










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. vol.22 no.1 Mendoza jun. 2015
ARTÍCULO
Relación de los ataques de Desmodus rotundus con el manejo del ganado caprino y algunas características del hábitat en la prepuna de Bolivia
M. Isabel Moya1,2,5, Luis F. Pacheco2,3 y Luis F. Aguirre4,5
1 Museo Nacional de Historia Natural, Cota Cota, Calle 26 s/n, La Paz, Bolivia. Teléfonos: (591) 2-2795364, (591) 73293884 [correspondencia: M. Isabel Moya <isamoyadiaz@gmail.com>].
2 Colección Boliviana de Fauna, Cota Cota, Calle 27 s/n, Campus Universitario, La Paz, Bolivia. Teléfono: (591) 2-2792582.
3 Instituto de Ecología, Carrera de Biología, Universidad Mayor de San Andrés, Cota Cota, Calle 27 s/n, La Paz, Bolivia. Teléfono: (591) 2-2792416, int. 5.
4 Centro de Biodiversidad y Genética, Universidad Mayor de San Simón, Calle Sucre, Parque La Torre, Cochabamba, Bolivia. Teléfono: (591) 4-4540796.
5 Fundación Programa para la Conservación de los Murciélagos y la Biodiversidad, Urbanización las Magnolias 2, Nº 30C, Cochabamba, Bolivia.
Recibido 14 enero 2014.
Aceptado 5 diciembre 2014.
Editor asociado: M Sandoval
RESUMEN.
El conflicto entre las actividades humanas y los murciélagos vampiros (Desmodus rotundus) ha sido evidente en la Prepuna boliviana en las últimas décadas. Se han reportado frecuentes ataques al ganado caprino, lo que condujo a la aplicación de estrategias de control inadecuadas de las poblaciones del vampiro. Con el objetivo de aportar al manejo y mitigación del conflicto, se evaluó si la incidencia de ataques está asociada a factores como la distribución y la abundancia del ganado, la distancia de los corrales a los principales refugios de los vampiros, el tipo de hábitat y la abundancia relativa del vampiro. Los resultados sugieren que la incidencia de ataques decrece con la cantidad de ganado y con la distancia de los corrales a los refugios de los vampiros, y se incrementa con la abundancia relativa de vampiros en la zona. Por otro lado, la abundancia de vampiros está relacionada positivamente con la disponibilidad de refugios cercanos a sus áreas de forrajeo pero no así con la abundancia de ganado. Proponemos que el factor principal que limita la abundancia de vampiros, e indirectamente determina la incidencia de ataques, es la disponibilidad de refugios cercanos a los corrales. Es así que las localidades que se encontraban cerca de refugios presentaban valores altos de abundancia de vampiros y de incidencia de ataques al ganado. Con base en estos resultados, sugerimos la regulación de operaciones mineras que involucran potenciales refugios de vampiros y la implementación de programas de monitoreo de la incidencia de ataques como un indicador de la abundancia del vampiro.
ABSTRACT.
Relationships between vampire bat (Desmodus rotundus) attacks to goats, livestock management, and some habitat characteristics in the Bolivian prepuna.
The conflict between human activities and vampire bats (Desmodus rotundus) has been evident in the Bolivian Prepuna for decades. Vampire attacks on domestic goats have been frequently reported, and led to the application of inadequate control strategies of vampire populations. In order to contribute to the management and mitigation of this conflict, we assessed whether the incidence of attacks is associated with factors like the distribution and abundance of livestock, the distance from the stockyards to vampire roosts, the habitat type, and the relative abundance of the vampire. The results suggest that the incidence of attacks decreases with the density of goats and the distance from stockyards to vampire roosts, while it increases with the relative abundance of vampires in the area. Besides, the abundance of vampires is positively correlated to the availability of roosts near their foraging areas and is not correlated to livestock abundance. We propose that the main factor limiting the abundance of vampires, and indirectly determining the incidence of attacks, is the availability of roosts near foraging areas. Coincidently, the localities situated near roosts presented higher abundance of vampires and recorded the highest incidence of attacks on livestock. Based on our results, we propose the regulation of mining operations involving potential vampire refuges. We also suggest the implementation of monitoring programs of the incidence of attacks as an indicator of the vampire abundance.
Palabras clave: Bolivia; Conflicto actividades humanas–vampiros; Incidencia de ataques; Refugios; Socavones; Vampiro.
Key words: Bolivia; Incidence of attacks; Human activitiy–vampire conflict; Mining tunnels; Roosts; Vampire bat.
INTRODUCCIÓN
Las especies que ocurren naturalmente encuentran niveles de coexistencia estables y resilientes, al considerarse escalas de tiempo grandes (Odum, 1971). Es así que, las especies nativas están mejor adaptadas a coexistir con depredadores, competidores y enfermedades transmitidas por otras especies silvestres nativas (Messmer, 2000). La modificación de las comunidades faunísticas, por ejemplo, por la introducción de especies exóticas como el ganado, puede provocar cambios numéricos en algunas poblaciones silvestres; en el caso de los murciélagos vampiros (Desmodus rotundus, en adelante vampiros), como una respuesta a la oferta de un recurso alimenticio adicional, este cambio es positivo y origina conflictos con los humanos por las implicaciones económicas y por el efecto que esta interacción tiene sobre la salud pública. Por tal motivo, en algunos casos los vampiros son considerados plaga (Ojasti, 2000; Conover, 2001).
