










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkMastozoología neotropical
versão impressa ISSN 0327-9383versão On-line ISSN 1666-0536
Mastozool. neotrop. vol.22 no.2 Mendoza dez. 2015
NOTA
Observaciones del nado de un grupo de monos aulladores (Alouatta palliata) en el Océano Pacífico de Costa Rica
Hansel Herrera1, Sergio Escobar-Lasso1,2, Héctor González-Ancín1 y Marie-Cécile Béal3
1 Programa de Posgrado en Conservación y Manejo de Vida Silvestre, Instituto Internacional en Conservación y Manejo de Vida Silvestre. ICOMVIS. Heredia, Costa Rica [Correspondencia: Hansel Herrera <bluecloudcr@gmail.com>.
2 Fundación R.A.N.A. (Restauración de Ambientes Neotropicales Alterados), Heredia, Costa Rica.
3 Instituto de Oceanología de Costa Rica, Guanacaste, Costa Rica.
RESUMEN.
Se han observado individuos solitarios de mono aullador Alouatta palliata nadando en bosques inundables, pantanos, lagunas y ríos. Sin embargo, dicho evento nunca había sido observado en grupos de individuos ni tampoco en el mar. Por lo tanto, el objetivo de la presente nota es reportar por primera vez la observación de una tropa de A. palliata nadando del continente hacia una isla cercana en el Pacífico de Costa Rica.
ABSTRACT.
Observations on swimming in a group of mantled howler monkeys (Alouatta palliata) in the Pacific Ocean of Costa Rica.
Mantled howler monkeys, Alouatta palliata, have been observed swimming in flooded forests, swamps, lagoons and rivers. This type of event had only been documented on solitary individuals in fresh water bodies so far. Therefore, the purpose of this note is to report, for the first time, a troop of A. palliata swimming from the continent to a nearby island in the Pacific coast of Costa Rica.
Palabras clave: Comportamiento social; Desplazamiento acuático; Dispersión oceánica; Movimiento animal en grupo; Primates neotropicales.
Key words: Animal group movement; Aquatic movement; Neotropical primates; Ocean dispersal; Social behavior.
Recibido 4 enero 2015.
Aceptado 20 mayo 2015.
Editor asociado: M Kowalewski
Los homínidos son el único grupo de primates que normalmente utilizan el agua como medio de transporte o alimentación (Williamson et al., 1988). No obstante, la capacidad de nadar ha sido documentada en varios primates del Viejo Mundo, como el macaco coronado (Macaca radiata), el mono narigudo (Nasalis larvatus), el papión oliva (Papio anubis) y el macaco japonés (Macaca fuscata) (Wata, 1981; Bennett, 1988; Bennet y Sebastian, 1988; Yeager, 1991; Forthman, 2000; Agoramoorthy et al., 2000). Entre los platirrinos (primates del Nuevo Mundo), únicamente los monos aulladores (Alouatta spp.) y los monos araña (Ateles geoffroyi) han mostrado esta conducta (Chaves y Stoner, 2010; González-Socoloske y Snarr, 2010). Sin embargo, el mono chucuto (Cacajao melanocephalus) ha sido visto nadando específicamente después de caer al agua de manera accidental y su reacción inmediata fue trepar al árbol más cercano (Bezerra et al., 2011).
En todos los casos reportados, el nado en los primates neotropicales ha sido realizado en cuerpos de agua dulce, como ríos y pequeños lagos, principalmente por individuos solitarios (González-Socoloske y Snarr, 2010). Existe un único reporte de un grupo de tres individuos de A. geoffroyi atravesando un río en el sur de México (Chaves y Stoner, 2010). El nado en primates neotropicales se ha asociado a causas antropogénicas como la degradación del hábitat y la cacería, así como a la búsqueda de mejores fuentes de alimentación y al escape de los depredadores (Matsuda et al., 2008; Chaves y Stoner, 2010; González-Socoloske y Snarr, 2010; Yeager, 1991). A pesar de los reportes existentes, nunca antes se había documentado el desplazamiento acuático de un primate neotropical desde el continente hacia una isla marina. El objetivo de la presente nota es reportar por primera vez la observación de una tropa de A. palliata nadando del continente hacia una isla cercana en el océano Pacífico de Costa Rica.
