC).
Al comparar las 3 medidas de diversidad obtenidas en las coberturas evaluadas, la mayor diversidad efectiva se registró en el BG (eH' = 2.13), indicando que la cobertura es más diversa que el promedio de las otras dos. El inverso de Simpson fue más alto en el mosaico de cultivos, pastos y espacios abiertos (Cinv = 9.21) en comparación con los resultados obtenidos en los bosques (Tabla 1).
Tabla 1 Representación taxonómica de la mastofauna e índices de diversidad de Simpson (Cinv) y exponencial de Shannon (eH') para los 3 tipos de coberturas evaluadas en la vereda Chorrillo (BS, BG, MC; ver referencias en leyenda Fig. 1), municipio de Ambalema, Tolima-Colombia

El valor del índice de similitud Morisita- Horn para los 3 tipos de coberturas fue de 0.93 indicando que son muy similares, pese a la diferencia en el número de especies. Sin embargo la cobertura cultivos, pastos y espacios abiertos y el bosque de galería y ripario en términos de composición de especies resultaron similares en el 84% (Fig. 3).
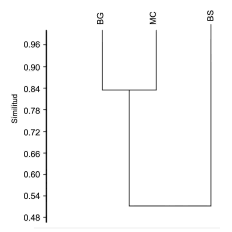
Fig. 3. Índice de similitud Morisita-Horn, para el ensamblaje de murciélagos para las 3 coberturas evaluadas en la vereda Chorrillo, municipio de Ambalema, Tolima- Colombia.BS = bosque natural denso-bosque natural fragmentado, BG = bosque de galería y ripario, MC = mosaico de cultivos, pastos y espacios abiertos.
A partir de este estudio se completa el primer listado de medianos y grandes mamíferos para el BST del departamento del Tolima, el cual incluye 7 géneros, 11 familias y 13 especies (Apéndice 1). El orden mejor representado fue Carnivora, con 5 especies (Puma yagouaroundi, Cerdocyon thous, Galictis vittata, Lontra longicaudis y Procyon cancrivorus).
Mediante la curva de rarificación y bajo el supuesto de que los 3 tipos de coberturas hubieran sido evaluadas a través de 15 puntos se estimó la mayor riqueza de especies para el BG (8 spp.), seguido del BS (6 spp.) y el M (2 spp.). Sin embargo, las curvas de rarificación no alcanzaron la asíntota, por lo tanto se espera encontrar más especies en dichos sitios al aumentar el esfuerzo de muestreo (Fig. 4). Las especies Aotus griseimembra, Mazama zetta y Didelphis marsupialis reportaron las abundancias relativas más altas, mientras que entre las especies con la menor abundancia se encuentran C. thous, L. longicaudis, P. cancrivorus y Pecari tajacu (Fig. 5).
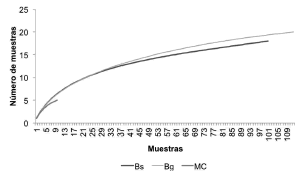
Fig. 4. Curva de rarificación para los ensamblajes de mamíferos medianos y grandes en las 3 coberturas evaluadas en la vereda Chorrillo, municipio de Ambalema, Tolima-Colombia. BS = bosque natural denso-bosque natural fragmentado, BG = bosque de galería y ripario, MC = mosaico de cultivos, pastos y espacios abiertos.

Fig. 5. Abundancia relativa de medianos y grandes mamíferos en los 3 tipos de coberturas evaluadas en la vereda Chorrillo, municipio de Ambalema, Tolima-Colombia. BS = bosque natural denso-bosque natural fragmentado, BG = bosque de galería y ripario, MC = mosaico de cultivos, pastos y espacios abiertos.
Al evaluar la diversidad que tendría este ensamblaje integrado por i especies igualmente comunes y al comparar la magnitud de la diferenciación entre las coberturas evaluadas se observó que el bosque natural denso-bosque natural fragmentado es 10.38 más diverso que el promedio de las otras coberturas evaluadas. Por su parte el inverso de Simpson manifestó un mayor valor en la cobertura mosaico de cultivo y de espacios abiertos, lo que significa que la probabilidad de que dos individuos seleccionados aleatoriamente en el hábitat pertenezcan a la misma especie es 1 en 9.03 (Tabla 1).