Históricamente se han desarrollado estudios sobre los conflictos que se generan entre distintos organismos en estado silvestre y las actividades humanas, incluyendo el ataque de animales a campos agrícolas (Oppenheimer y Lange, 1969; Pérez y Pacheco, 2006; Romañach et al., 2007), los ataques de carnívoros grandes al ganado (Allen y Sparkes, 2001; Beckmann et al., 2004; Pacheco et al., 2010; Zarco-González et al., 2012) y aquellos referidos a la transmisión de rabia por vampiros (Belotto, 2001; Constantine, 2009; Carvalho-Costa et al., 2012; Nuñez et al., 2012), incluyendo a seres humanos (Shneider et al., 2009; Johnson et al., 2010). La mortalidad de ganado bovino por causa de rabia transmitida por vampiros fue estimada para la década de los 60 en un millón de cabezas por año en Latinoamérica (Badger y Schmidt, 1979) y los mismos autores indican que la rabia era el problema de salud animal más grave en Bolivia hasta 1972. Sin embargo, aunque uno de los conflictos más comunes en áreas ganaderas es el que se establece entre humanos y vampiros, específicamente D. rotundus, son escasos aquellos estudios destinados a entender la problemática subyacente, en particular respecto a la relación de variables ecológicas de las poblaciones de murciélagos y de las poblaciones de animales domésticos o aspectos relacionados al manejo del ganado (Mayen, 2003).
Se sabe que D. rotundus es el único murciélago que se alimenta de sangre de ganado doméstico (Brown, 1994; Barclay y Harder, 2003), lo que redunda en pérdidas económicas para los ganaderos. La respuesta de la gente es la realización de acciones de control de las poblaciones de vampiros, medidas que por lo general son inadecuadas y no selectivas y que ponen en riesgo poblaciones de otras especies de murciélagos (Aguirre, 2007; MMAyA, 2009).
Este estudio estuvo enfocado a entender el conflicto entre vampiros y ganadería, luego de varias noticias de prensa (La Patria, 2006; La Razón, 2006; El Potosí, 2006) que reportaban la severidad del problema en la Provincia Nor Chichas del departamento de Potosí, región característica de la Prepuna boliviana (López, 2000). Es interesante notar que, según la percepción de los propios pobladores, los socavones que quedaron abandonados luego del cierre de las actividades mineras se constituyeron en refugios para los murciélagos, especialmente para vampiros. Se ha reportado que la abundancia del vampiro está positivamente relacionada con la disponibilidad de refugios y alimento (Brown, 1994). En este sentido, la introducción de ganado doméstico podría tener un efecto positivo en la abundancia de las poblaciones de vampiros. Considerando que una de las actividades económicas más importantes de nuestra zona de estudio es la crianza de ganado caprino, pensamos que esta oferta adicional de alimento podría representar otra de las causas del conflicto. De esta forma, tanto la oferta adicional de refugios (los socavones mineros abandonados) como la oferta adicional de alimento (el ganado caprino) estarían incidiendo positivamente en las poblaciones de vampiros. Por otro lado, también se sabe que los ataques no son aleatorios; en un estudio realizado en Costa Rica, Turner (1975) encontró que las reses que pasaban la noche en zonas arboladas y aquellas que se encontraban al borde de los grupos eran las que tenían mayor probabilidad de ser atacadas, lo que muestra que otros factores también podrían estar incidiendo en los ataques al ganado.
La forma de enfrentar el problema por los pobladores se centra en tres actividades principales: destrucción de los potenciales refugios de vampiros, que se realiza dinamitando los socavones abandonados, envenenamiento de todos los murciélagos, sean o no vampiros, con productos altamente peligrosos, como los raticidas; y caza directa de los murciélagos que se acercan a las fuentes de luz de las casas o las que se encuentran cerca de los corrales, lo que se realiza con plantas espinosas (A. Quitón, SENASAG, com. pers., 2006). Estas prácticas afectan poblaciones de otras especies de murciélagos que comparten los refugios con los vampiros o que son confundidos con estos, que en el caso específico de nuestro sitio de estudio implica a murciélagos insectívoros como Histiotus montanus (Verpertilionidae) y Molossus sp. (Molossidae), considerados importantes en el control de poblaciones de insectos, incluyendo aquellos que transmiten enfermedades a humanos (Kalko y Aguirre, 2007).
Nuestro objetivo fue entender cuáles son los patrones de la incidencia de ataques de vampiros en la Prepuna boliviana. Para esto se evaluó si la incidencia de ataques es afectada por la distribución y cantidad del ganado como recurso alimenticio, la distancia de los corrales a los principales refugios de los vampiros, la abundancia relativa del vampiro y algunas características propias del hábitat como la cobertura y altura de la vegetación. Asimismo, discutimos algunos puntos a considerar para una estrategia de manejo del conflicto entre vampiros y ganado doméstico, con sus implicancias sobre otras especies de murciélagos.
MATERIALES Y MÉTODOS
Área de estudio
El trabajo fue realizado en los Municipios de Cotagaita y Tumusla de la Provincia Nor Chichas, departamento de Potosí, Bolivia. La región de la Prepuna boliviana se caracteriza por presentar inviernos fríos, con temperaturas extremas de -13ºC, y veranos con temperatura máximas de ~30ºC, siendo la temperatura promedio anual de 15ºC y la precipitación media anual de 300 mm (López, 2000). La vegetación está dominada por chaparrales espinosos deciduos, en los cuales son frecuentes las cactáceas columnares y las plantas rastreras, con presencia de bosquecillos dominados por especies de Prosopis, Cercidium y Acacia (López, 2000).