Las observaciones del nado fueron hechas en el área marina entre Isla Plata (10°26’51.63” N; 85°48’11.75” O) y Punta Salinas (10°26’34.01” N; 85°47’39.90” O), en el distrito de Cabo Velas, Guanacaste, Costa Rica (Fig. 1). Dicha área se clasifica como bosque seco tropical y presenta dos estaciones bien definidas (sensu Holdridge, 1982; Janzen, 1983). La estación seca se extiende de diciembre a mayo y la lluviosa de junio a noviembre. La precipitación media anual es de 900-2400 mm (Janzen, 1983).
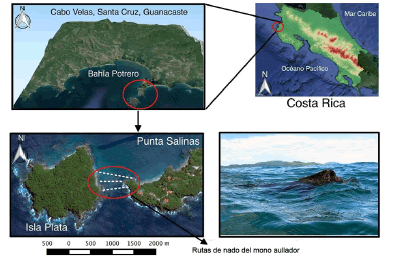
Fig. 1. Ubicación geográfica del sitio de observación del mono aullador A. palliata nadando en la costa pacífica de Costa Rica. Las líneas blancas punteadas representan las rutas de nado de los monos aulladores. Imágenes satelitales tomadas de Google Earth.
Punta Salinas es una extensión continental de 1.5 km, la cual alberga algunas casas de veraneo, pequeños hoteles y complejos turísticos. La Isla Plata se encuentra separada de punta Salinas por unos 170 m lineales de mar (Fig. 1). Dicha isla tiene una extensión de 19 ha y se encuentra totalmente deshabitada (IOC, 2013). El bosque presente en la isla está conformado por dos estratos: un dosel compuesto por árboles entre 15-25 m de alto y un sotobosque denso formado por hierbas y arbustos entre 3-5 m de alto (IOC, 2013).
Los monos aulladores fueron observados nadando el 2 de marzo de 2013 a las 14:00 h por el equipo de investigación del Instituto de Oceanología de Costa Rica (IdO) mientras navegaban en bote hacia la Isla Plata (IOC, 2013). Los investigadores observaron una tropa de quince individuos nadar de Punta Salinas a Isla Plata (Fig. 2), los cuales fueron fotografiados con una cámara GoPro Hero 4. La tropa tardó 35 minutos en atravesar la distancia entre Punta Salinas e Isla Plata (170 m aprox.) (Fig. 2).

Fig. 2. Mono aullador A. palliata nadando de Punta Salinas (área continental) a la Isla Plata en la costa pacífica de Costa Rica, Guanacaste. Fotografía de Andrés Jiménez Orozco.
Durante expediciones ocasionales realizadas por los investigadores del IdO entre febrero de 2013 y diciembre de 2014 se pudo corroborar la presencia de monos aulladores en la Isla Plata. Además, en 2006 Tandy (2015) observó un grupo de A. palliata en la isla. Lo anterior sugiere que dentro de Isla Plata podría haber una población residente de A. palliata, lo cual debe corroborarse con futuras investigaciones (IOC, 2013; Tandy, 2015).
El desplazamiento a través de cuerpos de agua es un acto inusual en los primates neotropicales (Chaves y Stoner, 2010). Los monos aulladores (Alouatta spp.), especialmente A. seniculus, A. palliata y A. pigra, son los que más muestran esta conducta (Chaves y Stoner, 2010; González-Socoloske y Snarr, 2010; Izawa y Lozano, 1990). En los platirrinos, se han documentado siete eventos de nado de los cuales cinco corresponden a Alouatta spp. (Collins y Southwick, 1952; Neville et al., 1988; Izawa y Lozano, 1990; Kinzey, 1997; Chaves y Stoner, 2010; González-Socoloske y Snarr, 2010; Bezerra et al., 2011).
