










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMastozoología neotropical
versión impresa ISSN 0327-9383versión On-line ISSN 1666-0536
Mastozool. neotrop. vol.25 no.1 Mendoza jun. 2018
ARTÍCULO
Vanished Lagoa Santa: rodent species loss in the last two centuries
Natália L. Boroni¹, Ulyses F. J. Pardiñas² and Gisele Lessa¹
¹ Museu de Zoologia João Moojen, Departamento de Biologia Animal, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil. [Correspondence: <natalia_boroni@hotmail.com>]
² Instituto de Diversidad y Evolución Austral (IDEAus-CONICET), Puerto Madryn, Chubut, Argentina.
Recibido 16 junio 2017.
Aceptado 18 enero 2018.
Editor asociado: A Noss
ABSTRACT.
The extensive work developed by Peter Lund and other naturalists in the nineteenth century in Lagoa Santa Karst has generated a large volume of data on the extinct and extant local fauna. Despite these classic pioneer works, little work has been done about the fauna currently present in the region. Therefore, this study analyzes the richness and diversity of the modern fauna of small rodents of the Lagoa Santa region and compares the results with the historical data of Lund and Winge. We collected owl pellets from calcareous caves in Lagoa Santa, Cordisburgo and Sete Lagoas. These pellets were divided in two categories for comparison: superficial (collected on the surface of the cave floor) and stratified (collected in small excavations in the cave floor). We identified a total of 18 rodent species: 13 Cricetidae, 3 Echimyidae and 2 Muridae. Richness considering the minimum number of individuals (MNI) was greater in stratified samples than in superficial samples, but the two sets of samples did not differ when considering rarefaction. According to the Shannon- Wiener and Pielou indices, the stratified samples were more diverse and with less species dominance. Nine species mentioned or described by Lund and Winge in the nineteenth century as inhabiting this region were absent in our samples. The proportion of small mammals in owl pellets in the modern samples was different from historical data, with predominance of Calomys spp. instead of Necromys lasiurus, as recorded by Lund using similar methods. It is likely that human activities in the region over these 180 years have influenced the fauna of Lagoa Santa, triggering local or regional extinctions.
RESUMO.
Lagoa Santa devastada: perda das espécies de roedores nos últimos dois séculos.
O extenso trabalho desenvolvido por Peter Lund e outros naturalistas no século XIX no Carste de Lagoa Santa gerou um enorme volume de dados sobre a fauna local extinta e existente. Apesar destes trabalhos pioneiros clássicos, pouco se sabe sobre a fauna atual na região. Portanto, este estudo analisa a riqueza e diversidade da fauna moderna de pequenos roedores da região de Lagoa Santa e compara os resultados com os dados históricos obtidos por Lund e Winge (1836). Para isso, foram coletadas pelotas de coruja em cavernas calcáreas em Lagoa Santa, Cordisburgo e Sete Lagoas. As amostras foram divididas em duas categorias: superficial (coletadas na superfície do chão das cavernas) e estratificada (coletadas em pequenas escavações do chão das cavernas). Um total de 18 espécies de roedores foram identificados: 13 Cricetidae, 3 Echimyidae e 2 Muridae. A riqueza foi maior nas amostras estratificadas quando comparadas no mesmo MNI com amostras superficiais, mas não diferiram na análise de rarefação.De acordo com os índices Shannon-Wiener e Pielou, a estratificada foi considerada mais diversificada e com menor predominância de espécies. Nove espécies mencionadas ou descritas por Lund e Winge no século XIX como habitando a região foram ausentes em nossas amostras. A proporção de pequenos mamíferos nas amostras modernas foi diferente dos dados históricos, com predominância de Calomys spp. em vez de Necromys lasiurus, conforme registrado por Lund por métodos similares. É provável que as atividades humanas na região ao longo desses 180 anos tenham influenciado a fauna de Lagoa Santa, provocando extinções locais ou regionais.
Key words: Cerrado; Environmental degradation; Herluf Winge; Owl pellets; Peter Lund.
Palavras-chave: Cerrado; Degradação ambiental; Herluf Winge; Pelotas de coruja; Peter Lund.
INTRODUCTION
The karst region of Lagoa Santa in Minas Gerais has a great historical and scientific importance to paleontology and mammalogy in South America. This is mainly due to the work of Peter Wilhelm Lund in the nineteenth century, who collected and described a considerable diversity of fossil and living animals, including megafauna and human remains (Paula-Couto 1950; Voss & Myers 1991; Cartelle 2002). Lunds work is one of the first scientific mammal surveys conducted in Brazil and represents an accurate picture of the local fauna almost two hundred years ago (Cartelle 2002).
In his incursions, Lund found a large quantity of recent and fossil owl pellets in limestone caves, which allowed the description of many species of small mammals, particularly rodents (Paula-Couto 1950). This was the first small mammal survey using this methodology in South America; in fact, Lund carefully recorded materials from recent or fresh samples of pellets as well as those obtained from sediments. Later, all material collected by Lund was sent to Denmark and studied in European scientific circles (Ávila-Pires 1960), resulting in several papers on the Lagoa Santa fauna, not only by Lund, but by other naturalists as well, particularly Herluf Winge (Winge 1887). With this material, Winge described a considerable number of small mammals from cave sediments and recent deposits in the Lagoa Santa area (Voss & Myers 1991).
Much of the current knowledge regarding small mammals in South America is due to the study of barn owl pellets (see Pardiñas & Cirignoli 2002). The barn owl (Tyto furcata) is one of the main predatory birds of small mammals around the world (Taylor 1994). The barn owl regurgitates all undigested material in a rounded mass of fur and bones, which are known as pellets or egagropiles (Andrews 1990; Taylor 1994). These birds have high affinity for their shelters (e.g. caves), occupying them for long periods and documenting species assemblages in the pellets (Terry 2010a). Due to climatic conditions and environmental stability, these environments have a high preservation potential, which leads to the conservation of these deposits and eventual fossilization of bone material in the shelters (e.g. Hadly 1996; Grayson 2000). The barn owl is characterized as a generalist predator and can be considered a good sampler of the small mammal community at local and regional scales (Andrews 1990; Heisler et al. 2016). From assemblages derived from owl pellets it is possible to study the small mammal community, environmental changes (from present to ancient times), and anthropogenic impacts, like deforestation, on local diversity (Terry 2010a; Pardiñas et al. 2012).
The karst area from central Minas Gerais is one of the regions in Brazil with the most information regarding the living and extinct fauna, and the type locality of several species, especially small mammals. Despite considerable data secured by Lund about small mammals derived from owl pellets, no posterior studies were carried out in Lagoa Santa region using this approach. The present paper aims to compare, for the first time, the nineteenth century historical record rodent assemblage in the Lagoa Santa region with the modern assemblage, as defined by recent samples of owl pellets collected on the surface and in excavations. Special attention was directed to detect qualitative and quantitative changes in the two assemblages and potential correlations with environmental transformations in the region during the last 180 years.
