










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. vol.24 no.2 Mendoza dic. 2017
ARTÍCULO
Differences in the mammalian habitat use in a mosaic of vegetation types of an Atlantic rain-forest reserve, Brazil
Atilla C. Ferreguetti1, Walfrido M. Tomas2, and Helena G. Bergallo1
1 Departament of Ecology, Rio de Janeiro State University, Rio de Janeiro, Brazil [Correspondence: Atilla C. Ferreguetti <atilla.ferreguetti@gmail.com>].
2 Wildlife Laboratory, Embrapa Pantanal, Mato Grosso do Sul, Brazil.
Recibido 28 mayo 2017.
Aceptado 5 july 2017.
Editor asociado: T Lacher
ABSTRACT.
This study aimed to evaluate how habitat use by medium and large sized mammals was related to different vegetation types in an Atlantic rain-forest reserve in south-eastern Brazil. Our hypotheses were that each vegetation type found in the Vale Natural Reserve (VNR) will present a different mammal species composition, and that mammal species richness and relative abundance will be higher in areas nearest to water. We installed 39 camera traps in the VNR, and used two covariates to characterize the different vegetation types (density of trees and lianas). We also used one spatial covariate: the distance to the nearest water resource. We recorded 26 medium- and large-sized mammal species. The mammalian assemblage was structured by vegetation type and the proximity of water. The results presented here show that even in a single reserve such as VNR there is a difference in the assemblage composition of medium- and large-sized mammals. Our results also indicate the relevance of water availability when designing protected areas to ensure habitat quality for mammal species. The VNR is one of the major remaining portions of the coastal plain forest of the Atlantic rain forest, and also one of the largest contiguous blocks of this biome.
RESUMO.
Diferenças no uso do habitat por mamíferos em um mosaiaco de fitofisionomias em uma reserva de Mata Atlântica, Brasil.
O objetivo desse estudo foi verificar como o uso do habitat por espécies de mamíferos de médio e grande porte estava relacionada com as diferentes fitosionomias em uma área protegida de Mata Atlântica, Sudeste do Brasil. Nossas hipóteses foram: cada fitofisionomia encontrada na Reserva Natural Vale (RNV) apresentaria diferentes composições de espécies de mamíferos, e que a riqueza e abundancia relativa das espécies seria maior em áreas próximas dos recursos hídricos. Nós instalamos 39 armadilhas fotográficas na RNV, e utilizamos duas covariadas para caracterizar as diferentes fitofisionomias (i.e. densidade de árvores e de lianas). Nós também utilizamos uma covariada espacial: a distância do recurso hídrico mais próximo. Nós registramos 26 espécies de mamíferos de médio e grande porte. A comunidade de mamíferos foi estruturada pelos diferentes tipos de fitofisionomias e pela proximidade da água. Os resultados apresentados aqui mostra que mesmo em uma única reserva é, como a RNV, existe uma diferença na composição de mamíferos de médio e grande porte. Além disso, nossos resultados também destacam a relevância da água na determinação de uma área protegida para garantir a qualidade do habitat para as espécies de mamíferos. A RNV faz parte de um dos maiores blocos de Mata de Tabuleiro da Mata Atlântica, e também representam um dos maiores remanescentes do bioma.
Key words: Abiotic effects; Camera-traps; Coastal plain forest; Community ecology; Mammalian species.
Palavras-chave: Armadilhas fotográficas; Efeitos abióticos; Ecologia de comunidade; Mammalia; Mata de tabuleiro.
INTRODUCTION
The hypothesis that habitat heterogeneity determines an increase in species diversity, initially developed by MacArthur & MacArthur (1961), considers that more heterogeneous environments provide greater resource availability, which would result in more niches and, consequently, would support higher species diversity than simpler environments (Bazzaz 1975). This positive relationship between increased habitat heterogeneity and increasing species richness has already been registered for several taxa (Moreno-Rueda & Pizarro 2009; Stein et al. 2014). However, depending on the taxonomic group and the spatial scale, richness can also have a negative relation with increasing habitat heterogeneity (Tews et al. 2004; González- Megias et al. 2007).