Las actividades económicas principales de los pobladores locales son la agricultura y la ganadería de caprinos, aunque también se cría ganado vacuno, porcino y aves de corral. El ganado caprino pasa las noches dentro de corrales con muros construidos con plantas espinosas y durante el día está suelto en las áreas de pastoreo comunales.
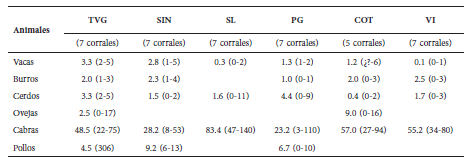
Para el estudio se consideraron seis localidades, separadas por un mínimo de 8 km y un máximo de 25 km, que albergan poblaciones humanas de 50 y 60 familias (Fig. 1). En las localidades de Pampa Grande (PG), Sinandoma (SI), Tambo Valle Grande (TVG) y Vichacla (VI) cada corral está junto a la casa a la que pertenece, mientras que en San Lorenzo (SL) y Cotagaitilla (CO) los corrales se encuentran un poco alejados de las casas en un área común. La cantidad de cabras es siempre mayor respecto a la cantidad de otros animales en todas las localidades (Tabla 1).

Fig. 1. Mapa de la Provincia Nor Chichas en el Departamento de Potosí, Bolivia. Se indican las seis localidades donde se realizó el estudio. Cada localidad es una pequeña población
Tabla 1 Animales criados por las personas en cada una de las localidades estudiadas. En la tabla se presentan los promedios y rango (entre paréntesis) de la cantidad de animales por tipo de ganado. TVG = Tambo Valle Grande; SIN = Sinandoma; SL = San Lorenzo; PG = Pampa Grande; COT = Cotagaitilla; VI = Vichacla.
Métodos de muestreo
El diseño de muestreo consideró dos aproximaciones:
1) La evaluación de la relación entre la incidencia de ataques de vampiro y las características propias de cada localidad: a) la abundancia relativa de vampiros, b) la distancia de la localidad a los refugios de vampiros más cercanos (socavones de minas abandonadas), y c) la altura y la cobertura de la vegetación. Para este propósito, se consideraron todos los ranchos de cada localidad como unidades de evaluación y las localidades como unidades de respuesta.
2) La evaluación de la relación entre la incidencia de ataques y las características propias de cada corral, para lo que se consideraron las siguientes variables: a) número de cabezas de ganado, b) superficie del corral en el cual el ganado descansa por las noches, c) presencia o ausencia de techo en el corral, d) porcentaje del corral que presentaba techo (cobertura de techo), y e) material con el cual fue construido el corral. Para este propósito, cada corral fue tratado como una unidad de respuesta, independientemente de la localidad a la que pertenecía.
Cálculo de la incidencia de ataques al ganado caprino
Para obtener la incidencia de ataques al ganado caprino se revisó a todos los animales de cada corral en busca de heridas nuevas (de la noche anterior), durante 22 días consecutivos simultáneamente en las seis localidades entre febrero y marzo de 2007. Durante esta revisión también se verificó la posible reincidencia de ataques (nuevos ataques sobre cicatrices). Este trabajo fue desarrollado conjuntamente con los dueños de los corrales, quienes contabilizaron las heridas diariamente, luego de un periodo de entrenamiento para uniformar criterios. La incidencia de ataques (I) para cada corral se calculó a partir de la siguiente relación:
I = (No animales con heridas recientes / días de observación) / cantidad ganado
Por otro lado, para cada localidad se calculó la incidencia como el promedio de incidencias obtenidas por corral en dicha localidad. Para comparar esta incidencia entre localidades se utilizó una prueba de Kruskall-Wallis (Siegel y Castellan, 1995).
Análisis de las características ambientales en las localidades
Para este análisis se consideró la incidencia de ataques como dependiente de la abundancia relativa de vampiros para cada localidad. La abundancia relativa se cuantificó en base al esfuerzo de muestreo: número de murciélagos capturados por metro de red/hora (Medellín et al., 2000). Entre febrero y marzo de 2007 (época de lluvias) se capturaron vampiros en cada localidad instalando de forma aleatoria entre 10 y 15 redes de niebla (de 6 x 3 m), dependiendo de la extensión del área de los corrales. Las redes fueron dispuestas a 1 m de la barda de los corrales para garantizar la captura de vampiros y fueron operadas por tres noches consecutivas en cada localidad, entre las 18:30 y 00:30, en una sola ocasión.
Además, se evaluó la distancia de los principales refugios (minas abandonadas) con relación al centro de la localidad (cuando los corrales estaban asociados a las casas) o al área de los corrales (en el caso de que todos los corrales se hallaran ubicados en un área común), en un radio de 10 km. Se eligió esta distancia ya que, aunque estos murciélagos pueden volar hasta 20 km buscando alimento, en general forrajean en áreas de 5 a 8 km desde sus refugios (Greenhall y Schmidt, 1988; Medina et al., 2007). Para esto se utilizaron las coordenadas geográficas obtenidas de los socavones que se encontraban en ese radio de 10 km y, mediante un Sistema de Información Geográfica (Arcview 9.3), se calculó la distancia entre este socavón y el centro de la localidad.