MATERIALS AND METHODS
Study area
This study was conducted in the karst region of central Minas Gerais, by manually collecting egagropiles in four caves in three different localities (Fig. 1): Salitre Cave (19º 07′ 17″ S and 44º 28′ 24″ W; SC, thereafter), Toca do Lixo (19º 33′ 30″ S, 43º 57′ 03″ W; TL, thereafter), Mariposas Cave (19° 33′ 38″ S, 43° 57′ 39″ W; MC, thereafter), and Mata Grande (19º 30′ 02″ S, 44º 15′ 59″ W; MG, thereafter). SC is very near the famous Maquiné Cave, one of the first caves studied by Lund (1836) in the Peter Lund Natural Monument, located in Cordisburgo, where sixteen caves have been explored (Travassos 2010) (Fig. S1). TL and MC are two small caves in Sumidouro State Park, which protects 100 registered caves and 170 historic and prehistoric archaeological sites (IEF 2008). Finally, MG, in the city of Sete Lagoas is near Rei do Mato Cave, another famous cave and important archaeological site, with cave paintings and ceramic artifacts.
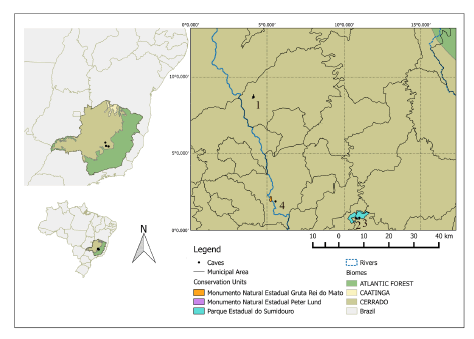
Fig. 1. Collecting localities of owl pellets samples: (1) Salitre cave, Peter Lund Natural Monument, Cordisburgo; (2) Mariposa cave and (3) Toca do Lixo, Sumidouro State Park, Pedro Leopoldo / Lagoa Santa; (4) Mata Grande cave, Sete Lagoas. Source: IBGE.
The climate in the region is Aw tropical humid, according to the Köppen classification (IEF 2008; Travassos 2010). The annual average temperature in the region is 21.7 ºC with an annual rainfall of 1291 mm (IEF 2008, Travassos 2010).
The vegetation is typical of the Cerrado biome, but with the occurrence of some enclaves similar to Caatinga in the limestone outcrops, known as the Deciduous Seasonal Forest or "Mata Seca" (Piló 1998). At present, natural vegetation is restricted to only a few patches—most of them disturbed (Berbert-Born 2002); the landscape matrix is dominated by pastures, crops and Eucalyptus plantations (Travassos 2010).
Data from owl pellets
Modern rodent community
Modern samples were collected from fresh and disaggregated owl pellets from SC (collected in 2009), TL (collected in 2010), MC (collected in 2010) and MG (collected in 2011). These samples vary in quantity, with hundreds to thousands of rodent specimens. Modern samples are divided in two categories: superficial and stratified.
Modern superficial samples were swept from cave floors and cavities. Most of the material was already fragmented, with few whole pellets. These samples are accumulations over several months to years. Modern stratified samples were collected from a small excavation made in the floors of two caves. In the Salitre Cave, one excavation was performed at three different depths (layers) at the same point, each layer about 10 cm deep and 20 cm in diameter. In Toca do Lixo only one excavation was made (approx. 10 x 20 cm), because the soil was shallow over solid rock. MG and MC lacked any soil layer and excavation was not possible. Unfortunately, it was not possible to date these stratified samples, but we think it is reasonable to assume they represent several decades, since one fragment of modern ceramic was found together with the specimens in the second stratum excavation in Salitre Cave. Although the data were collected in different layers of stratified samples in Salitre Cave and they are not dated, they were analyzed as a single layer.
Fresh pellets typical of the barn owl (Tyto furcata) were identified at the collection sites, in addition to feathers and direct observations in some caves, indicating that the modern samples had originated from T. furcata pellets. Analysis of the stratified material also suggests the source to be T. furcata, because the bone material was well-preserved. Among the Strigiform birds, T. furcata is the species that causes the least damage to bones in the digestion process (Andrews 1990). Usually the bones in the T. furcata pellets are in good condition, with a relatively complete skeleton and longer non-fragmented bones (Andrews 1990). Barn owls generally eat small mammals weighing less than 200 g (Motta- Junior 2006), and in the samples small rodents less than 200 g predominated, corroborating the origin proposed for the material.
All samples were placed in the sun for dehydration, and then sieved to remove the sediment. Skulls, maxillas, molars and mandibles of rodents were manually separated. The identification of the rodent fragments was based on craniomandibular material by analyzing the cranial structures and morphology of the teeth using a stereomicroscope. For identification at the lowest possible taxonomic level, reference collections and specialized bibliography were used, although some fragments did not allow reliable identification to the species level, when they were very fragmented or did not preserve diagnostic features. In addition, some genera did not have all specimens identified to the species level due to their many morphological similarities. Nomenclature of the species was based on Wilson & Reeder (2005) and Patton et al. (2015). For Cricetidae rodent teeth, the nomenclature follows Reig (1977), while for Echimyidae it follows Emmons & Vucetich (1998). All the material collected was deposited in the Mammalogy collection of the João Moojen Museum of Zoology (Universidade Federal de Viçosa, Brazil) (Appendix 1). The material was deposited in batches by species and stratum per cave, so all material for each species of each stratum of each cave has one voucher number.
Historical rodent community
Lagoa Santa has a long history of research on living and fossil small rodents during the nineteenth century (Winge 1887; Paula-Couto 1950). These records were collected, for the most part, from 1835 to 1849 by Lund, and were divided in two categories in this study: Historical and Fossil. Historical data represent the community that was present almost two hundred years ago as observed in fresh and superficial pellets, in addition to live animals. Fossil data are the records found exclusively in the sediments of the caves and not described as living species in the Lagoa Santa region by Lund or Winge.
A bibliographic survey was conducted to obtain the actual synonymy of the names proposed by Lund and Winge (e.g. Paula-Couto 1950; Ávila-Pires 1960; Voss & Myers 1991; Wilson & Reeder 2005; Pardiñas et al. 2008, 2016; Pardiñas & Teta 2013, 2011a; b, Patton et al. 2105). These Historical and Fossil data were used to compare the rodent fauna observed in the Modern samples. Since the exact number of specimens collected and the sampling effort employed by Lund are not known, it is not possible to carry out statistical analyses comparing modern and historical samples. The comparisons in this paper are theoretical based on some abundance data of small mammals in pellets described by Lund in his memoirs (see Paula-Couto 1950). These comparisons are in the Discussion section below.
Data analysis
For each taxon the number of identified specimens (NISP) was determined as the sum of all the fragments of a taxon, and the minimum number of individuals (MNI) considered only the most frequent element of either the right or left side (Grayson 1973). In this study we only counted skulls, man dibles and maxiilae; no part of the post-skull was considered in the calculation. The percentage of occurrence of each rodent species was estimated by dividing the number of items collected from each species by the total number of collected items. Species richness was defined as the number of species collected in each cave and the abundance of each species was determined by MNI.