Las especies Chironectes minimus, L. longicaudis, P. cancrivorus y M. zetta fueron reportadas exclusivamente a través del registro de huellas. En combinación con la metodología anterior y de cámaras-trampa solo se logró capturar la especies P. yagouaroundi, mientras que el resto de las especies se registraron mediante avistamientos a través de los recorridos de transectos (Fig. 6).
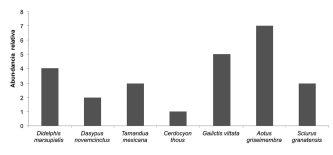
Fig. 6. Abundancia relativa por transectos de las especies de mamíferos medianos y grandes en la vereda Chorrillo, municipio de Ambalema, Tolima- Colombia.
DISCUSIÓN
La riqueza de especies de mamíferos registrados de forma acumulada en las zonas estudiadas (39 especies) representa el primer inventario de la mastofauna del Tolima, ya que los estudios previos se han concentrado en grupos particulares, como el análisis de la diversidad de quirópteros en una franja altitudinal (Bejarano-Bonilla et al., 2007) o listados de especies de quirópteros a escala departamental (Galindo-Espinosa et al., 2010).
Este estudio se constituye en una herramienta importante para adelantar acciones de conservación de este ecosistema (BST), dado que en un contexto global, la zona representa aproximadamente el 40% de las áreas boscosas del trópico (Bautista et al., 2004) y enfrenta un acelerado deterioro.
Se registraron tan solo 2 especies pertenecientes a los pequeños mamíferos no voladores, pese a que este grupo es el segundo mejor representado de la mastofauna colombiana, con 12 especies que corresponde al 25% de las 492 especies de mamíferos reportadas para el país (Solari et al., 2013).
Los resultados obtenidos evidencian a Chiroptera como el orden mejor representado con 24 especies, información que coincide con lo observado para la mastofauna del Neotrópico (López-Higareda, 2006) y para la cordillera central colombiana (Castaño y Corrales, 2010).
Se resalta que en este estudio se registra el 33% de la quiropterofauna reportada para el Tolima (72 especies; Galindo-Espinosa et al., 2010), porcentaje que es significativo, dado que es solo 1 localidad la muestreada y una única zona de vida. Los resultados obtenidos en este estudio son comparables con los reportados por otros estudios realizados en el BST, específicamente en los departamentos de Córdoba y Sucre. En el estudio realizado por Ballesteros et al. (2007) se registran 15 especies, de las cuales se comparten 7, mientras Calonge et al. (2010) reportaron 20 especies, compartiendo 12.
Stenodermatinae presentó la mayor abundancia relativa compatible con una mayor resistencia de sus representantes a las transformaciones del ambiente y posiblemente se benefician con la fragmentación, ya que utilizan tanto bosques continuos, como ambientes transformados y remanentes, vegetación riparia, vegetación secundaria e incluso árboles y arbustos aislados en los pastizales (Galindo-Gonzales et al., 2000). C. perspicillata registró la abundancia relativa más alta del ensamble, siendo una especie asociada a vegetación secundaria (Muñoz-Saba et al., 1999). Además, esta especie se puede adaptar a diferentes situaciones de oferta alimentaria, ya que consumen especies vegetales disponibles cuando el recurso especifico escasea (Loayza et al., 2006).
Al estimar la curva de acumulación de especies con la ecuación Clench para todo el ensamblaje de murciélagos se obtiene que el inventario está dentro de lo aceptable, puesto que se registró más del 50% de las especies esperadas. Se evidencian diferencias en la diversidad obtenida en cada una de las coberturas evaluadas, lo cual podría estar influenciado por los procesos de transformación, puesto que la fragmentación es un proceso que genera la reducción del tamaño original del hábitat, provocando un cambio en sus propiedades, lo que aumenta el tamaño del borde, modifica el número de fragmentos (parches) e incrementa el aislamiento entre estos (Fahrig, 2003; Bennett, 2004); asimismo los cambios espaciales producen efectos a través de los diferente niveles de organización biológica, y en sí, en los procesos ecológicos modificando el funcionamiento de los ecosistemas (Bennett, 2004). Las formaciones bosque de galería y ripario y mosaico de cultivos y de espacios abiertos presentan la mayor similitud en la comunidad de murciélagos, posiblemente empleen estas áreas como corredores estratégicos de forrajeo o refugio (Kalko y Handley, 2001).