Finalmente, otras dos variables que también se consideraron en este análisis, porque suponíamos que podían afectar la incidencia de los ataques a través de la actividad de los murciélagos, fueron la cobertura y la altura de la vegetación en el área circundante a los corrales. Para evaluar estas variables se contabilizaron todas las plantas con DAP mayor a 1.5 cm en 10 transectos de 50 x 2 m (modificado de Mostacedo y Fredericksen, 2000). Para cada transecto se anotó la altura de cada planta y se calculó la densidad (número de árboles o arbustos / superficie), considerando dos estratos de altura: el arbustivo, plantas < 1 m, y el arbóreo, > 1 m.
Considerando nuestro tamaño de muestra (n = 6 localidades) se decidió calcular correlaciones de Spearman (Siegel y Catellan, 1995) entre la incidencia de ataques y las variables propuestas. En todas las correlaciones se consideró un α = 0.05 y para determinar la magnitud y sentido de la correlación el valor del coeficiente de correlación (rs).
Análisis de las características del manejo del ganado
Para este análisis se consideraron variables obtenidas para cada uno de los corrales. La cantidad de ganado por corral fue considerada como una variable que podría explicar la incidencia de ataques en la región. El número de cabezas de ganado caprino fue obtenido realizando un censo de los animales en cada corral. La superficie del corral también fue considerada como variable independiente, bajo el supuesto de que corrales de mayor tamaño pueden ser localizados más fácilmente por los vampiros. La superficie de cada corral fue estimada utilizando una cinta métrica y de acuerdo a la forma del corral se aplicó la fórmula correspondiente de cálculo de área. Adicionalmente también se estimó el porcentaje del corral que estaba cubierto por techo y el material con el cual se construían las bardas (como barreras al ingreso de vampiros).
Se utilizó un Modelo Lineal Generalizado —GLM por sus siglas en inglés— (Mardia et al., 1979) para determinar las variables que explican mejor la incidencia de ataques. Se consideraron como variables independientes la cantidad de cabezas de ganado caprino por corral, la superficie del corral y el porcentaje de cobertura del techo; y como variables independientes categóricas el material con el cual se construyó la barda del corral (1: corrales construidos con plantas espinosas, 2: corrales construidos con adobe y piedra, y 3: corrales mixtos) y la localidad, que fue considerada como bloque debido a la variación de la incidencia que se registró entre localidades. Previamente a correr el GLM, se realizó un análisis exploratorio para verificar posibles autocorrelaciones entre variables (Underwood, 1997; Gotelli y Ellison, 2004). Para identificar las variables independientes que explican mejor la incidencia de ataques calculada, se realizó una prueba “Stepwise” estándar, que incluye un análisis de eliminación y adición de variables (Gotelli y Ellison, 2004). Se consideró el cambio de valor del criterio de información de Aikaike (AIC) para escoger el mejor modelo y el R2 para evaluar el ajuste de este modelo (Gotelli y Ellison, 2004).
RESULTADOS
En la zona se contabilizaron 2044 cabras, en un total de 40 corrales revisados en las seis localidades estudiadas. En el periodo de evaluación, se registraron 637 ataques de vampiro, de los cuales solo el 7.1% (45 ataques) fueron realizados en cabras que ya habían sido mordidas la noche anterior. Estos resultados dan un promedio de 27 animales atacados por día considerando las seis localidades evaluadas. La incidencia promedio (ataques recientes) fue de 0.031 ± 0.062 animales mordidos/día en cada uno de los corrales. La incidencia de ataques fue distinta entre las seis localidades evaluadas (K-W χ²5 = 25.75; P≤0.05), con la mayor incidencia registrada en Pampa Grande y la menor en Vichacla y Sinandoma (Fig. 2). Las mordidas se dieron con mayor frecuencia detrás de las orejas (50% de los ataques) y en el lomo (35% de los ataques).
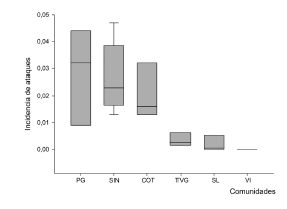
Fig. 2. Incidencia de ataques de vampiros al ganado caprino por localidad estudiada. La gráfica muestra como medida de tendencia central la mediana y las cajas representan los cuartiles. PG = Pampa Grande; SIN = Sinandoma; COT = Cotagaitilla; TVG = Tambo Valle Grande; SL = San Lorenzo; VI = Vichacla.
Análisis de las características ambientales en las localidades
La incidencia de ataques de vampiro al ganado caprino no se correlacionó con las variables de altura de la vegetación arbolada (rs = 0.07; P = 0.89), altura de la vegetación arbustiva (rs = 0.27; P = 0.59), densidad de vegetación baja alrededor de los corrales (rs = -0.63; P = 0.179), ni con la distancia de la localidad al socavón abandonado más cercano (rs = -0.71; P = 0.11). Sin embargo, fue claro que las localidades con mayor abundancia relativa de vampiros fueron las que presentaron mayor incidencia de ataques al ganado (rs = 0.83; N = 6; P = 0.04; Fig. 3).

Fig. 3. Relación entre la abundancia relativa de vampiros y la incidencia de ataques (r2=0.83; N=6; P=0.04).
Por otro lado, la abundancia relativa de vampiros en las localidades disminuyó con la distancia al refugio más cercano (rs = -0.79; N = 6; P = 0.054), lo que sugiere que las localidades con mayores problemas por el ataque de vampiros son aquellas que tienen refugios más cercanos. Es interesante notar que la abundancia relativa de vampiros baja abruptamente en Vichacla y San Lorenzo, que son localidades que están a más de 6 km del refugio más cercano (Fig. 4).