For the analysis of α diversity, the Shannon-Wiener (H), Simpson (D) and Pielou evenness (J) indexes were calculated (Magurran 2004). A Students t test was performed to assess the differences between the Shannon-Wiener and Simpson values obtained for Salitre Cave and Toca do Lixo samples, in pairs, following the methodology of Magurran (2004). To compare species abundance in the Modern superficial and Modern stratified samples, a chi-square test was performed (x²) for samples from the Salitre Cave and Toca do Lixo. These analyses were performed in the Past program (Hammer et al. 2001).
Rarefaction analyses were performed to compare the richness between the Modern superficial and Modern stratified samples, employing the Estimate program, version 9.1 (Colwell 2013). The statistical significance of these comparisons was evaluated using 95% confidence limits for the rarefaction curve. The sampling unit was considered to be the number of samples collected by cave and stratum. The sampling effort was considered to be the sum total of MNI by cave and stratum.
In these statistical treatments (rarefaction and α diversity), Calomys spp. and Oligoryzomys spp. were considered only at the generic level. As most specimens of these genera were identified to the generic level, if we used only the specimens identified to the species level, we could underestimate the abundance of these species.
RESULTS
Six modern samples originating from the pellets of T. furcata were analyzed from four different caves: four from the cave floor surfaces (Superficial) and the other two from excavations (Stratified). These samples included a total of 8658 identifiable items (NISP) of small rodents, comprising skulls, maxilla and mandible fragments. The minimum number of individuals (MNI) was 2871, corresponding to 15 genera and 18 species of rodents (Table 1, Figs. S2, S3).
Table 1 Rodentia found in owl pellets in Salitre Cave (SC), Peter Lund Natural Monument, Cordisburgo; Toca do Lixo (TL) and Mariposa Cave (MC), Sumidouro State Park, Pedro Leopoldo / Lagoa Santa; Mata Grande Cave (MG), Sete Lagoas. MNI = minimum number of individuals. NISP = sum of all the fragments of the taxon. Modern superficial: SC-A, TL-A, MG, MC. Modern stratified: SC-S, TL-S. (%) Percentage of occurrence.
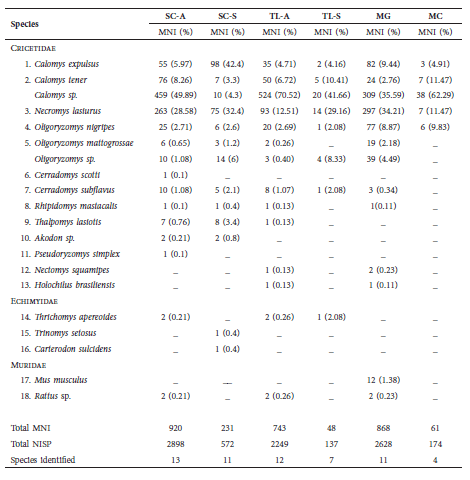
Calomys tener, C. expulsus, Necromys lasiurus and Oligoryzomys nigripes were common in all the samples. In the Modern superficial sample from Salitre Cave (MNI = 920), 13 rodent species were identified, of which Cerradomys scotti and Pseudoryzomys simplex stand out as exclusive to this sample. In the Modern stratified sample from the same cave (MNI = 231), 11 species were recorded, of particular note being two species of spiny rats, Carterodon sulcidens and Trinomys setosus. In the Toca do Lixo Modern superficial (MNI = 743) and Modern stratified (MNI = 48) samples, 12 and 7 species were observed, respectively. Nectomys squamipes and Holochilus brasiliensis were found only in Toca do Lixo and Mata Grande Cave. In Mariposa Cave (MNI = 61), only four species were recorded. In Mata Grande Cave (MNI = 868), 11 species were identified; this was the only sample that registered the murine rodent, Mus musculus.
Comparisons between the Modern superficial and Modern stratified
Richness and abundance varied among the samples, being greater in the Modern superficial samples (considering C. tener and C. expulsus as Calomys sp., and O. nigripes and O. mattogrossae as Oligoryzomys sp.). In the superficial samples of SC and TL, higher species richness was observed (11 and 10 species, respectively) than in the stratified samples (9 and 5 species, respectively), although the sample size was much higher in the superficial sample (MNI SC = 920 and TL = 743) than stratified (MNI SC = 231 and TL = 48). Cerradomys scotti, P. simplex, N. squamipes and H. brasiliensis have only been recorded in the superficial samples. Trinomys setosus and C. sulcidens were exclusive to the stratified samples of SC. However, rarefaction curves showed no significant differences in species richness among the samples (Figs. S4). When comparing MNI across samples, greater diversity is seen in the Modern stratified than in the Modern superficial samples (Figs. S5, S6). But the confidence intervals (95% CI) were high and overlapped; thus, these samples were not distinct from each other in terms of species richness. The rarefaction curves also do not reach an asymptote, therefore, the sampling cannot be considered exhaustive, especially in the case of the stratified samples.
According to the Shannon-Wiener index (H), the Modern superficial samples from SC and TL were less diverse than Modern stratified samples (p < 0.006 and p = 0.001, respectively) (Table 2). In relation to species abundance, the Pielou evenness index (J) shows that the Modern stratified data are more uniform in abundance across the species. Modern stratified data also demonstrate greater diversity and lower species dominance, with Simpson index (D) values closest to one (p < 0.0032 for SC and p < 0.0001 for TL cave).
Table 2 Diversity indices for modern owl pellets samples in Salitre Cave (SC), Peter Lund Natural Monument, Cordisburgo; Toca do Lixo (TL) and Mariposa Cave (MC), Sumidouro State Park, Pedro Leopoldo / Lagoa Santa; Mata Grande Cave (MG), Sete Lagoas. Modern superficial: SC-A, TL-A, MG, MC. Modern stratified: SC-S, TL-S. N = Sample size (MNI); S = Total species; H = Shannon-Weaver; J = Equitability of Pielou; D = Simpsons dominance.

The fauna from the Superficial and Stratified modern samples are significantly different (x2 SC = 57.589, x² TL = 23.561, p < 0.05). For the TL cave, the abundance of Necromys (p=0.002) the difference in the samples (Table 3). Several genera like Calomys (p = 0.013), Oligoryzomys (p = 0.001), Thalpomys (p = 0.001) and Trinomys (p = 0.046) differ significantly in abundance between Superficial and Stratified modern samples from SC.
Table 3 Comparison of rodent species abundance between superficial and stratified modern owl pellet samples in Salitre Cave (SC), Peter Lund Natural Monument, Cordisburgo; Toca do Lixo (TL), Sumidouro State Park, Pedro Leopoldo / Lagoa Santa. Modern superficial: SC-A, TL-A. Modern stratified: SC-S, TL-S. *Significant values (p < 0.05).

DISCUSSION
This survey found a high species richness (S = 18), greater than other short- and long-term studies using fresh owl pellets in the Cerrado, Pantanal and Atlantic Forest (Motta-Junior & Talamoni 1996; Motta-Junior & Alho 2000; Bonvicino & Bezerra 2003; Escarlate-Taveres & Pessoa 2005; Roda 2006; Scheibler & Christoff 2007; Bueno & Motta-Junior 2008; Magrini & Facure 2008; Souza et al. 2010; Rocha et al. 2011; Lemos et al. 2015). The results from this study indicate the most diverse sampling of pellets ever recorded for these biomes (Table S1), except for Peter Lunds survey.