The Vale Natural Reserve (VNR) is a private protected area covered densely by the Atlantic rain forest (IBGE 1993), composed of a mosaic of habitats with four main vegetation types with different levels of habitat heterogeneity (adapted from Jesus 1987; Peixoto & Gentry 1990): i) coastal plain forest (tabuleiro forest), ii) ecotone forest, iii) sandy soil forest (mussununga forest), and iv) natural grassland. The coastal plain forest is the most representative vegetation type in the VNR (about 68% of the reserve). It differs from other vegetation types by a higher density of trees with a diameter at breast height (dbh) larger than 30 cm (average of 13 trees per ha), with heights ranging from 5 to 31 m (average = 24 m). It presents a lower density of lianas (average of 21.8 lianas per ha) with lower understorey cover (34%). Sandy soil forest, which covers approximately 8% of the VNR, is a type of woody vegetation on sandy soils, physiognomically similar to the early or intermediate stage of regeneration of the coastal plain forest. This vegetation presents a lower density of trees with dbh > 30 cm (average of 8.7 trees per ha) and tree height between 6 to 15 m. There is a high density of lianas (average of 82.2 lianas per ha), and bromeliads. The ecotone forests, which cover about 10% of the reserve, are represented by a transition between coastal plain and sandy soil forests. These transition areas have an average density of trees with dbh > 30 cm around 6.5 per ha, and a predominance of palm tree species. The natural grasslands, which cover about 6% of the reserve, occur in sandy soils and differ from other vegetation types found in VNR due to the predominance of grasses and shrubs. This type of vegetation occurs forming enclaves within the forest habitat. Additionally, approximately 8% of the reserve is covered by wetlands (swamps) and streams.
In addition to these differences between the types of vegetation, water can also be a limiting factor for the richness and abundance of species present in an assemblage. Temporal environmental variation also influences the structure and species composition (i.e. which species may occur) of a community (O’Connell 1989). The seasonality of water resources may be an important element influencing the structure of animal communities in tropical forests (Hawkins et al. 2003), especially under scenarios of climate change and the observed lower precipitation rates in the last decades (Marengo 2006; Dos Santos et al. 2010; Nobre et al. 2016). Usually, areas located near water resources have higher food availability for aquatic and terrestrial species (Naiman et al. 1993; Kondolf et al. 1996). Such areas can hold a higher diversity of mammals (Naiman et al. 2005), as many mammal species may be directly associated with/or even dependent on water for food resource, and even shelter (Virgós 2001; Naiman et al. 2005).
Even with all the existing information about Neotropical species of medium and large mammals, there are still many gaps in our knowledge (Voss & Emmons 1996; Costa et al. 2005), especially when compared to other groups of vertebrates (e.g. reptiles, amphibians, birds and small mammals). Different mammalian assemblages in the world are very heterogeneous (Ceballos et al. 2005). The Neotropical region is the most diverse in the number of species (Cole et al. 1994) and Brazil is the country with the highest diversity of mammals (Costa et al. 2005; Paglia et al. 2012). Usually, the difference between the composition of medium-large mammal species has been demonstrated on a regional scale because large-bodied species require large ranges (Fernández & Vrba 2005; Haugaasen & Peres 2005; Parry et al. 2007). However, on a smaller scale (local), it is not expected to find a well-defined difference in the composition of medium- and large-sized mammals between types of vegetation due to the large body size of these species and the presence of large home ranges.
This study aimed to evaluate how the habitat use by medium- and large-sized mammals related to different vegetation types on a local scale, and how the distance from water resources affect the assemblage in the VNR. We hypothesize that (1) each vegetation type found in the VNR will present a difference in species composition; and (2) mammal species richness and relative abundance will be higher in areas nearest to water.