Fig. 4. Relación entre la abundancia de vampiros y la distancia a la mina más cercana (r2 = -0.79; N = 6; P = 0.058). La línea punteada representa la distancia lineal promedio que se ha encontrado que los murciélagos recorren por noche en la zona.
Análisis de las características del manejo del ganado
A través del análisis “Stepwise” estándar del GLM se generaron cuatro modelos alternativos para explicar la incidencia de ataques de vampiros al ganado (Tabla 2). La variable que mejor ayuda a explicar la incidencia de ataques es la cantidad de ganado en cada corral, que varía inversamente con la incidencia (modelo 4 en Tabla 2), lo que indica que la incidencia de ataques disminuye con incrementos en la abundancia del recurso, en este caso las cabras en cada corral (Fig. 5).
Tabla 2 Comparación de los AIC y R2 de cada uno de los modelos generados a través del “Stepwise” estándar para evaluar el efecto de variables ambientales sobre la incidencia de ataques de vampiro (ver MATERIALES Y MÉTODOS).


Fig. 5. Relación negativa del total de ganado con la incidencia de ataques por corral (F = 7.922; P = 0.008). Los puntos que se muestran encerrados en el círculo corresponden a los ranchos de las localidades de Tambo Valle Grande, San Lorenzo y Vichacla, en los que se registraron baja densidad de ganado y bajos niveles de incidencia de ataques.
No se halló correlación entre el porcentaje de cobertura del techo en los corrales y la incidencia de ataques de vampiros. Sin embargo, es importante notar que los dos corrales cubiertos totalmente tuvieron incidencias muy bajas (entre 3 y 10 veces menores que el promedio para la región).
DISCUSIÓN
La incidencia de ataques de vampiros fue distinta entre las localidades estudiadas: aquellas con mayor incidencia eran las que se encontraban a distancias menores a 6 km de los refugios potenciales (socavones abandonados). A su vez, las localidades con mayor incidencia de ataques fueron también las que presentaron mayores abundancias relativas de vampiros, coincidentemente con lo reportado por Gonzales y Mitchell (1976) para México. Sin embargo, la mayor incidencia no se dio en las localidades que tenían mayor cantidad de ganado (principal recurso alimenticio para los vampiros). Esto sugiere que el factor limitante no es la disponibilidad de alimento (ganado), sino la disponibilidad de refugios cercanos al recurso (los socavones), que albergan una cantidad limitada de vampiros.
Por otro lado, los valores de incidencia obtenidos en las diferentes localidades fueron en general bajos. La localidad con la incidencia más alta fue Pampa Grande, con un animal mordido cada 7 o 9 días por corral. Mientras que en localidades con incidencias menores se observó un animal mordido cada 30 días por corral. Estos resultados sugieren que la cantidad de animales mordidos no es muy alta, en comparación con los reportes de Gonzales y Mitchell (1976), quienes registraron 93 915 reses mordidas de un total de 164 834 animales evaluados en un periodo de dos años en Nicaragua. Adicionalmente, en la zona de estudio los ataques a personas no son comunes, a diferencia de lo que ocurre en las regiones amazónicas (i.e. Schneider et al., 2001; Schneider et al., 2009; Carvalho-Costa et al., 2012; Romero- Sandóval et al., 2013). Pese a esto, en el área de estudio los ganaderos responden con prácticas inadecuadas y no selectivas de control de las poblaciones de vampiros, afectando sobre todo a las poblaciones de murciélagos insectívoros, que se abastecen cerca de las fuentes de luz artificiales asociadas a viviendas humanas o viven en casas abandonadas.
La presencia de techos parciales no impide el acceso de los vampiros a los corrales, sin embargo la incidencia de ataques sí se reduce con una cobertura de techo total; las incidencias en los corrales cubiertos totalmente, fueron las más bajas en comparación a los otros corrales. Esto sugiere que probablemente es suficiente un hueco en los techos parciales para permitir el ingreso de los vampiros.
La reincidencia de los vampiros sobre las presas está reportada en la literatura (Brown, 1994) y esto ha sido utilizado como base para los protocolos de control del vampiro. Por ejemplo, se sugiere colocar el vampiricida en la herida de los animales mordidos previamente. Sin embargo, durante el trabajo se registró un número muy bajo de animales que hubieran sido mordidos varias noches consecutivas (65 de 637 animales atacados). Esto sugiere que, al menos en el área de estudio, el colocar la pomada vampiricida en un animal mordido no incrementa la probabilidad de que el vampiro lo ingiera debido a que la probabilidad de que reincida sobre esa presa es baja.