Se han observado individuos solitarios de monos aulladores nadando en bosques inundables, pantanos, lagunas y ríos, pero nunca en el mar (Collins y Southwick, 1952; Neville et al., 1988; Kinzey, 1997; González-Socoloske y Snarr, 2010). Por lo tanto, nuestras observaciones representan el primer registro del nado en el mar para el género Alouatta y posiblemente para los primates neotropicales. Además, nuestras observaciones también constituyen el primer registro de un grupo de monos aulladores atravesando un cuerpo de agua, ya que todos los registros anteriores corresponden a individuos solitarios.
El estilo de nado de los monos observados se caracterizó por movimientos alternos de las cuatro extremidades (similar al de un perro en el agua), con la cabeza a flote y el resto del cuerpo sumergido en el agua (Fig. 2). Esta forma de nado es consistente con la reportada en otros estudios sobre el género Alouatta y con los testimonios de algunas personas en las distintas localidades donde se ha observado esta conducta (Izawa y Lozano, 1990; Chaves y Stoner, 2010; González-Socoloske y Snarr, 2010).
De los primates neotropicales, los monos aulladores son los que tienen una mayor capacidad de dispersión en hábitats fragmentados (Kinzey, 1997; Bicca-Marques, 2003; Estrada et al., 2006). Alouatta palliata ha sido observado moviéndose a través de carreteras, cercas vivas, bosques de galería, pastizales, cultivos, áreas semiurbanas y cuerpos de agua (Neville et al., 1988; Kinzey, 1997; Asensio et al., 2009; González-Socoloske y Snarr, 2010). La conducta de nado en los monos aulladores demuestra la flexibilidad que tienen para sobrevivir en hábitats fragmentados y la capacidad para dispersarse o movilizarse en busca de mejores fuentes de alimentación (Bicca-Marques, 2003; González-Socoloske y Snarr, 2010).
El papel de los ríos como barreras para la dispersión y la variabilidad genética de las especies y las comunidades ha sido estudiado anteriormente (Gascon et al., 2000). En los platirrinos, el efecto limitante de los cuerpos de agua ha sido investigado por Peres et al. (1996) y Collins y Dubach (2000). El acto de nado en el mar aquí documentado indica que este medio no representa en sí mismo una barrera para los monos aulladores. Sin embargo, es posible que la longitud del cuerpo de agua sea un factor determinante para la dispersión de A. palliata en este medio (González-Socoloske y Snarr, 2010). Las observaciones del nado en los monos aulladores muestran que la longitud de los cuerpos de agua que atraviesan es relativamente corta. Por ejemplo, se han observado distancias máximas de nado de 200 m en A. seniculus (Izawa y Lozano, 1990), 88 m en A. pigra (Chaves y Stoner, 2010), 150 m en A. palliata (González-Socoloske y Snarr, 2010) y el presente registro de 170 m en A. palliata (este estudio). Por lo tanto, se podría esperar que los cuerpos de agua que superen considerablemente los 200 m podrían constituir barreras para la dispersión de Alouatta spp. Sin embargo, es necesario contar con un mayor número de observaciones de campo para verificar lo anterior.
Tanto la distancia como la turbulencia del agua han sido propuestos como mecanismos reguladores de los desplazamientos de los monos aulladores en los ríos y lagos (Chaves y Stoner, 2010). El paso marino entre Isla Plata y Punta Salinas (continente) presenta fuertes corrientes oceánicas (Obs. pers.) y se constituye como un paso dificultoso para los humanos y probablemente también para los monos aulladores y otras especies menores. No obstante, nuestra observación evidencia la capacidad de A. palliata de nadar en aguas turbulentas, con presencia de fuertes corrientes y bordeadas por rocas de difícil acceso.
