










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. vol.23 no.2 Mendoza dic. 2016
NOTA
Madrigueras de Dolichotis patagonum como recurso para otros vertebrados en Península Valdés
Virginia Alonso Roldán1 y Daniel Udrizar Sauthier1, 2
1 Instituto Patagónico para el Estudio de los Ecosistemas Continentales (IPEEC), CONICET. Bvd. Brown 2915, 9120 Puerto Madryn, Chubut, Argentina. [Correspondencia: Virginia Alonso Roldán <virginia.a.roldan@gmail.com>].
2 Facultad de Ciencias Naturales, sede Puerto Madryn, Universidad Nacional de la Patagonia San Juan Bosco. Bvd. Brown 3051, 9120 Puerto Madryn, Chubut, Argentina.
Recibido 30 noviembre 2015.
Aceptado 15 junio 2016.
Editor asociado: M Busch
RESUMEN.
La mara (Dolichotis patagonum) construye madrigueras que podrían constituir un recurso para otras especies. Con el fin de estimar la abundancia de madrigueras y documentar su utilización por parte de otros vertebrados, se realizaron relevamientos en seis áreas en Península Valdés y se observaron 42 madrigueras con cámaras de video durante el día. En promedio se encontraron 0.62 madrigueras/km2 y se registraron cinco especies adicionales utilizándolas. Concluimos que la mara provee un recurso utilizado por otras especies y es un caso de estudio que debe ser profundizado en el marco de la ingeniería de ecosistemas.
ABSTRACT.
Warrens of Dolichotis patagonum as resource for other vertebrates in Península Valdés.
Maras (Dolichotis patagonum) dig warrens which could be a resource for other species. In order to estimate the abundance of warrens and to document their use by other vertebrates, surveys were conducted in six areas of Península Valdés and 42 warrens were surveyed with video cameras during day-time. We found an average of 0.62 warrens/km2 and recorded use by five additional species. We conclude that maras provide a resource used by other species and is a case study that must be further researched within the framework of ecosystem engineering theory.
Palabras clave: Interacciones interespecíficas; Mamíferos excavadores; Mara; Patagonia; Refugio.
Keywords: Burrowing mammals; Interspecific interactions; Mara; Patagonia; Shelter.
Un ingeniero de ecosistemas es un organismo que modula la disponibilidad de recursos para otras especies mediante cambios en el estado de materiales bióticos o abióticos (Jones et al., 1994). A pesar de que este concepto ha sido objetado por su posible aplicación a todos los organismos, se rescata su utilidad como marco teórico de referencia para estudiar las interacciones bióticas mediadas por cambios en el ambiente físico (Jones y Gutiérrez, 2007). En este contexto, los mamíferos excavadores son un claro ejemplo, pues modifican su entorno creando bocas de entrada y conductos en la matriz del suelo que, además de provocar cambios relacionados con los procesos de formación del suelo (Jones et al., 2006), aumentan la oferta de refugio para otras especies de vertebrados y hábitat adecuado para invertebrados (Machicote et al., 2004; McDonough y Loughry, 2008). Particularmente en ecosistemas áridos, las cuevas representan un valioso recurso como refugio para evitar los predadores y atenuar las duras condiciones ambientales (Reichman y Smith, 1990; Kinlaw, 1999). Es frecuente observar la reutilización de una cueva a través del tiempo por individuos de la misma o diferentes especies (e. g. Ceballos et al., 1999; Williams et al., 1999; Machicote et al., 2004). Con este comportamiento las especies que ocupan las cuevas de los excavadores primarios se benefician pues ahorran tiempo y energía (Kinlaw, 1999).