Nuestra observación de que la incidencia de ataques es mayor en corrales con menor cantidad de ganado parece contraintuitiva, ya que en el norte argentino se ha reportado que la abundancia de murciélagos se duplicaba en zonas ganaderas (Delpietro et al., 1992). Sin embargo, es posible que la explicación sea que existe un número constante de vampiros que se abastece en cada localidad y que estos se repartirán entre las presas disponibles cerca de su refugio. Esta hipótesis está apoyada por dos resultados: la correlación positiva entre la incidencia de ataques al ganado con la abundancia relativa de vampiros, que implicaría que el número de ataques registrados en las localidades está relacionado directamente con el número de vampiros que se encuentran en cada localidad; y que la abundancia de vampiros está inversamente relacionada con el número de cabezas de ganado presente en las localidades estudiadas. Bajo este escenario podemos especular que, en la Prepuna boliviana, las poblaciones de vampiros no están respondiendo directamente a la disponibilidad de recursos alimenticios, sino a la presencia de refugios cercanos a sus áreas de abastecimiento y que son los refugios el factor limitante del número de individuos en las poblaciones de vampiros y por lo tanto de la incidencia de ataques de los mismos sobre el ganado. Entonces, un incremento en la abundancia de cabras debería reducir la probabilidad de ataque para cada cabra, coincidiendo con el efecto de dilución de la probabilidad de ataque per capita con el incremento en la densidad de presas (Davies et al., 2012).Un dato adicional apoya nuevamente nuestra hipótesis. Se sabe que los vampiros en general forrajean en un rango de 6 a 8 km (Wimsatt, 1969). Un estudio de radiotelemetría, realizado en la comunidad de Sinandoma con cinco individuos de D. rotundus, indicó que la distancia máxima que los vampiros se alejan de su refugio en busca de alimento es 5.2 km y que tienden a ser fieles a sus refugios y a sus áreas de forrajeo (I. Moya, datos no publicados). Con base en estos resultados, es muy ilustrativo que las localidades de Pampa Grande, Sinandoma y Cotagaitilla, que tienen los mayores valores de incidencia, se encuentren a menos de 6 km de distancia de los potenciales refugios, y que la incidencia decaiga abruptamente en las otras localidades, que se encuentran a distancias mayores de los refugios.
Por otro lado, de acuerdo a nuestros resultados, en general un animal es atacado sólo por un vampiro cada noche, a diferencia de lo que ocurre en Costa Rica, donde cada animal puede presentar hasta 10 heridas por noche (Turner, 1975). Si la cantidad de ganado se mantiene constante, es posible postular que la incidencia de ataques reflejaría de forma confiable la abundancia relativa de vampiros que se encuentran en nuestro sitio de estudio. Para hacer estas suposiciones nos hemos basado en que los vampiros son altamente eficientes para alimentarse y no requieren más de 15 o 20 minutos para obtener el alimento necesario (Greenhall y Schmidt, 1988; Wimsatt, 1969), por lo cual no necesitan visitar más de una cabra por noche. De esta forma, la incidencia de ataques puede ser un método confiable de monitoreo de la abundancia relativa de vampiros y puede ayudar a determinar el momento adecuado para realizar una campaña de control del vampiro. Sin embargo, también debemos considerar que se han reportado situaciones en las que varios murciélagos se alimentaban de una misma herida (Greenhall et al., 1971), lo que podría resultar en una subestimación de la abundancia de vampiros. Este aspecto debería ser considerado en los programas de prevención que se realicen recomendando siempre que las acciones deberían realizarse mucho antes de que los conflictos se inicien.
En el mismo escenario, si se acepta que la abundancia de refugios es el factor limitante de las poblaciones de vampiros, un incremento en la actividad minera reactivando los socavones abandonados o abriendo otros nuevos, implicaría a la larga un incremento en las poblaciones de vampiros, una vez que estos sean abandonados, lo que tendrá como consecuencia un aumento en la incidencia de ataques al ganado en la zona. Por lo tanto, una de nuestras recomendaciones respecto a esta situación es que para el manejo del conflicto se recomiende o incluso se obligue a cerrar los socavones que van a ser abandonados, ya que en esta zona no se han registrado otras especies compartiendo los refugios con D. rotundus, por lo que no habría un efecto sobre las otras especies de murciélagos. Otra recomendación que podemos extraer de nuestros datos es generar programas de control letal mucho más efectivos aplicando la warfarina a los vampiros capturados en sus refugios. Sin embargo, es importante reconocer que para obtener resultados positivos a largo plazo, estos controles deben ser repetidos anualmente (Conover, 2001). En el caso de los vampiros, si bien se ha llegado a estimar una reducción de hasta el 95% de los individuos de una población luego de una campaña de control (Flores, 2003), aspectos como su capacidad reproductiva, cuidado parental y capacidad de vuelo (como mecanismo de repoblamiento) deben ser tomados en cuenta al momento de determinar los intervalos y la intensidad de la intervención. Es importante destacar que si solo se emplea el control letal, las poblaciones de vampiros podrían restablecerse por efecto de la migración (habiendo en la zona refugios libres y alimento a disposición); y si solo se cierran los socavones, los vampiros podrían buscar otros refugios por la misma zona, incluyendo los entretechos de las casas, lo que incrementaría los conflictos con los humanos. Todos estos impactos van a tener aún consecuencias en la economía de los pobladores locales llevándolos a mantener sus prácticas no selectivas de control del vampiro.
También deben ser consideradas las campañas de vacunación del ganado contra la rabia que deberían incluirse dentro de una estrategia integral, asegurando de esta forma la sanidad animal en la zona. En este sentido, se sabe que la vacunación es el mejor método para evitar brotes de rabia (Anderson et al., 2012), sin embargo si no está acompañada de campañas de control letal del vampiro aún es posible que se mantengan otros problemas, como las heridas en los animales susceptibles a infecciones o los problemas de salud relacionados a la pérdida de peso y reducción en la producción de leche por parte del ganado (Acha y Málaga Alba, 1988). Por todo lo expuesto, lo más apropiado sería realizar todas las acciones simultáneamente, generando programas integrales que permitan asegurar la salud humana y animal a través de campañas de vacunación antirrábica y manteniendo las poblaciones de vampiro en niveles bajos previniendo posibles conflictos con los pobladores locales.