Los muestreos de mamíferos medianos y grandes proveen los primeros registros existentes en los remanentes de bosques intervenidos en el norte del departamento del Tolima. En el contexto nacional y de acuerdo con Solari et al. (2013), para este departamento no se registraban Tamandua mexicana, Artibeus planirostris, G. vittata, M. zettay y A. griseimembra, especies que han sido capturadas o avistadas en el presente estudio y se constituyen en el primer registro formal para el departamento del Tolima.
Mediante el análisis de rarificación, se estableció que pese a la representatividad de los muestreos, la curva no alcanzó una asíntota en ninguna de las 3 coberturas, lo cual sugiere que aún faltan más muestreos para registrar todas las especies de mamíferos medianos y grandes presentes en estas coberturas.
Los valores de riqueza y abundancia fueron altos en las coberturas boscosas (BS y BG), lo cual sugiere que la fauna fuera de estas áreas se ha visto reducida como consecuencia de la pérdida y fragmentación del hábitat y la cacería con fines comerciales, actividad que es reiterativa en el departamento del Tolima, en donde los mamíferos ocupan el segundo lugar en el comercio de la región (Rojas-Briñez, 2011).
En este sentido, el orden Primates es el más comercializado, representado por las familias Aotidae, Atelidae, Cebidae y Pitheciidae (Rojas- Briñez, 2011), lo que se encuentra relacionado con su fácil identificación al conformar grupos sociales (Duarte, 2007) y su alta demanda en el mercado, ya sea como mascotas o como modelos para investigaciones biomédicas y etológicas o del comportamiento animal. De las 13 especies registradas de medianos y grandes mamíferos, 8 se encuentran en el BG, cobertura que puede ser usada como sitio de forrajeo por las especies, además que la vegetación asociada a los cuerpos de agua provee de sitios de alimentación y perchas temporales nocturnas que pueden ser usadas por estas especies.
Basado en lo anterior, la presencia de G. vittata y L. longicaudis estaría determinada por factores como la disponibilidad de presas, la presencia de árboles y arbustos (cobertura de escape) y variables estructurales propias del bosque de ribera bien conservado, el cual debe ofrecer buena cobertura de hojarasca, que es empleada como cobertura térmica (Restrepo y Botero-Botero, 2011).
Adicionalmente, la riqueza de plantas leñosas y desarrollo vertical, aportan alimento y refugio, así como un hábitat más estructurado que permitiría la coexistencia de una mayor diversidad de especies (Bernard y Fenton, 2002).
En contraste, el bosque natural denso-bosque natural fragmentado y el mosaico de cultivos, pastos y espacios abiertos albergan muy pocas especies, lo cual permite inferir que las condiciones de refugio, alimentación y percha son reducidas en estas áreas. Esto puede ser inducido por los procesos antrópicos que son muy marcados en estas áreas, producto de la tala, extensión de la frontera ganadera y cultivos no tecnificados inmersos en matorrales y pastos naturales.
Evaluar la composición de la mastofauna a través de la variación espacial permitió ampliar la información sobre este grupo faunístico en la cordillera central colombiana. Estos resultados constituyen el primer inventario de mamíferos presentes en la región del valle del Magdalena y evidencian la presencia de especies como L. longicaudis y A. griseimembra, que revisten gran interés de conservación, dado que son especies que se encuentran catalogadas en estado vulnerable de acuerdo a la Resolución 192 del 10 de febrero de 2014 del Ministerio de Ambiente y Desarrollo, por la cual se establece el listado de especies silvestres amenazadas de la diversidad biológica colombiana.
AGRADECIMIENTOS
Las autoras agradecen al Grupo de Investigación en Zoología por el apoyo económico y logístico; a Emma Yicel Galindo y Karina Alexandra Gutiérrez por su apoyo técnico; a Andrés Fabián Santos, Andrea Tarquino, Fredy Quintero e integrantes del Semillero de Mastofauna por su acompañamiento en las tareas de campo; a Jaider Manuel Peña, Héctor Fabio Cruz y Miguel Moreno por su valiosa colaboración en el desarrollo del proyecto. Deseamos agradecer a los revisores anónimos, por sus comentarios y correcciones a la redacción del texto. Las 2 primeras autoras agradecen de manera especial a COLCIENCIAS por la beca otorgada en el programa “Jóvenes Investigadores e Innovadores Virginia Gutiérrez de Pineda” 566-2012. Esta es la contribución 436 al Grupo de Investigacion en Zoología de la Universidad del Tolima (GIZ-UT).