MATERIAL AND METHODS
Study area
This study was conducted in Vale Natural Reserve (VNR), a protected area of 23 000 hectares belonging to the Vale Company. The reserve is located in the neighboring municipalities of Linhares and Jaguaré (19° 06’-19° 18’ S, 39° 45’-40° 19’ W), in north-eastern Espírito Santo, Brazil (Fig. 1).
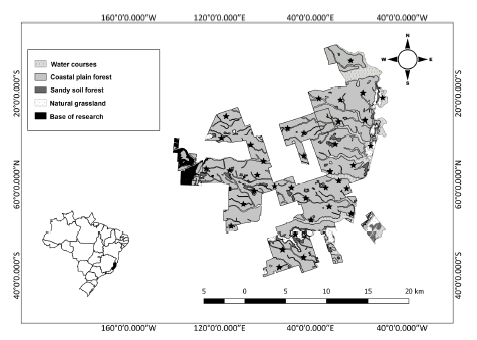
Fig. 1. Distribution of camera traps (represented by stars) at the Vale Natural Reserve, Linhares, Espírito Santo state, Brazil, sampled from May 2013 to June 2014.
Methods
Camera-trapping
We selected 39 sampling sites using a systematic random design stratified by vegetation type to ensure that all four of the principal vegetation types found in the VNR were represented. We placed a random grid over a digital map of the reserve, and identified the sampling points by selecting grid cells divided by each region of the reserve (i.e. north, west, and south, Fig. 1). In each selected grid cell, we randomly located a camera trap. This approach resulted in a proportional distribution of points within the VNR, while maintaining independence among points, which were separated from one another by a distance of more than 1 km (Ancrenaz et al. 2012). At each site, we installed one passive infrared sensor (i.e. detection caused by movement from individuals) Bushnell® camera trap in picture function, approximately 40-50 cm above the ground, for continuous surveying throughout the study (May 2013-June 2014). All stations were examined every 20-25 days to change batteries, when necessary. We did not use bait to attract mammals.
Measurement of covariates
We used three covariates to characterize habitat heterogeneity of each sampled location (i.e. camera-trap points). Two were density of trees with a dbh > 30 cm, and density of lianas. We also used one spatial covariate, the distance to the nearest water resource (m).
At each sample site, we established four plots (30 × 50 m) arranged by the cardinal compass points (north, south, east and west). In each plot, we measured the dbh of each tree and counted the number of trees with dbh > 30 cm. The density of lianas was measured along the central (longitudinal) line of each plot, considering 5 m on each side of the line (transect width of 10 m). The density of lianas was based on the number of stems rooted within each 10 × 50-m transect. Distance to the nearest water resource was measured for each sampled sites using the ArcGIS software (ESRI*ArcMap 10.1).
Data analysis
For each camera-trap, we calculated the richness of medium- and large-sized mammal species. We calculated the photographic rate of each species by dividing the number of captures (photo or sequence of photos) of each species at each site by the total days of sampling. Photocaptures of a given species at a site, on the same day, were considered as a unique capture, in order to avoid pseudoreplication. We tested for difference in species richness between the different portions in VNR (i.e. north, south, and west) using analysis of variance (ANOVA).
To test for variation in beta-diversity between vegetation types we used the ‘betadisper’ function in the R software Vegan package version 2.0-4 for community analysis (Oksanen et al. 2013). In this test, b-diversity is defined as the average distance-to-centroid, measured as the average distance using the Bray-Curtis dissimilarity index (or compositional dissimilarity) from a camera-trap to the centroid of the group of all camera-traps within a region.