The majority of the species found are common in the Cerrado Biome—inhabiting open areas such as grasslands, gallery forests, and woodland savanna—for example C. expulsus, C. tener, N. lasiurus, T. lasiotis (Patton et al. 2015). Despite reports of sympatry among Calomys tener, C. expulsus and C. cerqueirai in the Lagoa Santa region (Bonvicino et al. 2010), in this study only C. expulsus and C. tener were identified. Although C. tener is slightly smaller than C. expulsus in some cranial features, the two species are difficult to separate, especially when the skull is fragmented, because the molars do not differ in discrete characters. Calomys cerqueirai exhibits many morphological similarities with C. expulsus, showing differences mainly in the external characteristics and some descriptive skull measurements, but with great overlap (see Bonvicino et al. 2010). Therefore, it was not possible to differentiate between the two species in this work. Some specimens collected may in fact be Calomys cerqueirai, which was described in the Lagoa Santa region (Bonvicino et al. 2010). A similar situation occurred with Akodon sp. specimens. Akodon cursor and A. montensis are sympatric in the Lagoa Santa region (see Geise et al. 2001), but the specimens from the pellets were too fragmented to differentiate the species (see Geise et al. 2005; Gonçalves et al. 2007). More complete specimens would need to be collected for a more accurate taxonomic identification.
Among the less frequent species in the sample are rodents with semi-aquatic habitat (H. brasiliensis, P. simplex and N. squamipes), the arboreal Rhipidomys mastacalis, and the spiny rat Thrichomys apereoides. Probably they are not well represented in the pellet samples because of their habitat preferences and medium to large body size. According to Taylor (1994), barn owls usually hunt smaller prey. Studies in Brazil have shown that they prefer rodents weighing an average of 10 to 40 g (Bonvicino & Bezerra 2003; Scheibler & Christoff 2007; Bueno & Motta-Junior 2008). In other studies, especially in flooded areas, an increase in the mass of prey is observed, represented by larger specimens such as Holochilus sp. (Roda 2006; Rocha et al. 2011).
Comparison of the richness and diversity in the Modern community: Superficial and Stratified data
The proportions of the faunistic elements differ between the Superficial and Stratified samples. In the deeper layer, there is a decrease in the proportion of Calomys and an increase in N. lasiurus and other species like Oligoryzomys sp., T. lasiotis and T. setosus (see Table 3). Habitat specialist rodents (such as T. lasiotis and T. setosus) are more abundant in the stratified samples, whereas the superficial samples have greater representation of the more generalist and open habitat species (C. tener and C. expulsus). In other studies, when comparing the subfossil and modern samples, changes in the diversity of small mammals can be observed, with the most recent samples showing an impoverishment of owl prey species richness, and an increase in the presence of certain opportunistic taxa (Pardiñas et al. 2000; Terry 2010b).
Faunal composition changes in owl pellets could reflect the variation in prey availability or different foraging seasons (Lyman et al. 2003). However, as we lack data on the present abundance of the local rodents, we cannot directly assess whether prey availability has changed. Moreover, these samples are aggregations of pellets over several years, and we could not analyze annual or seasonal variations in prey. Samples incorporated in the stratigraphic record should be understood as samples accumulated over time and, therefore, representing the general trend over several years and no single moments in time (Fürsich & Aberhan 1990).
Another explanation for the change in the proportion of rodents could be the differences in sample sizes. Considering the superficial sample of the Salitre Cave (MNI 920) and the stratified sample (MNI 231) at the same site, 11 and 9 species are represented respectively, but they did not differ in richness according to the rarefaction analysis (Fig. S5). Nonetheless, according to the Shannon diversity index, the stratified sample is more diverse and abundant (H = 1.217) than the superficial sample (H = 0.93) in the Salitre Cave. This is contrary to the trend observed through many surveys, that diversity increases with increasing sample size (Grayson 1984).
Barn owls specialize in catching small mammals, but they are also opportunistic in the sense that they hunt a wide variety of species, especially those that are more abundant (Marti 1988). The decrease of Calomys sp. and the increase of N. lasiurus in the Stratified sample may be attributable to the greater availability of N. lasiurus in the past. Habitat changes in the region have probably caused population declines of some habitat specialist species, like T. lasiotis and C. sulcidens, since it is known that some species are more vulnerable to habitat loss and fragmentation than others (Bright 1993). Habitat changes and habitat loss, along with other factors, may be related to a decrease in the diversity of small mammal species in the barn owls diet during the twentieth century (Love et al. 2000).
Unfortunately, it was not possible to maintain better stratigraphic control and date the samples to establish the age of the deposits. We know that the stratified samples are not very old; and probably represent no more than the last century. Nevertheless, we found that the older samples tend to have a rodent fauna (abundance) more similar to that described by Lund in the nineteenth century.
Comparison of Modern and Historical community
The small mammals in the pellet samples reflect local wildlife in the landscape (Hadly 1999; Terry 2010a). Several species listed by Lund (1836, 1837, 1838, 1839) and Winge (1887) as living in Lagoa Santa were not found in the modern owl pellets collected in the region (Table 4). Winge and Lund recognized 25 species of Cricetidae rodents in the region, nine species of Echimyidae and two Muridae between the Historical materials and Fossil found on the floor in the caves (Winge 1887; Voss & Myers 1991). Of the 36 total species, 13 were found only in the sediments of caves, not being reported as living by Winge and Lund, and 23 were described as living forms. In this study, we only found 16 species in owl pellets (not considering C. scotti and O. mattogrossae, species not recognized at the time of Winges publication [1887]), and two of these were recorded only in the Stratified samples. Modern species richness is lower than the Historical assemblage described by Lund and Winge, with few or no arboreal and fossorial species, and relative abundances differ as well between the two periods.
Table 4 Comparison of Historical community (specimens of rodents collected in the Lagoa Santa region in the XIX century) and Modern community. P = specimens found in the owl pellets in the caves; S = specimens with skin collected.


In 1838, five species of spiny rats were described by Lund in this area (Lund 1838, p. 249). Carterodon sulcidens was known to Lund only by the remains found in caves (Lund 1837, p. 167). Today, C. sulcidens is Endangered (EN) in Minas Gerais (COPAM 2010). Meanwhile, Clyomys laticeps was rarely found in the region, while T. setosus and T. apereoides were more common (Lund 1837, p. 167; Lund 1838, p. 231, 262). The genus Phyllomys was later described from a few skeletal fragments, found in some caves located north of Lagoa Santa (Lund 1838b, p. 227). Today, the genus has two Endangered (EN) species in Minas Gerais (P. brasiliensis and P. lundi) and four in Brazil (P. thomasi, P. unicolor, P. brasilensis and P. lundi) (COPAM 2010, MMA 2014).