La mara (Dolichotis patagonum), como otros mamíferos excavadores, podría ser considerada como un organismo ingeniero de ecosistemas. Esta especie construye madrigueras con sistemas de túneles de hasta 50 m de longitud (Taber, 1987), con una o más bocas de entrada, que los cachorros mantienen limpios y despejados (Dubost y Genest, 1974; Taber y MacDonald, 1992a). Los individuos pueden utilizar la misma madriguera año tras año durante la temporada reproductiva (agosto a diciembre) para refugiar a los cachorros, e incluso varias parejas pueden utilizar conjuntamente una madriguera (Taber y MacDonald, 1992b; Baldi, 2007; Alonso Roldán, 2012). Por otra parte, algunas madrigueras utilizadas durante una temporada reproductiva no vuelven a serlo la temporada siguiente y ninguna es ocupada fuera de la temporada reproductiva (marzo a julio), ya que los adultos no la utilizan como refugio (Taber y MacDonald, 1992b; Alonso Roldán, 2012).
Aquí se analiza el rol de las madrigueras de mara como recurso para otras especies, como un primer paso en la evaluación de la función de la mara como ingeniero de ecosistemas. Nuestros objetivos fueron: estimar la abundancia de madrigueras de mara y documentar su utilización por parte de otros vertebrados.
Este estudio se llevó a cabo en el Área Natural Protegida Península Valdés, declarada Patrimonio Natural de la Humanidad por UNESCO en 1999. Fueron seleccionadas 6 áreas de 2000 ha, en las cuales se tenían registros de maras (Fig. 1). Estas áreas se consideraron de a pares, en ambientes contrastantes de la Península Valdés: arbustal (estepa arbustiva de Chondalia microphilla y Licyum spp.), pastizal (estepa herbácea de Piptochaetium napostaense, Nassella tenuis y Plantago patagónica) y mosaico de arbustal y pastizal (estepa arbustiva-herbácea de Chuquiraga avellanedae y Nassella tenuis), seleccionados en base a la caracterización florística de Bertiller et al. (1981). En cada área se realizaron búsquedas de madrigueras durante el verano 2008-2009 siguiendo 20 transectas paralelas de 5 km de largo y separadas por 200 m (para más precisiones, consultar Alonso Roldán et al., 2015). Se calculó el número medio de madrigueras por kilómetro cuadrado en cada ambiente. En agosto de 2010, las madrigueras que habían sido localizadas se inspeccionaron para verificar que siguieran en uso y no hubieran sido obstruidas por sedimento. De esta forma, fueron seleccionadas 42 madrigueras donde se registró la actividad en la madriguera y su entorno durante las horas de luz con cámaras de video (ver detalles en Alonso Roldán et al., 2015; Fig. 2). Las cámaras fueron cambiadas de ubicación cada 24 hs, volviendo a localizarse en una misma madriguera cada 15 días. Se completaron seis ciclos de observación en cada madriguera, totalizando 1913.6 horas de filmación. Las cámaras se dispusieron a aproximadamente 15 m de la boca de la madriguera y los muestreos fueron realizados entre principios de septiembre y finales de noviembre. Los videos resultantes se observaron en busca de actividad de mara y otras especies que hicieran uso de las madrigueras durante el día. Se determinó que una madriguera era utilizada si se observaba entrar o salir de la misma a algún vertebrado. Adicionalmente, se consideraron observaciones asistemáticas realizadas en madrigueras distribuidas en otros sectores de la Península Valdés.
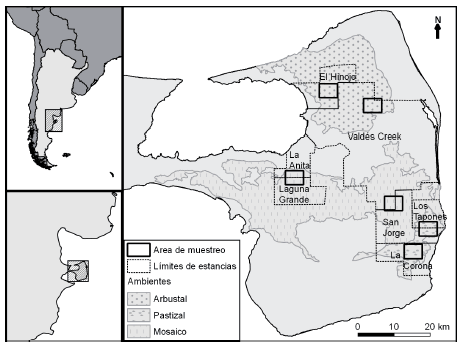
Fig. 1. Ubicación geográfica de las seis áreas que fueron consideradas en este trabajo. Se muestran los límites de las estancias en las cuales se trabajó y también de las principales unidades florísticas de la Península Valdés (sensu Bertiller et al., 1981).