A Canonical Correspondence Analysis (CCA) was used in order to investigate the relationship between species composition and photographic rates to environmental covariates. With this analysis we aimed to assess the possible influence of covariates that could represent a key-habitat feature for the mammalian species. Normality and homogeneity of variance of the data were verified prior to the analysis, and all data accorded with these assumptions. Canonical correspondence analysis, a direct gradient analysis, was used because of its ability to examine several environmental gradients simultane ously (Palmer 1993). It also gives reliable results even with interrelationships among habitat characteristics (i.e. multicollinearity) and skewed distributions of species (e.g. the distribution of specialist species among the sites sampled, Palmer 1993). The analyses were performed in R version 2.15.0 using a CCA function in the Vegan package version 2.0-4 for community analysis (Oksanen et al. 2013).
We performed simple linear regressions using the first two axes of the CCA to test the effect of habitat covariates and water resource in the mammalian community. Forward stepwise multiple linear regression was conducted to analyze the relationships between habitat covariates and the partial species richness. Analyses were performed in R version 2.15.0.
RESULTS
A total of 23 245 photos of 26 species of medium- and large-sized mammals were obtained from an effort of 7020 trapping days. We found a higher mammal species richness in the northern portion (22), than in the south and west (17) (ANOVA, df = 36; F = 11.6, P < 0.001). With the beta-diversity analysis we found that vegetation types of forest were on average 72% more similar to each other than with respect to non-forest environments that showed 33% similarity to the forest areas of the VNR. The photographic rates by species and species list are shown in Appendix 1.
As a summary, the two-dimensional CCA plot allowed the simultaneous representation of the similarity between sites, and the relationship between mammals and measured covariates (Canonical R = 0.81; df = 33; F = 9.54; P < 0.001; Fig. 2). The first Axis of CCA indicated that the mammalian community was structured according to density of trees with dbh > 30 cm (R2 = 0.81; df = 38; P < 0.001; Fig. 3). Species such as Dasyprocta leporina, Guerlinguetus brasiliensis, Eira barbara, and Mazama americana had a positive relationship with the density of trees. The species Euphractus sexcinctus, Puma yagouaroundi, Sylvilagus brasiliensis, Mazama gouazoubira and Cerdocyon thous had a negative relationship with the density of trees, as they occurred more frequently in the native grasslands (Fig. 2).

Fig. 2. Bi-plot and scores from Canonical Correspondence Analysis (CCA) plotted for species, sites, and habitat covariates of the Vale Natural Reserve, Espírito Santo, Brazil (No. of sites = 39). The first two canonical axes with the percentage of variance explained and the eigenvalues (between parentheses). Dots represents sample sites, arrows represent environmental covariates, and abbreviated name of each species with the first two letters of the genus and specific epithet. tate = Tapirus terrestris, nana = Nasua nasua, sybr = Sylvilagus brasiliensis, dano = Dasypus novemcinctus, euse = Euphractus sexcinctus, tamte = Tamandua tetradactyla, lepa = Leopardus pardalis, puya = Puma yagouarundi, tape = Tayassu pecari, peta = Pecari tajacu, guin = Guerlinguetus ingrami, cupa = Cuniculus paca, maam = Mazama americana, mago = Mazama gouazoubira, eiba = Eira barbara, dale = Dasyprocta leporina. ceth = Cerdoncyon thous, and diau = Didelphis aurita. Gray circle = natural grasslands, black circle = coastal forest, black square = sandy forest, and gray square = ecotone. Habitat covariates: dist_water = distance to water resource, dens_tree = density of trees with dbh > 30 cm, and dens_liana = density of lianas.

Fig. 3. Relationship between the canonical axes 1 (CCA 1) and the abundance and composition of species for each camera-trap located in the Vale Natural Reserve, Linhares, Espírito Santo, Brazil, sampled from May 2013 to June 2014. guin = Guerlinguetus brasiliensis, maam = Mazama americana, eiba = Eira barbara, lepa = Leopardus pardalis, dano = Dasypus novemcinctus, dale = Dasyprocta leporina, nana = Nasua nasua, cupa = Cuniculus paca, tate = Tapirus terrestris, tape = Tayassu pecari, peta = Pecari tajacu, diau = Didelphis aurita, mago = Mazama gouazoubira, ceth = Cerdoncyon thous, euse = Euphractus sexcinctus, tamte = Tamandua tetradactyla, sybr = Sylvilagus brasiliensis, and puya = Puma yagouarundi.