Callistomys sp. and Kannabateomys amblyonyx were only found in the Fossil material (Winge 1887), not occurring presently in the Lagoa Santa region, which may indicate a recent reduction in the geographic distribution of both genera (Voss & Myers 1991; Emmons & Vucetich 1998), or the difficulty of capturing these animals whose behavior makes them less accessible (Voss & Myers 1991). A recent study recorded K. amblyonyx about 72 km southeast of Sumidouro State Park, in a transition area of Atlantic Forest and Cerrado (Unidade Ambiental Peti, São Gonçalo do Rio Abaixo) (Da Silva et al. 2012). Euryzygomatomys mordax and Dicolpomys fossor, rodents with fossorial habits, were found only in fossil material and date from the late Pleistocene / Holocene (Hadler et al. 2008).
Among the Echimyidae listed by Lund, only T. apereoides was frequent in the Modern fauna. Carterodon sulcidens, a rodent very common and abundant in pellets in the nineteenth century (Lund 1836, p. 126), presented one mandible in Stratified samples of the SC. There are hardly any recent records for this species (Bezerra et al. 2011).
Several Cricetidae—Thaptomys nigrita, Bibimys labiosus, Kunsia tomentosus, Hyaleamys laticeps and Oxymycterus dasytrichus—described by Lund (1839b, p.291) and Winge (1887) as living fauna, were not found in our sample, although T. nigrita and O. dasytrichus (identified as O. roberti) have been collected near the study area (Ávila-Pires 1960; Leal et al. 2008). The lack of recent records of K. tomentosus in the Lagoa Santa region may be related to the intense landscape modification by human activities (Pardiñas et al. 2008).
Several species cited by Lund and Winge as living in the Lagoa Santa region (Lund 1839b, p. 291; Winge 1887) were found in this study and in nearby regions: Oligoryzomys sp., C. tener, R. mastacalis, N. squamipes, and H. brasiliensis (Ávila-Pires 1960; Leal et al. 2008).
Winge (1887) reported that P. simplex, C. subflavus and A. cursor were common in fresh owl pellets and in fossil deposits in the caves. However, in our studies P. simplex was extremely rare, with only a single skull fragment found in the Modern data, and Akodon represented just 0.21% of the modern rodents of the SC.
In Modern samples, Calomys sp. represents 64% of the SC fauna and 80% of the sample of TL, and N. lasiurus was the second most abundant species, whereas in 1836, Lund reported that N. lasiurus was the most abundant rodent species in owl pellets, representing about 80% of individuals (Table 5).
Table 5 Proportion of specimens in owl pellets described by Lund (1836), collected in Salitre cave (SC, 2009) and Toca do Lixo (TL, 2011). Modern superficial: SC-A, TL-A. Modern stratified: SC-S, TL-S.
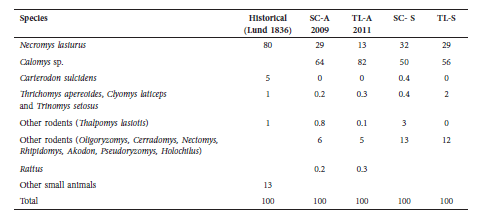
In Modern samples, N. lasiurus abundance increased in the stratified samples with depth, but the difference in the proportion was only significant in TL Superficial vs. Stratified (p = 0.003). The same pattern was observed for T. lasiotis, being more abundant in the stratified samples. This species has also been collected in nearby locations such as Serra do Cipó and Serra das Sempre Vivas (Leal et al. 2008), and this species is considered Vulnerable (VU) in Brazil (MMA 2014).
Some Cricetidae found exclusively as Fossils from the caves (Lund 1839, p. 291; Winge 1887)—for example Sooretamys angouya, Blarinomys breviceps and Delomys plebejus— were not found in our samples. The last record for these species close to Sumidouro State Park was from Conceição do Mato Dentro, 50 km away from Lagoa Santa (Ávila-Pires 1960). The B. breviceps collected by Lund was fossil dated from the Pleistocene. It is probably extinct in the locality, and its existence in the region in the past could have been related to the more mesic environment (Silva et al. 2003). Meanwhile, D. plebejus should be considered a nomen dubium, as its equivalence to any of the living species is indeterminate (Voss 1993).
Environmental degradation
The differences in richness and abundance are important and may not be explained only by sample size or taphonomic bias. These changes in the small mammal proportions and possible local population declines can probably be related to environmental changes over the past 180 years. The fragmentation of the natural vegetation was undoubtedly the main factor contributing to the decline of medium and large mammals in the region (Trolle et al. 2007). During the last century, the Lagoa Santa region has also suffered bird population declines and the disappearance of many species recorded by Lund, Reinhardt and Burmeister. Over the years, the bird community, originally so diverse, has been replaced by a few generalist species, none of which were common in the last century (Christiansen & Pitter 1997).
Lund (1838b, p. 209, 210) already reported impacts caused by farming and exploitation of saltpeter, including possible extinctions of flora. Today, the Cerrado remnants are restricted to the dolines and rock outcrops, surrounded by pastures (Berbert-Born 2002; Travassos 2010). Studies on modern, historical and fossil rodents using owl pellets in a degraded area in North America showed variation in the richness and abundance in comparison with a preserved area, suggesting quick changes in the ecological baseline in the region. Thus, it is likely that anthropogenic processes there have led to a change in the vegetation and in the rodent fauna (Terry 2010b), and our study found similar patterns in the Lagoa Santa region. Such landscape modifications over the last 180 years are likely to have resulted in homogenization of habitats and these changes were reflected in T. furcata diets, with a marked increase of Calomys (C. expulsus and C. tener). Calomys species have been described as opportunistic and dominant in the anthropogenic environments (e.g. Bellocq 1990; Mills et al. 1991; Pardiñas et al. 2000). Pardiñas et al. (2000) have observed a similar pattern, with the dispersion and increased abundance of Calomys (C. laucha and C. musculinus) to the detriment of other small mammals, in human-modified landscapes.
These changes in rodent abundance and geographic ranges over the past 180 years could also be explained by climate change, although little is known about how rodent communities respond to climate change through time (Hadly 1996; Grayson 2000; Blois & Hadly 2009; Blois et al. 2010). For small rodents, a warmer climate may cause a local increase in abundance of species tolerant to higher temperatures (Terry et al. 2011). Unfortunately, no analysis has been done with meteorological data for this region and period to infer any changes.
CONCLUSIONS
In this paper, a great richness of the rodents in the central karst region of Minas Gerais was revealed by owl pellets, with Calomys tener and C. expulsus prevalent in the modern samples. The Modern rodent community is less rich than the Historical community described by Lund and Winge. This lower diversity of rodents currently observed in the Lagoa Santa region may result from environmental degradation in the last century. Despite the increase in studies about the rodent fauna, Brazil still presents gaps in information about historical and fossil communities. Studies should also emphasize digging sequences (Stratified samples) to understand the changes in the small mammal communities over the centuries. Such studies would generate more complete conclusions about how anthropogenic degradation affects extinction processes, population declines and species invasion.
ACKNOWLEDGMENTS
We are grateful to: Leonardo Dias and Camilo Arias for their valuable assistance in the field work; Sumidouro State Park staff, Peter Lund Natural Monument and Mining Company Mata Grande, for permitting the use of their facilities for sample collections; Marcelo Weksler, Rodolfo Stumpp, Fernando Perini, Andrew Noss and the anonymous reviewers for their pertinent comments on improving this work; Rodolfo Stump for assistance in the analyses and figures.