La capacidad de los primates de seleccionar sectores del río que tienen un ancho menor para ser utilizados como pasos ha sido estudiada por Madsuda et al. (2008). Esta destreza se relaciona con una mayor capacidad cognitiva en los primates mayores (y en algunos primates neotropicales como el mono araña y el aullador) y la capacidad de estos de construir mapas mentales (Madsuda et al., 2008; Garber et al., 2009). Es posible que esta conducta sea una estrategia evasiva contra los depredadores acuáticos, como los cocodrilos y los tiburones, y una forma de minimizar el riesgo de ahogamiento (Chaves y Stoner, 2010). En esta ocasión, los monos aulladores escogieron la ruta más corta entre el continente y la Isla Plata (ver Fig. 1).
El desplazamiento acuático de A. palliata aquí documentado es un ejemplo de coordinación grupal (Fichtel et al., 2011) cuyas causas desconocemos. No obstante, este evento demuestra la alta cohesión del grupo y el riesgo que la tropa está dispuesta a asumir con tal de llegar a la isla. Es importante realizar una investigación sistemática y profunda que permita comprender las causas reales de esta inusual conducta. Así mismo, recomendamos estudiar la dinámica poblacional y el comportamiento de los individuos con el fin de comprender cómo la fragmentación del paisaje y otras perturbaciones humanas afectan a los monos aulladores de la zona.
Agradecimientos.
Agradecemos al Instituto de Oceanología de Cosa Rica por brindar todo el apoyo logístico para realizar la presente investigación, a Andrés Jiménez Orozco por compartir sus fotografías con nosotros, a Elpis Joan Chávez Calderón por revisar el manuscrito en forma y contenido y al gobierno de los Estados Unidos por otorgarnos una beca para cursar los estudios de posgrado en Costa Rica.
LITERATURA CITADA
1. AGORAMOORTHY G, I SMALLEGANGE, I SPRUIT y MJ HSU. 2000. Swimming behaviour among Bonnet macaques in Tamil Nadu: A preliminary description of a new phenomenon in India. Folia Primatologica 71:152-153. [ Links ]
2. ASENSIO N, V ARROYO-RODRÍGUEZ, JC DUNN y J CRISTÓBAL-AZKARATE. 2009. Conservation value of landscape supplementation for Howler monkeys living in forest patches. Biotropica 41:768-773. [ Links ]
3. BENNETT E. 1988. Proboscis monkeys and their swamp forests in Sarawak. Oryx 22:69-74. [ Links ]
4. BENNETT E y AC SEBASTIAN. 1988. Social organization and ecology of Proboscis monkeys (Nasalis larvatus) in mixed coastal forest in Sarawak. International Journal of Primatology 9:233-255. [ Links ]
5. BEZERRA BM, AA BARNETT, A SOUTO y G JONES. 2011. Ethogram and natural history of Golden-backed uakaries (Cacajao melanocephalus). International Journal of Primatology 32:46-68. [ Links ]
6. BICCA-MARQUES JC. 2003. How do Howler monkeys cope with habitat fragmentation? Pp. 283-303, en: Primates in fragments: Ecology in conservation (LK Marsh, ed.). Springer Press, New York. [ Links ]
7. CHAVES OM y KE STONER. 2010. River crossing by Ateles geoffroyi and Alouatta pigra in southern Mexico: A preliminary report. Revista Chilena de Historia Natural 83:435-442. [ Links ]
8. COLLINS AC y JM DUBACH. 2000. Biogeographic and ecological forces responsible for speciation in Ateles. International Journal of Primatology 21:421-444. [ Links ]
9. COLLLINS N y C SOUTHWICK. 1952. A field study of population density and social organization in Howling monkeys. Proceedings of the American Philosophical Society 96:143-156. [ Links ]
10. ESTRADA A, J SAENZ, C HARVEY, E NARANJO, D MUÑOZ y M ROSALES-MEDA. 2006. Primates in agroecosystems: Conservation value of some agricultural practices in Mesoamerican landscapes. Pp. 437-470, en: New Perspectives in the Study of Mesoamerican Primates (A Estrada, PA Garber, MS Pavelka y L Luecke, eds.). Springer Press, New York. [ Links ]