The second axis of CCA indicated that the distance to the nearest water resource was also a covariate responsible for structuring mammalian community (R2 = 0.94; df = 38; P < 0.001; Fig. 4). Tapirus terrestris, Tayassu pecari and Cuniculus paca were the species more associated with proximity to a water resource.

Fig. 4. Relationship between the canonical axes 2 (CCA 2) and the abundance and composition of species for each camera-trap located in the Vale Natural Reserve, Linhares, Espírito Santo, Brazil, sampled from. May 2013 to June 2014. diau = Didelphis aurita, mago = Mazama gouazoubira, maam = Mazama americana, guin = Guerlinguetus brasiliensis, puya = Puma yagouarundi, dale = Dasyprocta leporina, euse = Euphractus sexcinctus, ceth = Cerdoncyon thous, sybr = Sylvilagus brasiliensis, tate = Tapirus terrestris, tamte = Tamandua tetradactyla, nana = Nasua nasua, lepa = Leopardus pardalis, tape = Tayassu pecari, dano = Dasypus novemcinctus, peta = Pecari tajacu, cupa = Cuniculus paca, and eiba = Eira Barbara.
We found a positive relationship between the partial mammal species richness and the partial of density of trees with dbh > 30 cm (R2 = 0.72; df = 38; P < 0.001; Fig. 3). We also found a negative relationship between the partial mammal’s richness and the partial of distance to the nearest water resource (R2 = 0.81; df = 38; P < 0.001; Fig. 4).
DISCUSSION
We found a relatively high richness of medium- to large-sized mammal species (26). This may be related to the method used in our study (camera-trap). By comparing the data from this study with those of other studies that used camera traps, we may conclude that the effort was enough for making accurate claims and predictions about the mammalian community. Srbek-Araujo & Chiarello (2005) recorded 21 species for the mountain region of Espírito Santo state, and Srbek-Araujo & Chiarello (2013) recorded 28 species for the Linhares-Sooretama protected area block in Brazil, which includes our study area, VNR, based on larger effort (4 y). Ahumada et al. (2011) by evaluating mammal species from seven rain-forest sites in Africa, Asia and Latin America found a richness average of 20.6, range, 13-28 species per site.
We observed differences in the relative abundance, species richness and the structure of large- and medium-sized mammal assemblages at different vegetation types of the VNR. This could be explained by differences in habitat heterogeneity between the vegetation types, because habitat heterogeneity plays an important role in structuring mammalian communities in different tropical environments (August 1983; Tews et al. 2004; Haugassen & Peres 2005; Stein et al. 2014), and it is not surprising that several studies have demonstrated that species richness is proportional to habitat heterogeneity in the Atlantic rain forest as well (Grelle 2003; Vieira & Monteiro-Filho 2003; Prevedello et al. 2008). In fact, the species more closely associated with areas with a higher degree of habitat heterogeneity (i.e. forest areas with greater resource availability) are predominantly present in forested areas, which is the case of Dasyprocta leporina and Mazama americana (Silvius & Fragoso 2003; Ferreguetti et al. 2015). Areas with lower habitat heterogeneity (i.e. open areas) were represented by species of mammals that usually occur in open vegetation types, which is the case of Sylvilagus brasiliensis, Cerdocyon thous and Euphractus sexcinctus (Eisenberg & Redford 1999; Juarez & Marinho-Filho 2002; Ferreguetti et al. 2016).