Fig. 2. a) Equipo de filmación dispuesto a 15 m de una madriguera en ambiente de mosaico de arbustal y pastizal; b) ejemplo de una imagen capturada por la cámara en una madriguera utilizada por maras; c) ejemplo de una imagen capturada por la cámara en una madriguera de maras utilizada por peludos; la flecha indica al peludo ingresando en la madriguera.
En promedio, se detectaron 0.62 madrigueras/km2. Del total de madrigueras filmadas (n = 42), el 19% fue utilizado por maras (siendo estas madrigueras reutilizadas, pues todas habían sido detectadas después de la temporada reproductiva anterior). De las 42 madrigueras, en el 26% se registró actividad de uso por parte de otras especies de vertebrados. De acuerdo a nuestros registros, las madrigueras fueron uti lizadas por lechucita de las vizcacheras (Athene cunicularia), liebre europea (Lepus europaeus), peludo (Chaetophractus villosus), zorrino (Conepatus chinga), martineta (Eudromia elegans), zorro gris (Lycalopex gymnocercus) y yarará ñata (Bothrops ammodytoides) (Fig. 3). También se observaron lagartijas del género Liolaemus en las madrigueras durante los registros asistemáticos.
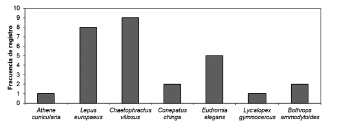
Fig. 3. Frecuencia de registro de especies de vertebrados en las madrigueras de mara.
En siete de los casos en que se observaron peludos ingresando a las madrigueras las mismas no estaban siendo usadas por maras. Sin embargo, en un caso se registraron adultos de mara y en otro la madriguera estaba ocupada por dos crías de mara. En ambos casos, después del registro de los peludos ingresando en la madriguera no volvió a registrarse la presencia de maras. De igual manera, se observó el cese de la actividad de maras en la madriguera donde se registró el ingreso de un individuo de zorro gris. Por el contrario, las crías de varias parejas de maras alojadas en una madriguera no la abandonaron luego de registrarse el ingreso y permanencia por unos 30 minutos de un zorrino. Tampoco se observó el abandono de las madrigueras en que se había registrado el ingreso de liebres europeas o martinetas.
Con su actividad excavadora, la mara genera posibles refugios que son utilizados por otras especies, por lo que puede ser considerada como un ingeniero de ecosistemas. Si bien la densidad de madrigueras de mara es baja comparada con las densidades reportadas para otros organismos considerados ingenieros de ecosistemas, como el conejo europeo (Oryctolagus cuniculus; 250 bocas/ha en la península Ibérica; Delibes Mateos y Gálvez Bravo, 2009) o el perro de la pradera (Cynomys ludovicianus; 52.7 madrigueras/ha en México; Ceballos et al., 1999), la oferta de este recurso no sería limitante para su uso, ya que solo el 26% de las madrigueras fueron utilizadas por otras especies de vertebrados. Sin embargo, es posible que la proporción de madrigueras utilizadas pueda estar subestimada, ya que la metodología utilizada no registra especies de actividad nocturna (e. g. gatos, hurones, ratones de campo, etc.). Por otra parte, la disponibilidad de este recurso se mantendría todo el año, pues incluso durante la temporada reproductiva de maras solo el 19% de las madrigueras fue reutilizado por esta especie. Esta baja proporción de utilización por otras especies se ha reportado también en el caso de la vizcacha (Lagostomus maximus) y la lechucita de las vizcacheras en la provincia de La Pampa (Machicote et al., 2004). La baja tasa de uso no está, necesariamente, relacionada con la importancia del recurso o la relación entre especies. Por ejemplo, la lechucita de las vizcacheras usa en una tasa similar las cuevas de vizcacha (25.75%) pero su éxito reproductivo depende de ellas ya que los nidos fuera de las colonias de vizcacha fallan (Machicote et al., 2004); algo parecido ocurre con la disponibilidad de madrigueras de perros de las praderas en Estados Unidos (Desmond et al., 2000). La relación entre la mara y la lechucita de las vizcacheras podría ser equivalente a los casos anteriores ya que hemos registrado el uso de madrigueras por parte de lechucitas de las vizcacheras y las características del hábitat donde se encuentran las madrigueras de mara estarían asociadas a la detección de predadores y son similares a las que se vinculan con el éxito reproductivo de las lechucitas de las vizcacheras en La Pampa (Taber y MacDonald, 1992b; Machicote et al., 2004; Baldi, 2007; Alonso Roldán, 2012).