Finalmente, sería necesario establecer conjuntamente a estas actividades un programa de capacitación y educación, dirigido tanto a los técnicos de las diferentes instituciones que trabajan en la zona en temas relacionados a la producción y sanidad animal, como a la gente de las diferentes localidades, con el objetivo de generar conciencia sobre cuán importante será poner en práctica una estrategia conjunta de control del vampiro, prevención de la rabia en la zona y conservación de las otras especies de murciélagos.
AGRADECIMIENTOS
Agradecemos al Postgrado en Ecología y Conservación de la Universidad Mayor de San Andrés y a la Organización Panamericana de la Salud por haber apoyado financieramente la realización del proyecto. Al Programa para la Conservación de los Murciélagos de Bolivia por el apoyo logístico y a sus miembros que participaron en el trabajo de campo (Raquel Galeón, Oswaldo Palabral, Sandra Rivera). Al Servicio Nacional de Sanidad Agropecuaria e Inocuidad Animal (SENASAG) por el soporte logístico en campo y a Idea Wild por haber apoyado con equipo. También agradecemos a María Sandoval y a los revisores anónimos del manuscrito cuyos aportes mejoraron sustancialmente la interpretación de los resultados. Finalmente también queremos extender nuestro agradecimiento a cada una de las localidades por habernos permitido realizar el trabajo.
LITERATURA CITADA
1. ACHA PN y A MÁLAGA ALBA. 1988. Economic losses due to Desmodus rotundus. Pp. 208–214, en: Natural History of Vampire Bats (AM Greenhall and U Schmidt, eds.). CRC Press, Boca Raton, FL.
2. AGUIRRE LF. 2007. Historia natural, distribución y conservación de los murciélagos de Bolivia. Centro de Ecología y Difusión Simón I. Patiño, Santa Cruz de la Sierra. [ Links ]
3. ALLEN LR y EC SPARKES. 2001. The effect of dingo control on sheep and beef cattle in Queensland. Journal of Applied Ecology 38:76-87. [ Links ]
4. BADGER DD y KM SCHMIDT. 1979. Evaluation of the vampire bat control program. Departament of Agricultural Economics, Oklahoma State University, Oklahoma. [ Links ]
5. BARCLAY RMR y LD HARDER. 2003. Life histories of bats: Bats in the slow lane. Pp. 209-246, en: Bat Ecology (TH Kunz y MB Fenton, eds.). The University of Chicago Press, Chicago and London. [ Links ]
6. BECKMANN JP, CW LACKEY y J BERGER. 2004. Evaluation of deterrent techniques and dogs to alter behavior of “nuisance” black bears. Wildlife Society Bulletin 32(4):1141-1146.
7. BELOTTO AJ. 2001. Raiva transmitida por morcegos nas Américas: impacto na saúde pública e na produção. Pp. 24-25, en: Seminario internacional de murciélagos como transmisores de rabia. São Paulo, SP. Programas e resumos. Secretaria da Saúde, São Paulo. [ Links ]
8. BROWN DE. 1994. The vampire bat in fact and fantasy. High-Lonesome Books, Silver City, N. Mexico D.F. [ Links ]
9. CARVALHO-COSTA FA, VL TEDESQUI, M DE JESUS NASCIMENTO MONTEIRO y MN BÓIA. 2012. Outbreaks of attacks by hematophagous bats in isolated riverine communities in the Brazilian Amazon; A challenge to rabies control. Zoonoses and Public Health 59:272-277. [ Links ]
10. CONOVER MR. 2001. Resolving human-wildlife conflicts: The science of wildlife damage management. Ed. Lewis Publishers, Boca Ratón, Florida. [ Links ]
11. CONSTANTINE DG. 2009. Bat rabies and other Lyssavirus infections. Reston, Va. U.S. Geological Survey Circular 1329, 68 p. [ Links ]
12. DAVIES NB, JR KREBS y SA WEST. 2012. An introduction to behavioural ecology. Blackwell Scientific Publications, Oxford. [ Links ]
13. DELPIETRO HA, N MARCHEVSKY y E SIMONETTI. 1992. Relative population densities and predation of the common vampire bat (Desmodus rotundus) in natural and cattle-raising areas in north-east Argentina. Preventive Veterinary Medicine 12:13-20. [ Links ]