Water availability is one relevant factor structuring communities of medium- to large-sized mammals in the VNR, and this may be a result of the unequal distribution of water resources in the area. The hydrography of the reserve consists of a network of two main drainage systems (Barra Seca and Pau Atravessado rivers), located in the northern and in the south regions, respectively (Jesus & Rolim 2005). The tributary streams of these two major rivers are mostly ephemeral, with a well-defined dry season (Kierulff et al. 2014), with the difference that in the northern area most streams run all year long (Jesus & Rolim 2005). The north is also where we found a higher richness of mammal species. Naiman et al. (2005) showed that sites near water resources areas can present a high diversity of mammals. This study confirms the relationship found between richness and the distance from water resource, where areas nearest to the water resource presented a higher richness of mammal species. The current dry period could presumably increase the competition for water, which in the VNR could be considered one of the biggest limiting features. In the last few years, climate change has been causing long periods of drought throughout the Atlantic rain forest of Brazil (CPTEC 2015; Nobre et al. 2016). In this scenario of lack of rainfall, many streams are disappearing (Marengo 2006; Nobre et al. 2016). The long period of drought, can make the environment more susceptible to fires (e.g. fire events in 2015 and 2016, pers. obs.), which can represent a major impact for the community of mammal species, and the whole ecosystem.
The results presented here are one of the first to show that even in a single reserve such as VNR there is a difference in the assemblage composition of medium- and large-sized mammal species. Our results also indicate the relevance of long-term water availability when designing protected areas to ensure habitat quality for mammal species. The VNR is one of the major remaining portions of the coastal plain forest of the Atlantic rain forest, and also one of the largest contiguous blocks of this biome. These results not only provide subsidies for the increase of knowledge of mammalian ecology, but also for future strategies for the conservation of these species. Therefore, more attention should be given to the monitoring of these communities in the long term, since this area could still harbor some viable populations of large mammals.
ACKNOWLEDGEMENTS
We thank Vale Natural Reserve and Vale Company for the support in the research. AFC thanks Coordenacão de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the scholarship received and Brandon LaBumbard for the English review. HGB thanks FAPERJ (E-26/103.016/2011, E26 /201.267/ 2014), Prociência/UERJ and CNPq (457458/2012-7, 307715/2009-4) for the research and productivity grants. This study is part of the Research Program in Biodiversity (PPBio MA). Data are maintained with support from PPBio MA (CNPq - 457458/2012-7) and FAPERJ (E-26/111.394/2012).
LITERATURE CITED
1. Ahumada, J. A. et al. 2011. Community structure and diversity of tropical forest mammals: data from a global camera trap network. Philosophical Transactions of the Royal Society of London Biological Sciences 366:2703-2711. [ Links ]
2. Ancrenaz, M., A. Hearn, J. Ross, R. Sollmann, & A. Wilting. 2012. Handbook for wildlife monitoring using camera-traps. Small Carnivore Specialist Group, Sabah, Malaysia. 83 pp. [ Links ]
3. August, P. V. 1983.The role of habitat complexity and heterogeneity in structuring tropical mammal communities. Ecology 64:1495-1507. [ Links ]
4. Bazzaz, F. A. 1975. Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology 56:485-488. [ Links ]
5. Ceballos, G., P. R. Ehrlich, J. Soberón, I. Salazar, & J. P. Fay. 2005. Global mammal conservation: what must we manage? Science 309:603-607. [ Links ]
6. Cole, F. R., D. M. Reeder, & D. E. Wilson. 1994. A synopsis of distribution patterns and the conservation of mammal species. Journal of Mammalogy 75:266-276. [ Links ]
7. Costa, L. P., Y. L. R. Leite, S. L. M. Mendes, & A. D. Ditchfield. 2005. Mammal conservation in Brazil. Conservation Biology 19:672-679. [ Links ]