Los casos en que la presencia de peludos provocó el abandono de madrigueras por parte de las maras plantean una interacción en que estas últimas podrían verse perjudicadas. Esta situación podría generar una potencial disminución en el éxito reproductivo si las parejas son desplazadas a sitios con características menos favorables (Baldi, 2007); o si las crías quedan temporalmente sin protección (Dubost y Genest, 1974) o si las crías estuvieran siendo atacadas por peludos y esta fuera la razón para el desplazamiento. En cuanto a esta última hipótesis, es interesante notar que la reacción de las maras fue similar en los casos en que se registró la presencia de zorros, señalados como depredadores de crías de mara (Taber, 1987).
Las interacciones reseñadas en esta nota señalan a la mara como un caso de estudio que debe ser profundizado en el marco de la ingeniería de ecosistemas. De esta investigación se desprende que son necesarios estudios con metodologías específicas enfocados en cada una de las posibles interacciones entre las especies mencionadas para determinar si la utilización de madrigueras es oportunista o si existe dependencia del recurso por parte de las especies que las utilizan. En este contexto sería valioso que futuros estudios midan variables como impactos relacionados con la riqueza de especies, relaciones entre estructuras físicas (madrigueras) y variables abióticas físicas y químicas, para conocer su efecto, función en el sistema y mejorar el entendimiento de los procesos que dan forma a los ecosistemas patagónicos.
Agradecimientos.
Agradecemos a los propietarios de los establecimientos El Hinojo, La Anita, Laguna Grande, Valdés Creek, San Jorge y La Corona, que permitieron trabajar en sus propiedades, y especialmente a Luis Porcel por su ayuda con relación al alojamiento y a “cuestiones de supervivencia” de los investigadores. A P. Torres, P. Contreras, S. Días, R. L. Vaughn, N. Hughes, P. Faiferman, C. Perondi, M. Agrelo, M. Zamero, N. Velázques, C. Pascheta, A. Formoso, L. Feugeas, D. E. Galván, P. Hackerman, L. Beltramino y R. D’Agostino por su asistencia en los trabajos de campo. A Cynthia Fernández por su ayuda en el análisis de los videos, a D. E. Galván, N. Velázquez Barloa, a la editora asociada M. Busch y a dos revisores anónimos por los comentarios realizados sobre versiones previas de este trabajo que ayudaron a mejorarlo. El Centro Nacional Patagónico y las fundaciones Patagonia Natural y Vida Silvestre Argentina (Reserva de Vida Silvestre San Pablo de Valdés) proveyeron soporte logístico. Las tareas de campo fueron autorizadas por la Dirección de Fauna y Flora Silvestre y la Secretaría de Turismo y Áreas Protegidas de la provincia del Chubut. Los fondos para este trabajo fueron provistos por People Trust for Endangered Species, Idea Wild, ALUAR y el Gobierno de la Provincia del Chubut.