LITERATURE CITED
1. Andrews, P. 1990. Owls, caves and fossils: Predation, preservation, and accumulation of small mammal bones in caves, with an analysis of the Pleistocene Cave faunas from Westbury-sub-Mendip, Somerset, UK. University of Chicago Press, Chicago. [ Links ]
2. Ávila-Pires, F. D. De. 1960. Roedores colecionados na região de Lagoa Santa, Minas Gerais, Brasil. Arquivos do Museu Nacional 50:25-46. [ Links ]
3. Bellocq, M. I. 1990. Composición y variación temporal de la dieta de Tyto alba en ecosistemas agrarios pampeanos, Argentina. Vida Silvestre Neotropical 2:32-35. [ Links ]
4. Berbert-Born, M. 2002. Carste de Lagoa Santa, MG: Berço da paleontologia e da espeleologia brasileira. Sítios Geológicos e Paleontológicos do Brasil (C. Schobbenhaus, D. A. Campos, E. T. Queiroz, M. Winge & M. L. Berbert-Born eds.). Comissão Brasileira de Sítios Geológicos e Paleobiológicos (SIGEP), Brasília. [ Links ]
5. Bezerra, A. M. R., J. Marinho-Filho, & A. P. Carmignotto. 2011. A Review of the distribution, morphometrics, and habit of Owls Spiny Rat Carterodon sulcidens (Lund, 1841) (Rodentia: Echimyidae). Zoological Studies 50:566-576. [ Links ]
6. Blois, J. L., & E. A. Hadly. 2009. Mammalian response to Cenozoic climatic change. Annual Review of Earth and Planetary Sciences 37:181-208. [ Links ]
7. Blois, J. L., J. L. Mcguire, & E. A. Hadly. 2010. Small mammal diversity loss in response to late-Pleistocene climatic change. Nature 465:771-774. [ Links ]
8. Bonvicino, C. R., & A. M. Bezerra. 2003. Use of regurgitated pellets of barn owl (Tyto alba) for inventorying small mammals in the Cerrado of central Brazil. Studies on Neotropical Fauna and Environment 38:37-41. [ Links ]
9. Bonvicino, C. R., J. A. Oliveira, & R. Gentile. 2010. A new species of Calomys (Rodentia: Sigmodontinae) from Eastern Brazil. Zootaxa 25:19-25. [ Links ]
10. Bright, P. W. 1993. Habitat fragmentation - problems and predictions for British mammals. Mammal Review 23:101-111. [ Links ]
11. Bueno, A. A., & J. C. Motta-Junior. 2008. Small mammal prey selection by two owl species in southeastern Brazil. Journal of Raptor Research 42:248-255. [ Links ]
12. Cartelle, C. 2002. Peter W. Lund, a naturalist of several sciences. Lundiana 3:83-85. [ Links ]
13. Christiansen, M. B., & E. Pitter. 1997. Species loss in a forest bird community near Lagoa Santa in southeastern Brazil. Biological Conservation. 80:23-32. [ Links ]
14. Colwell, R. K. 2013. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9.1.0. <http://purl.oclc.org/estimates> [ Links ].
15. COPAM - Conselho de Política Ambiental. 2010. Deliberação Normativa COPAM nº 147, de 30 de abril de 2010: Aprova a Lista de Espécies Ameaçadas de Extinção da Fauna do Estado de Minas Gerais. Diário do Executivo do Estado de Minas Gerais. [ Links ]
16. Da Silva, F. A., C. L. de Assis, R. A. da Silva, V. C. Antunes, G. Lessa, & F. M. Quintela. 2012. Distribution and conservation of the bamboo rat Kannabateomys amblyonyx (Rodentia, Echimyidae) in Minas Gerais State, Brazil. Neotropical Biology and Conservation 7:21-25. [ Links ]
17. Emmons, L. H., & M. G. Vucetich. 1998. The Identity of Winges Lasiuromys villosus and the description of a new genus of Echimyid Rodent (Rodentia: Echimyidae). American Museum Novitates 3223:1-12. [ Links ]
18. Escarlate-Tavares F., & L. M. Pessôa. 2005. Bats (Chiroptera, Mammalia) in barn owl (Tyto alba) pellets in northern Pantanal, Mato Grosso, Brazil. Mastozoología Neotropical 12:61-67. [ Links ]
19. Fürsich, F. T., & M. Aberhan. 1990. Significance of time averaging for palaeocommunity analysis. Lethaia 23:143-152. [ Links ]
20. Geise, L., D. Moraes, & H. S. da Silva. 2005. Morphometric differentiation and distributional notes of the three species of Akodon (Muridae, Sigmodontinae, Akodontini) in the Atlantic Coastal area of Brazil. Arquivos do Museu Nacional 63:63-74. [ Links ]
21. Geise, L., M. F. Smith, & J. L. Patton. 2001. Diversification in the genus Akodon (Rodentia: Sigmodontinae) in southeastern South America: mitochondrial dna sequence analysis. Journal of Mammalogy 82:92-101. [ Links ]
22. Gonçalves, P. R., P. Myers, J. F. Vilela, & J. A. Oliveira. 2007. Systematic of species of the genus Akodon (Rodentia: Sigmodontinae) in Southeastern Brazil and implications for the biogeography of the Campos de Altitude. Miscellaneous publications University of Michigan Museum of Zoology 197:1-24. [ Links ]
23. Grayson, D. K. 1973. On the methodology of faunal analysis. American Antiquity 38:432-439. [ Links ]
24. Grayson, D. K. 1984. Quantitative zooarchaeology. Academic Press, Orlando. [ Links ]
25. Grayson, D. K. 2000. Mammalian responses to middle Holocene climatic change in the Great Basin of the western United States. Journal of Biogeography 27:181-192. [ Links ]
26. Hadler, P., D. H. Verzi, M. G. Vucetich, J. Ferigolo, & A. M. Ribeiro. 2008. Caviomorphs (Mammalia, Rodentia) from the Holocene of Rio Grande do Sul State, Brazil: systematics and paleoenvironmental context. Revista Brasileira de Paleontologia 11:97- 116. [ Links ]
27. Hadly, E. A. 1996. Influence of late-holocene climate on northern Rocky Mountain mammals. Quaternary Research 46:298-310. [ Links ]
28. Hadly, E. A. 1999. Fidelity of terrestrial vertebrate fossils to a modern ecosystem. Palaeogeography, Palaeoclimatology, Palaeoecology 149:389-409. [ Links ]
29. Hammer, Ø., D. A. T. Harper, & P. D. Ryan. 2001. PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron. 4:1-9. <http://palaeo-electronica.org/2001_1/past/ issue1_01.htm> [ Links ].