LITERATURA CITADA
1. ARANDA J. 2000. Manual para el rastreo de mamíferos silvestres de México. Arte Editorial, México. [ Links ]
2. BALLESTEROS J, J RACERO y M NÚÑEZ. 2007. Diversidad de murciélagos en cuatro localidades de la zona costanera del departamento de Córdoba, Colombia. Revista de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad de Córdoba 12:1013-1019. [ Links ]
3. BAUTISTA LM, JT GARCÍA, RG CALMAESTRA, C PALACÍN, C MARTÍN, M MORALES, R BONAL y J VIÑUELA. 2004. Effect of weekend road traffic on the use of space by raptors. Conservation Biology 13:726-732. [ Links ]
4. BEJARANO-BONILLA D, A YATE-RIVAS y M BERNAL-BAUTISTA. 2007. Diversidad y distribución de la fauna quiróptera en un transecto altitudinal en el departamento del Tolima, Colombia. Caldasia 29:297-308. [ Links ]
5. BENNETT A. 2004. Enlazando el paisaje: el papel de los corredores y la conectividad en la conservación de la vida silvestre. UICN San José, Costa Rica. [ Links ]
6. BERNARD E y MB FENTON. 2002. Species diversity of bats (Mammalia: Chiroptera) in forest fragments, primary forests, and savannas in Central Amazonia, Brazil. Canadian Journal of Zoology 80:1124-1140 [ Links ]
7. CALONGE B, IM VELA-VARGAS y J PÉREZ-TORRES. 2010. Dieta y estructura trófica del conjunto de murciélagos frugívoros en una finca con remanentes de bosque seco tropical, Córdoba, Colombia. Revista de la Facultad de Medicina Veterinaria y Zootecnia de la Universidad de Córdoba 15:1938-1943. [ Links ]
8. CASTAÑO JH y JD CORRALES. 2010. Mamíferos de la cuenca del río La miel, Caldas: Diversidad y uso cultural. Boletín Científico del Museo de Historia Natural, Universidad de Caldas 1:56-78. [ Links ]
9. CEBALLOS G. 1995. Vertebrate diversity, ecology and conservation in neotropical dry forests. Pp195-220, en: Seasonally dry tropical forests (S Bullock, H. Mooney y E. Medina, eds.). Cambridge University Press, Cambridge, UK. [ Links ]
10. COLWELL RK. 2013. EstimateS (Statistical estimation of species richness and shared species from samples). Version 9.1. Persistent http://purl.oclc.org/estimates [ Links ]
11. FAHRIG L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34:487-515. [ Links ]
12. FERNÁNDEZ F. 2011. The greatest impediment to the study of biodiversity in Colombia. Caldasia 33(2):2-5. [ Links ]
13. GALINDO-ESPINOSA E, K GUTIERREZ-DIAZ y G REINOSO. 2010. Lista de los quirópteros del departamento del Tolima, Colombia. Biota Colombiana 11:107-116. [ Links ]
14. GALINDO-GONZÁLEZ J, S GUEVARA y VJ SOSA. 2000. Bat- and bird-generated seed rains at isolated trees in pastures in a tropical rainforest. Conservation Biology 14:1693-1703. [ Links ]
15. HERNÁNDEZ C, TH WALSCHBURGER, R ORTIZ y A HURTADO. 1992. Sobre el origen y distribución de la biota suramericana y colombiana. Pp. 55-104, en: Diversidad biológica de Iberoamérica. (G. Halffter, ed.). Secretaría de Desarrollo, México D.F. [ Links ]
16. JOST L. 2006. Entropy and diversity. Oikos 113:363-375. [ Links ]
17. KALKO EKV y CO HANDLEY. 2001. Neotropical bats in the canopy: Diversity, community structure, and implications for conservation. Plant Ecology 153:319-333. [ Links ]
18. LOAYZA A, R RIOS y D LARREA-ALCÁZAR. 2006. Disponibilidad de recurso y dieta de murciélagos frugívoros en la Estación Biológica Tunquini, Bolivia. Ecología en Bolivia 41:7-23. [ Links ]
19. LÓPEZ-HIGAREDA D. 2006. Mastofauna del bosque mesófilo de montaña de Tenango (Municipio de Tenango de Doria, Hidalgo). Tesis de Licenciatura, Facultad de Ciencias Naturales, Universidad Nacional Autónoma de México, México. [ Links ]
20. MAASS JM. 1995. Tropical deciduous forest conversion to pasture and agriculture. Pp. 399−422, en: Seasonally dry tropical forests (SH Bullock, HA Mooney y E Medina, eds.). Cambridge University Press, Cambridge, UK.