8. CPTEC. 2015. Centro de Previsão de Tempo e Estudos Climáticos. Available from http://www.cptec.inpe.br/ (accessed November 1, 2015). [ Links ]
9. Dos Santos, A. M., G. J. Domiciano, & M. S. M. Bezerra. 2010. Water resources and climatic changes: dissertations, impacts and conflicts. Revisa Geográfica Venezolana 51:59-68. [ Links ]
10. Eisenberg, J. F., & K. H. Redford. 1999. Mammals of the Neotropics, v. 3. Chicago University Press, Chicago, 624 pp. [ Links ]
11. Fernández, M. H., & E. S. Vrba. 2005. Body size, biomic specialization and range size of African large mammals. Journal of Biogeography 32: 1243-1256. [ Links ]
12. Ferreguetti, Á. C., W. M. Tomás, & H. G. Bergallo. 2015. Density, occupancy, and activity pattern of two sympatric deer (Mazama) in the Atlantic Forest, Brazil. Journal of Mammalogy 96:1245-1254. [ Links ]
13. Ferreguetti, Á. C., W. M. Tomás, & H. G. Bergallo. 2016. Density and niche segregation of two armadillo species (Xenarthra: Dasypodidae) in the Vale Natural Reserve, Brazil. Mammalian Biology 81:138-145. [ Links ]
14. González-Megias, A., J. G. Gómes, & F. Sánchez-Piñero. 2007. Diversity-habitat heterogeneity relationship at different spatial and temporal scales. Ecography 30:31-41. [ Links ]
15. Grelle, C. E. V. 2003. Forest structure and vertical stratification of small mammals in a secondary Atlantic Forest, Southeastern Brazil. Studies on Neotropical Fauna and Environment 38:81-85. [ Links ]
16. Haugassen, T., & C.A. Peres. 2005. Mammal assemblage structure in Amazonian flooded and unflooded forests. Journal of Tropical Ecology 21:133-145. [ Links ]
17. Hawkins, B. A. et al. 2003. Energy, water, and broad-scale geographic patterns of species richness. Ecology 84:3105-3117. [ Links ]
18. IBGE. 1993. Mapa de Vegetação do Brasil. Rio de Janeiro: Fundação Instituto Brasileiro de Geografia e Estatística. [ Links ]
19. Jesus, R. M. 1987. Mata Atlântica de Linhares: Aspectos Florestais. Desenvolvimento Econômico e Impacto Ambiental em Áreas de Trópico Úmido Brasileiro: a experiência da CVRD Secretaria Estadual de Meio Ambiente, Idaho Water Resource Board & Companhia Vale do Rio Doce, eds.). A Secretaria, Rio de Janeiro, Brazil. [ Links ]
20. Jesus, R. M., & S. G. Rolim. 2005. Fitossociologia da Mata Atlântica de Tabuleiro. Boletim Técnico da Sociedade de Investigações Florestais 19:1-149. [ Links ]
21. Juarez, K. M., & J. Marinho-Filho. 2002. Diet, habitat use, and home ranges of sympatric canids in central Brazil. Journal of Mammalogy 83:925-933. [ Links ]
22. Kierulff, M. C. M., L. H. S. Avelar, M. E. S. Ferreira, K. F. Povoa, & R. S. Bernils. 2014. Reserva Natural Vale: história e aspectos físicos. Ciência & Ambiente 49:7-40. [ Links ]
23. Kondolf, G., R. Kattelmann, M. Emburry, & D. Erman. 1996. Status of riparian habitat. Sierra Nevada Ecosystem Project: Final report to Congress, vol. II, Assessments and scientific basis for management options. Davis: University of California, Centers for Water and Wildland Resources, California. [ Links ]
24. Macarthur, R. H., & J. W. Macarthur. 1961. On bird species diversity. Ecology 42:594-598. [ Links ]
25. Marengo, J. A. 2006. Mudanças climáticas globais e seus efeitos sobre a biodiversidade: caracterização do clima atual e definição das alterações climáticas para o território brasileiro ao longo do século XXI. MMA, Brasília, Brasil. [ Links ]
26. Moreno-Rueda, G., & M. Pizarro. 2009. Relative influence of habitat heterogeneity, climate, human disturbance, and spatial structure on vertebrate species richness in Spain. Ecological research 24(2):335-344. [ Links ]
27. Naiman, R., H. Décamps, & M. Pollock. 1993. The role of riparian corridors in maintaining regional biodiversity. Ecological Applications 3:209-212. [ Links ]
28. Naiman, R., H. Décamps, & M. Mcclain. 2005. Riparia: ecology, conservation, and management of streamside communities. Elsevier, Burlington, 657 pp. [ Links ]
29. Nobre, C. A., J. A. Marengo, M. E. Seluchi, A. L. Cuartas, & L. M. Alves. 2016. Some characteristics and impacts of the drought and water crisis in Southeastern Brazil during 2014 and 2015. Journal of Water Resource and Protection 8:252-262. [ Links ]
30. O’Connell, M. A. 1989. Population dynamics of Neotropical small mammals in seasonal habitats. Journal of Mammalogy 70:532-548.