30. Heisler, L. M., C. M. Somers, & R. G. Poulin. 2016. Owl pellets: a more effective alternative to conventional trapping for broad-scale studies of small mammal communities. Methods in Ecology and Evolution 7:96-103. [ Links ]
31. IEF - Instituto Estadual de Floresta. 2008. Parque Estadual do Sumidouro: Plano de Manejo. Technical report unpublished. IEF/Gheosfera Consultoria Ambiental. [ Links ]
32. Leal, K. P., I. R. Batista, F. L. Santiago, C. G. Costa, & E. M. Câmara. 2008. Mamíferos registrados em três unidades de conservação na Serra do Espinhaço: Parque Nacional da Serra do Cipó, Parque Nacional das Sempre Vivas e Parque Estadual da Serra do Rola-moça. Sinapse Ambiental 5:40-50. [ Links ]
33. Lemos, H. D. M., C. A. O. Silva, F. D. M. Patiu, & P. R. Goncalves. 2015. Barn owl pellets (Aves: Tyto furcata) reveal a higher mammalian richness in the Restinga de Jurubatiba National Park, Southeastern Brazil. Biota Neotropical 15:1-9. [ Links ]
34. Love, R. A., C. Webbon, D. E. Glue, & S. Harris. 2000. Changes in the food of British barn owls (Tyto alba) between 1974 and 1997. Mammal Review 30:107-129. [ Links ]
35. Lund, P. W. 1836. Cavernas existentes no calcário do interior do Brasil, contendo algumas delas ossadas fósseis. Primeira memória. 1950. Peter Wilhelm Lund: memórias sobre a paleontologia brasileira (C. P. Couto, ed.). Instituto Nacional do Livro, Rio de Janeiro. [ Links ]
36. Lund, P. W. 1837. Segunda memória sobre a fauna das cavernas. Mamíferos. 1950. Peter Wilhelm Lund: memórias sobre a paleontologia brasileira (C. P. Couto. ed.). Instituto Nacional do Livro, Rio de Janeiro. [ Links ]
37. Lund, P. W. 1838. Terceira memória sobre a fauna das cavernas. Continuação dos mamíferos. 1950. Peter Wilhelm Lund: memórias sobre a paleontologia brasileira (C. P. Couto, ed.). Instituto Nacional do Livro, Rio de Janeiro. [ Links ]
38. Lund, P. W. 1839. Suplemento as duas últimas memórias sobre o reino animal no Brasil, antes da última revolução do globo. 1950. Peter Wilhelm Lund: memórias sobre a paleontologia brasileira (C. P. Couto, ed.). Instituto Nacional do Livro, Rio de Janeiro. [ Links ]
39. Lyman, R. L., Power, E., & R. J. Lyman. 2003. Quantification and sampling of faunal remains in owl pellets. Journal of Taphonomy 1:3-14. [ Links ]
40. Magrini, L., & K. G. Facure. 2008. Barn owl (Tyto alba) predation on small mammals and its role in the control of hantavirus natural reservoirs in a periurban area in southeastern Brazil. Brazilian Journal of Biology 68:733-40. [ Links ]
41. Magurran, A. E. 2004. Measuring biological diversity. Blackwell Publishing, Oxford. [ Links ]
42. Marti, C. D. 1988. A long-term study of food-niche dynamics in the common barn-owl: comparisons within and between populations. Canadian Journal of Zoology 66:1803-1812. [ Links ]
43. Mills, J. N., B. A. Ellis, K. T. Mckee, J. I. Maiztegui, & J. E. Childs. 1991. Habitat associations and relative densities of rodent populations in cultivated areas of central Argentina. Journal of Mammalogy 72:470-479. [ Links ]
44. MMA - Ministério do Meio Ambiente. 2014. Instrução Normativa nº. 444 de 17 de dezembro de 2014. Lista Nacional Oficial de Espécies da Fauna Ameaçadas de Extinção. Diário Oficial da República Federativa do Brasil, Brasília. [ Links ]
45. Motta-Junior, J. C. 2006. Relações tróficas entre cinco Strigiformes simpátricas na região central do Estado de São Paulo, Brasil. Revista Brasileira de Ornitologia 14:359-377. [ Links ]
46. Motta-Junior, J. C., & C. J. Alho. 2000. Ecologia alimentar de Athene cunicularia e Tyto alba (Aves: Strigiformes) nas Estações Ecológica de Jataí e na Experimental de Luiz Antônio, SP. Estação Ecológica de Jataí (J. E. dos Santos & J. S. R. Pires, eds.). Rima editora, São Carlos. [ Links ]
47. Motta-Junior J. C., & S. A. Talamoni. 1996. Biomassa de presas consumidas por Tyto alba (Strigiformes: Tytonidae) durante a estação reprodutiva no Distrito Federal. Ararajuba 4:38-41. [ Links ]
48. Pardiñas, U. F. J., & S. Cirignoli. 2002. Bibliografía comentada sobre los análisis de egagrópilas de aves rapaces en Argentina. Ornitologia Neotropical 13:31-59. [ Links ]
49. Pardiñas, U. F. J., G. D'Elía, & P. Teta. 2008. Una introducción a los mayores sigmodontinos vivientes: revisión de Kunsia Hershkovitz, 1966 y descripción de un nuevo género (Rodentia: Cricetidae). Arquivos do Museu Nacional 66:509-594. [ Links ]
50. Pardiñas, U. F. J., L. Geise, K. Ventura, & G. Lessa. 2016. A new genus for Habrothrix angustidens and Akodon serrensis (Rodentia, Cricetidae): again paleontology meets neontology in the legacy of Lund. Mastozoología neotropical 23:93-115. [ Links ]
51. Pardiñas, U. F. J., G. J. Moreira, C. M. Garcia- Esponda, & L. J. Santis. 2000. Deterioro ambiental y micromamíferos durante el Holoceno en el nordeste de la estepe patagónica. Revista Chilena de Historia Natural 73:9-21. [ Links ]
52. Pardiñas, U. F. J., & P. Teta. 2011a. Fossil history of the marsh rats of the genus Holochilus and Lundomys (Cricetidae, Sigmodontinae) in southern South America. Estudios Geologicos 67:111-129. [ Links ]
53. Pardiñas, U. F. J., & P. Teta. 2011b. On the taxonomic status of the Brazilian mouse Calomys anoblepas Winge, 1887 (Mammalia, Rodentia, Cricetidae). Zootaxa 2788:38-44. [ Links ]
54. Pardiñas, U. F. J., & P. Teta. 2013. Taxonomic status of Mus talpinus Lund (Rodentia: Sigmodontinae) from the Quaternary deposits of Lagoa Santa, Minas Gerais, Brazil and its paleoenvironmental meaning. Mammalia 77:347-355. [ Links ]
55. Pardiñas, U. F. J., D. E. U. Sauthier, & P. Teta. 2012. Micromammal diversity loss in central-eastern Patagonia over the last 400 years. Journal of Arid Environments 85:71-75. [ Links ]
56. Patton, J. L., U. F. J. Pardiñas, & G. DElía (Eds.). 2015. Mammals of South America, Vol. 2 - Rodents. The University of Chicago Press, Chicago. [ Links ]