21. MORENO CE. 2001. Métodos para medir la biodiversidad. Manuales y Tesis SEA 1:4-84. [ Links ]
22. MUÑOZ-SABA Y, H LÓPEZ-ARÉVALO y A CADENA. 1999. Aportes al conocimiento de la ecología de los murciélagos de los afloramientos de mármoles calizas, Sector del río Claro (Antioquia, Colombia). Revista Academia Colombiana de Ciencias 23:473-486. [ Links ]
23. OCHOA G J. 2000. Efectos de la extracción de maderas sobre la diversidad de mamíferos pequeños en bosques de tierras bajas de la Guayana venezolana. Biotropica 32:146-164. [ Links ]
24. PIZANO C y H GARCÍA. 2014. El Bosque Seco Tropical en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH), Bogotá [ Links ].
25. RESTREPO C y A BOTERO-BOTERO. 2012. Ecología trófica de la nutria neotropical Lontra longicaudis (Carnivora, Mustelidae) en el río La Vieja, alto Cauca, Colombia. Boletín Científico del Museo de Historia Natural Universidad de Caldas 16:207-214. [ Links ]
26. ROJAS-BRIÑEZ DK. 2011. Comercio de fauna silvestre en el departamento del Tolima, Colombia, bajo el contexto de la demanda internacional de especies. Tesis de maestría, Facultad de Ciencias, Universidad Internacional de Andalucía, Baeza, Jaén, España. [ Links ]
27. RUMIZ DI, C EULERT y R ARISPE. 1998. Evaluación de la diversidad de mamíferos medianos y grandes en el Parque Nacional Carrasco (Cochabamba, Bolivia). Revista Boliviana de Ecología y Conservación Ambiental 4:77-90. [ Links ]
28. SÁNCHEZ F, P SÁNCHEZ-PALOMINO y A CADENA. 2004. Inventario de mamíferos en un bosque de los Andes centrales de Colombia. Caldasia 26:291-309. [ Links ]
29. SOLARI S, Y MUÑOZ-SABA, JV RODRÍGUEZ-MAHECHA, TR DEFLER, H RAMÍREZ-CHAVES y F TRUJILLO. 2013. Diversidad, endemismo y conservación de los mamíferos de Colombia. Mastozoología Neotropical 20:301-365. [ Links ]
30. SMALLWOOD K y E FITZHUGH. 1995. A track count for estimating mountain lion Felis concolor californica population trend, Biological Conservation 71:251-259. [ Links ]
31. WILSON DE, FR COLE, JD NICHOLS, R RUDRAN y MS FOSTER. 1996. Measuring and monitoring biological diversity: Standard methods for mammals. Smithsonian Institution. Washington, D.C. [ Links ]
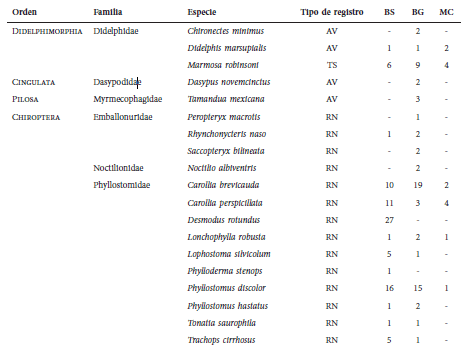
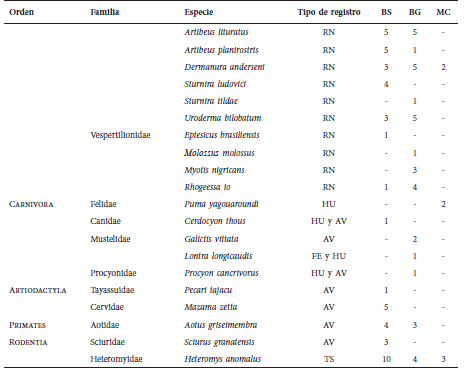
Mamíferos (en número de ejemplares) registrados para cada cobertura evaluada en la vereda
Chorrillo, municipio de Ambalema, Tolima-Colombia. BS = bosque natural denso-bosque natural
fragmentado, BG = bosque de galería y ripario y MC = mosaico de cultivos, pastos y espacios
abiertos. Tipo de registro, AV = Avistamientos, TS = trampa tipo Sherman, RN = redes de
niebla, FE = feca y HU = huella.