31. Oksanen, J. et al. 2013. Vegan: Community ecology package. R package version 2.0-4. [ Links ]
32. Paglia, A. P. et al. 2012. Lista Anotada dos Mamíferos do Brasil. 2ed. Arlington: Conservation International. [ Links ]
33. Palmer, M. W. 1993. Putting things in even better order: the advantages of canonical correspondence analysis. Ecology 74:2215-2230. [ Links ]
34. Parry, L., J. Barlow, & C. A. Peres. 2007. Large-vertebrate assemblages of primary and secondary forests in the Brazilian Amazon. Journal of Tropical Ecology 23:653-662. [ Links ]
35. Peixoto, A. L., & A. Gentry. 1990. Diversidade e composição florística da Mata de Tabuleiro na Reserva Florestal de Linhares (Espírito Santo, Brasil). Revista Brasileira de Botânica 13:19-25. [ Links ]
36. Prevedello, J. A., P. Ferreira, B. S. Papi, D. Loretto, & M. V. Vieira. 2008. Uso do espaço vertical por pequenos mamíferos no Parque Nacional Serra dos Orgãos, RJ: Um estudo de 10 anos utilizando três métodos de amostragem. Espaço & Geografia 11:35-58. [ Links ]
37. Silvius, K. M., & J. Fragoso. 2003. Red‐rumped Agouti (Dasyprocta leporina) Home range use in an Amazonian forest: implications for the aggregated distribution of forest trees. Biotropica 35:74-83. [ Links ]
38. Srbek-Araújo, A. C, & A. G. Chiarello. 2005. Is camera-trapping an efficient method for surveying mammals in Neotropical forests? A case study in south-eastern Brazil. Journal of Tropical Ecology 21:121-125. [ Links ]
39. Srbek-Araújo, A. C., & A. G. Chiarello. 2013. Influence of camera-trap sampling design on mammal species capture rates and community structures in southeastern Brazil. Biota Neotropica 13:51-62. [ Links ]
40. Stein, A., K. Gerstner, & H. Kreft. 2014. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecology letters 17:866-880. [ Links ]
41. Tews, J. et al. 2004. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography 31:79-92. [ Links ]
42. Vieira, E. M., & E. L. Monteiro-Filho. 2003. Vertical stratification of small mammals in the Atlantic rain forest of south-eastern Brazil. Journal of Tropical Ecology 19(5):501-507. [ Links ]
43. Virgós, E., 2001. Relative value of riparian woodlands in landscapes with different forest cover for medium-sized Iberian carnivores. Biodiversity and Conservation 10:1039-1049. [ Links ]
44. Voss, R. S., & L. H. Emmons. 1996. Mammalian diversity in Neotropical lowland rainforests: a preliminary assessment. Bulletin American Museum of Natural History 230:1-115. [ Links ]
Large- and medium-sized mammal species recorded at Vale Natural Reserve, Espírito Santo state of Brazil. We presented the photographic rates (F), the vegetation types (SF: sandy soil forest; NG: natural grassland; CF: coastal forest; E: Ecotone) and regions (N: northern; S: south; W: west) in which the species were recorded from May 2013 to June 2014 *exotic species; ¹endemic species to the Atlantic Forest.