57. Paula-Couto, C. 1950. Peter Wilhelm Lund: memórias sobre a paleontologia brasileira. Instituto Nacional do Livro, Rio de Janeiro. [ Links ]
58. Piló, L. B. 1998. Morfologia cárstica e materiais constituintes: Dinâmica e evolução da Depressão Poligonal Macacos-Baú - Carste de Lagoa Santa, Minas Gerais. Dissertation. Universidade de São Paulo, São Paulo. [ Links ]
59. Reig, O. A. 1977. A proposed unified nomenclature for the enamelled components of the molar teeth of the Cricetidae (Rodentia). Journal of Zoology 181:227-241. [ Links ]
60. Rocha, R. G., E. Ferreira, Y. L. Leite, C. Fonseca, & L. P. Costa. 2011. Small mammals in the diet of barn owls, Tyto alba (Aves: Strigiformes) along the mid- Araguaia River in central Brazil. Zoologia 28:709-716. [ Links ]
61. Roda, S. A. 2006. Dieta de Tyto alba na Estação Ecológica do Tapacurá, Pernambuco, Brasil. Revista Brasileira de Ornitologia 14:449-452. [ Links ]
62. Scheibler, D. R., & A. U. Christoff. 2007. Habitat associations of small mammals in southern Brazil and use of regurgitated pellets of birds of prey for inventorying a local fauna. Brazilian Journal of Biology 67:619-25. [ Links ]
63. Silva, C. R., A. R. Percequilo, G. E. Iack Ximenes, & M. De Vivo. 2003. New distributional records of Blarinomys breviceps (Winge, 1888) (Sigmodontinae, Rodentia). Mammalia 67:147-152. [ Links ]
64. Souza, D. P., P. H. Asfora, T. C. Lira, & D. Astúa. 2010. Small mammals in barn owl (Tyto alba-Aves, Strigiformes) pellets from Northeastern Brazil, with new records of Gracilinanus and Cryptonanus (Didelphimorphia, Didelphidae). Mammalian Biology 75:370-374. [ Links ]
65. Taylor, I. 1994. Barn Owls: predator-prey relationship and conservation. Cambridge University Press, Cambridge. [ Links ]
66. Terry, R. C. 2010a. On raptors and rodents: testing the ecological fidelity and spatiotemporal resolution of cave death assemblages. Paleobiology 36:137-160. [ Links ]
67. Terry, R. C. 2010b. The dead do not lie: using skeletal remains for rapid assessment of historical small-mammal community baselines. Proceedings of the Royal Society of London 277:1193-1201. [ Links ]
68. Terry, R. C., C. L Li, & E. A. Hadly. 2011. Predicting small‐mammal responses to climatic warming: autecology, geographic range, and the Holocene fossil record. Global Change Biology 17:3019-3034. [ Links ]
69. Travassos, L. E. 2010. Considerações sobre o carste da região de Cordisburgo, Minas Gerais, Brasil. Tradição Plana, Belo Horizonte. [ Links ]
70. Trolle, M., J. R. Bissaro, & H. M. Prado. 2007. Mammal survey at a ranch of the Brazilian Cerrado. Biodiversity and Conservation 4:1205-1211. [ Links ]
71. Voss, R. S. 1993. A revision of the Brazilian muroid rodent genus Delomys: with remarks on "thomasomyine" characters. American Museum novitates 3073:1-44. [ Links ]
72. Voss, R. S., & P. Myers. 1991. Pseudoryzomys simplex (Rodentia: Muridae) and the significance of Lund´s Collections from the Caves of Lagoa Santa, Brazil. Bulletin of the American Museum of Natural History 206:414-432. [ Links ]
73. Wilson, D. E., & D. M. Reeder (Eds.). 2005. Mammal species of the world: a taxonomic and geographic reference. Johns Hopkins University Press, Baltimore. [ Links ]
74. Winge, H. 1887. Jordfundne og nulevende Gnavere (Rodentia) fra Lagoa Santa, Minas Geraes, Brasilien. E Museo Lundii 1:1-200. [ Links ]
Voucher numbers
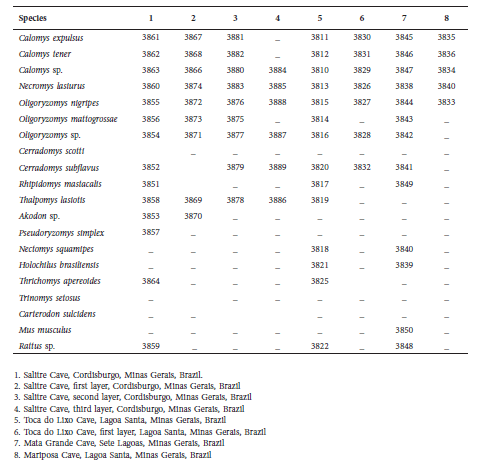
Table 1 Voucher numbers of specimens used in this study deposited in João Moojen Museum of Zoology (MZUFV).
Supplement 1
https://www.sarem.org.ar/wp-content/uploads/2018/06/SAREM_MastNeotrop_25-1_Boroni-sup1.docx
Fig. S1. Sumidouro State Park: A) Lagoon Sumidouro in the dry season; B) Vegetation; C) Limestone outcrop. Peter Lund Natural Monument: D) Salitre Cave, collection point indicated by the arrow.
Fig. S2. Mandibles (md) and skull in ventral view (vv) and dorsal (dv) of rodents in owl pellets in central karst region of Minas Gerais. (A) Calomys expulsus (dv, vv, md); (B) C. tener (dv, vv, md); (C) Necromys lasiurus (dv, vv, md); (D) Akodon sp. (md); (E) Thalpomys lasiotis (md); (F) Oligoryzomys nigripes (dv, vv, md); (G) O. mattogrossae (dv, vv, md); (H) Cerradomys subflavus (dv, vv, md); (I) C. scotti (md).
Fig. S3. Mandibles (md) and skull in ventral view (vv) and dorsal (dv) of rodents in owl pellets in central karst region of Minas Gerais. (A) Pseudoryzomys simplex (vv); (B) Holochilus brasiliensis (vv, md); (C) Nectomys squamipes (vv, md); (D) Rhipidomys matacalis (dv, vv, md); (E) Thrichomys apereoides (dv, vv, md); (F) Trinomys setosus (md); (G) Carterodon sulcidens (md).
Fig. S4. Rarefaction analysis graph of all Superficial samples. Blue = Salitre Cave; Green = Toca do Lixo; Yellow = Mata Grande Cave; Red = Mariposas Cave. Y axis = Estimated richness. X axis = sample number (MNI). CI = 95%.
Fig. S5. Rarefaction analysis graph of the Salitre Cave samples, Peter Lund Natural Monument, Cordisburgo. Red = Superficial; Blue = Stratified. Y axis = Estimated richness. X axis = sample number (MNI). CI = 95%.
Fig. S6. Rarefaction analysis graph of the Toca do Lixo samples, Sumidouro State Park, Pedro Leopoldo / Lagoa Santa. Blue = Superficial; Red = Stratified. Y axis = Estimated richness. X axis = sample number (MNI). CI = 95%.
Table S1. Rodents collected in owl pellets in the Cerrado and Atlantic Forest biomes. (A) Motta- Junior & Talamoni (1996); (B) Motta-Junior & Alho (2000); (C) Bonvicino & Bezerra (2003); (D) Bueno & Motta-Junior (2008); (E) Magrini & Facure (2008); (F) Rocha et al. (2011); (G) Roda (2006); (H) Souza et al. (2010); (I) Lemos et al. 2015; (J) Scheibler & Christoff (2007); (K) Escarlate-Tavares & Pessoa (2005); (L) Present study.