LITERATURA CITADA
1. ALONSO ROLDÁN V. 2012. Patrones de distribución de la mara (Dolichotis patagonum) a distintas escalas. Tesis de Doctorado, Departamento de Biología, Bioquímica y Farmacia, Universidad Nacional del Sur, Bahía Blanca. [ Links ]
2. ALONSO ROLDÁN V, L BOSSIO y DE GALVÁN. 2015. Sources of variation in a two-step monitoring protocol for species clustered inconspicuous points: Dolichotis patagonum as a case study. PLoS ONE 10.1371/journal. pone.0128133 [ Links ]
3. BALDI R. 2007. Breeding success of the endemic mara Dolichotis patagonum in relation to habitat selection: Conservation implications. Journal of Arid Environments 68:9-19. [ Links ]
4. BERTILLER MB, AM BEESKOW y O IRRISARI. 1981. Caracteres fisonómicos y florísticos de la vegetación del Chubut. 2. La Península de Valdés y el Istmo Ameghino. Contribución 41. CONICET. Centro Nacional Patagónico, Puerto Madryn. [ Links ]
5. CEBALLOS G, J PACHECO y R LIST. 1999. Influence of prairie dogs Cynomys ludovicianus on habitat heterogeneity and mammalian diversity in Mexico. Journal of Arid Environments 41:161-172. [ Links ]
6. DELIBES MATEOS M y L GÁLVEZ BRAVO. 2009. El papel del conejo como especie clave multifuncional en el ecosistema mediterráneo de la Península Ibérica. Ecosistemas 18:14-25. [ Links ]
7. DESMOND MJ, JA SAVIDGE y KM ESKRIDGE. 2000. Correlations between burrowing owl and black-tailed prairie dog declines: A 7-year analysis. Papers in Natural Resources Paper 162:http://digitalcommons.unl.edu/natrespapers/162. [ Links ]
8. DUBOST G y H GENEST. 1974. Le comportement social d’une colonie de maras Dolichotis patagonum Z. dans le parc de Branfere. Zeitschrift für Tierpsychologie 35:225-302.
9. JONES CG y JL GUTIÉRREZ. 2007. On the purpose, meaning, and usage of the ecosystem engineering concept. Pp. 3-24, en: Ecosystem engineers: Plants to protists (K Ciddington, JE Byers, WG Wilson y A Hastings, eds.). Acasemic Press. [ Links ]
10. JONES CG, JL GUTIÉRREZ, PM GROFFMAN y M SHACHAK. 2006. Linking ecosystem engineers to soil processes: A framework using the Jenny State Factor Equation. European Journal of Soil Biology 42:39-53. [ Links ]
11. JONES CG, JH LAWTON y M SHACHAK. 1994. Organisms as ecosystem engineers. Oikos 69:373-386. [ Links ]
12. KINLAW A. 1999. A review of burrowing by semi-fossorial vertebrates in arid environments. Journal of Arid Environments 41:127-145. [ Links ]
13. MACHICOTE M, LC BRANCH y D VILLARREAL. 2004. Burrowing owls and burrowing mammals: Are ecosystem engineers interchangeable as facilitators? Oikos 106:527-535. [ Links ]
14. MCDONOUGH CM y WJ LOUGHRY. 2008. Behavioral ecology of armadillos. Pp. 281-293, en: The biology of the Xenartha (SF Vizcaíno y WJ Loughry, eds.). University Press of Florida. [ Links ]
15. REICHMAN OJ y SC SMITH. 1990. Burrows and burrowing behaviour by mammals. Pp. 197-244, en: Current Mammalogy (HH Genoways, ed.). Plenum Press. [ Links ]
16. TABER AB. 1987. The behavioural ecology of the mara Dolichotis patagonum. Ph.D. dissertation, University of Oxford, United Kingdom. [ Links ]
17. TABER AB y DW MACDONALD. 1992a. Communal breeding in the mara, Dolichotis patagonum (Rodentia: Caviomorpha). Journal of Zoology of London 227:439-452. [ Links ]
18. TABER AB y DW MACDONALD. 1992b. Spatial organization and monogamy in the mara Dolichotis patagonum. Journal of Zoology of London 227:417-438. [ Links ]
19. WILLIAMS JB, BI TIELEMAN y M SHOBRAK. 1999. Lizard burrows provide thermal refugia for larks in the Arabian desert. The Condor 101:714-717. [ Links ]