11. FORTHMAN D. 2000. Note on water ‘play’ by an Olive baboon Papio anubis in Gilgil, Kenya. African Primates 4:74.
12. FICHTEL C, L PYRITZ y PM KAPPELER. 2011. Coordination of Group Movements in Non-human Primates. Pp. 37-56, en: Coordination in Human and Primate Groups. Springer-Verlag, Heidelberg. [ Links ]
13. GASCON C, JR MALCOLM, JL PATTON, MNF DA SILVA, JP BOGART, SC LOUGHEED, CA PERES, S NECKEL y PT BOAG. 2000. Riverine barriers and the geographic distribution of Amazonian species. Proceedings of the National Academy of Sciences 25:13672-13677. [ Links ]
14. GARBER PA , JC BICCA-MARQU E S y MA AZEVEDOLOPES. 2009. Primate cognition: Integrating social and ecological information in decision making. Pp. 365-385, en: South American primates: Comparative perspectives in the study of behavior, ecology, and conservation (PA Garber, A Estrada, JC Bicca-Marques, EW Heymann y K Strier, eds.). Springer Press, New York. [ Links ]
15. GONZÁLEZ-SOCOLOSKE D y KA SNARR. 2010. An Incident of Swimming in a Large River by a Mantled Howling Monkey (Alouatta palliata) on the North Coast of Honduras. Neotropical Primates 17:28-31. [ Links ]
16. HOLDRIDGE LR. 1982. Ecología basada en zonas de vida. IICA, San José, Costa Rica. [ Links ]
17. INSTITUTO DE OCEANOLOGÍA DE COSTA RICA (IOC). 2013. Evaluación ecológica rápida. Isla Plata. Cabo Velas, Santa Cruz de Guanacaste, Costa Rica. [ Links ]
18. IZAWA K y MH LOZANO. 1990. River crossing by a wild Howler monkey (Alouatta seniculus). Field studies of New World monkeys, La Macarena, Colombia 3:29-33. [ Links ]
19. JANZEN D. 1983. Costa Rican natural history. Chicago: The University of Chicago. [ Links ]
20. KINZEY WG. 1997. Synopsis of New World Primates (16 genera): Alouatta. Pp. 174-185, en: New World Primates: Ecology, Evolution, and Behavior (WG Kinzey, ed.). Aldine de Gruyter, New York. [ Links ]
21. MATSUDA I, A TUUGA, Y AKIYAMA y S HIGASHI. 2008. Selection of river crossing location and sleeping site by Proboscis monkeys (Nasalis larvatus) in Sabah, Malaysia. American Journal of Primatology 70:1097-1101. [ Links ]
22. NEVILLE MK, KE GLANDER, F BRAZA y AB RYLANDS. 1988. The Howling Monkeys, Genus Alouatta. Pp. 349-453, en: Ecology and Behavior of Neotropical Primates (RA Mittermeier, AB Rylands, AF Coimbra- Filho y GAB da Fonseca, eds.). World Wildlife Fund, Washington. [ Links ]
23. PERES CA, JL PATTON y MF DA SILVA. 1996. Riverine barriers and gene flow in Amazonian Saddle-back tamarins. Folia Primatologica 67:113-124. [ Links ]
24. TANDY M. 2015. Photographs of the natural world. http://photographsofthenaturalworld.wordpress.com/. [ Links ]
25. WATA K. 1981. Habitat utilization by wintering Japanese macaques (Macaca fuscata fuscata) in the Shiga Heights. Primates 22:330-348. [ Links ]
26. WILLIAMSON EA, CEG TUTIN y M FERNANDEZ. 1988. Western lowland Gorillas feeding in streams and on savannas. Primate Report 19:29-34. [ Links ]
27. YEAGER CP. 1991. Possible antipredator behavior associated with river crossings by proboscis monkeys (Nasalis larvatus). American Journal of Primatology 24:61-66. [ Links ]