14. EL POTOSÍ. 2006. “Vampiros” atacan a Ganado caprino y bovino en Cotagaita. http://www.elpotosi.net.
15. FLORES R. 2003. Técnicas, substancias y estrategias para el control del murciélago vampiro. Organización Panamericana de la Salud, ISBN970-92109-1-2. México D.F. [ Links ]
16. GONZALES R y GC MITCHELL. 1976. Vampire bat control programs in Latin America. Proceedings of the 7th Vertebrate Pest Conference. Paper 22. http://digitalcommons.unl.edu/vpc7/22. [ Links ]
17. GOTELLI N y AM ELLISON. 2004. A primer of ecological statistics. Sinauer Associates Inc., Sunderland MA. 510 p. [ Links ]
18. GREENHALL AM y U SCHMIDT. 1988. Natural history of vampire bats. Bocca Raton, CRC Press, Florida. [ Links ]
19. GREENHALL AM y U SCHMIDT. 1971. Attacking behavior of the vampire bat, under field conditions in Mexico. Biotropica 3 (2): 136-141. [ Links ]
20. JOHNSON N, A VOS, C FREULING, N TORDO, AR FOOKS y T MÜLLER. 2010. Human rabies due to Lyssavirus infection of bat origin. Veterinary Microbiology 142:151-159. [ Links ]
21. KALKO EKV y LF AGUIRRE. 2007. Insectivoría, en: Historia natural, distribución y conservación de los murciélagos de Bolivia (LF Aguirre, ed.). Centro de Ecología y Difusión Simón I. Patiño, Santa Cruz de la Sierra. [ Links ]
22. LA PATRIA. 2006. Vampiros chupan sangre a cabras y vacas en Cotagaita. www.enlared.org.bo [ Links ]
23. LA RAZÓN. 2006. Murciélagos invaden Vitichi y Cotagaita. www.la-razon.com [ Links ]
24. LOPEZ R. 2000. La Prepuna boliviana. Ecología en Bolivia 34:45-65. [ Links ]
25. MARDIA KV, JT KENT y JM BIBBY. 1979. Multivariate Analysis. Academic Press. ISBN 0-12-471252-5. [ Links ]
26. MAYEN F. 2003. Haematophagous bats in Brazil, their role in rabies transmission, impact on public health, livestock industry and alternatives to an indiscriminate reduction of bat population. Journal of Veterinary, Series B 50(10):469-472. [ Links ]
27. MEDELLÍN RA, M EQUIHUA y M AMIN. 2000. Diversity and abundance as indicators of disturbance in Neotropical Rainforests. Conservation Biology 6:1666-1675. [ Links ]
28. MEDINA A, CA HARVEY, DS MERLO, S VÍLCHEZ y B HERNÁNDEZ. 2007. Bat diversity and movement in an agricultural landscape in Matiguás, Nicaragua. Biotropica 39:120-128. [ Links ]
29. MESSMER TA. 2000. The emergence of human-wildlife conflict management: Turning challenges into opportunities. International Biodeterioration and Biodegradation 45:97-102. [ Links ]
30. MMAyA (Ministerio de Medio Ambiente y Agua). 2009. Libro rojo de la fauna silvestre de vertebrados de Bolivia. Ministerio de Medio Ambiente y Agua, La Paz. [ Links ]
31. MOSTACEDO B y T FREDERICKSEN. 2000. Manual de métodos básicos de muestreo y análisis en ecología vegetal. BOLFOR. Santa Cruz de la Sierra. [ Links ]
32. NÚÑEZ C, A PÁEZ, C HERNÁNDEZ, H ESCOBAR y A BONELO. 2012. Transmisión del virus de la rabia entre murciélagos urbanos del departamento del Valle del Cauca, Colombia, 1999-2008. Infectio 16(1):23-29. [ Links ]
33. ODUM EP. 1971. Fundamentals of ecology. WB. Saunders, Filadelfia. [ Links ]
34. OJASTI J y F DALLMEIER (Eds). 2000. Manejo de fauna silvestre neotropical. SI/MABSeries # 5. Smithsonian Institution / MAB Biodiversity Program, Washington D.C. [ Links ]
35. OPPENHEIMER JR y GE LANGE. 1969. Cebus monkeys: effects on branching of Gustavia trees. Science 165:187-188. [ Links ]
36. PÉREZ E y LF PACHECO. 2006. Damage by large mammals to subsistence crops in a montane forest of Bolivia. Crop Protection 25:933-939. [ Links ]
37. PACHECO LF, G GALLARDO, A NUÑEZ y F VARELA. 2010. Puma-livestock conflict in the Bolivian highlands: Lessons learned. Wild Felid Monitor 3(2):19. [ Links ]
38. ROMAÑACH SS, PA LINDSEY y R WOODROFFE. 2007. Determinants of attitudes towards predators in central Kenya and suggestions for increasing tolerance in livestock dominated landscapes. Oryx 41(2):185-195. [ Links ]
39. ROMERO-SANDOVAL, C PARRA, G GALLEGOS, A GUANOPATÍN, MF CAMPAÑA, M HARO, S CALAPAQUI, C MORETA, F VITERI, M FEIJOO-CID y M MARTIN. 2013. Hematophagous bat bites in Ecuadorian Amazon: Characterization and implications for sylvatic rabies prevention. Public Health Action 3:85-89. [ Links ]
40. SIEGEL S y J CASTELLAN. 1995. Estadística no paramétrica. Editorial Trillas, México D.F. [ Links ]
41. SCHNEIDER MC, J ARON, C SANTOS-BURGOA, W UIEDA y S. RUIZ-VELAZCO. 2001. Common vampire bat attacks on humans in a village of the Amazon region of Brazil. Cadernos de Saúde Pública, 17:1531-1536. [ Links ]
42. SCHNEIDER MC, PC ROMIJIN, W UIEDA, H TAMAYO, DF Da SILVA, A BELOTTO, JB Da SILVA y LF LEANERS. 2009. Rabies transmitted by vampire bats to humans: An emerging zoonotic disease in Latin America? Revista Panamericana de Salud Pública. 25:260-269. [ Links ]
43. TURNER DC. 1975. The vampire bat – A field study in behavior and ecology. John Hopkins University Press, Baltimore and Londres.
44. UNDERWOOD AJ. 1997. Experiments in ecology: Their logical design and interpretation using analysis of variance. Cambridge University, Nueva York. [ Links ]
45. ZARCO-GONZÁLEZ MM, O MONROY-VILCHIS, C RODRÍGUEZ-SOTO y V URIOS. 2012. Spatial factors and management associated with livestock predations by Puma concolor in Central Mexico. Human Ecology 40:631-638. [ Links ]
46. WIMSATT WA. 1969. Transient behavior, nocturnal, activity patterns and feeding efficiency of vampire bats (Desmodus rotundus) under natural conditions. Journal of Mammalogy 5:233-244. [ Links ]

