










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkMastozoología neotropical
Print version ISSN 0327-9383On-line version ISSN 1666-0536
Mastozool. neotrop. vol.25 no.2 Mendoza Dec. 2018
ARTÍCULO
Diversity and seasonality of a phyllostomid assemblage from the Atlantic Forest of southeastern Brazil
Breno Mellado, Lucas de Oliveira Carneiro, Marcelo R. Nogueira and Leandro R. Monteiro
Laboratório de Ciências Ambientais, CBB, Universidade Estadual do Norte Fluminense, Campos dos Goytacazes, RJ, Brasil [Correspondencia: Breno Mellado <breno_mellado_rocha@hotmail.com>].
Abstract
Phyllostomids are locally abundant, ecologically diverse, and form rich assemblages. They also provide essential ecosystem services, which may change along the year in response to changes in bat abundances. Here we report on a phyllostomid assemblage studied at Reserva Biológica União (RBU), an Atlantic Forest area with outstanding diversity in southeastern Brazil. We assessed seasonal variation in diversity profiles for alpha and beta components, as well as in the abundance of the most common species. Spatial variation in phyllostomid diversity profiles and components was also assessed, using published information from three nearby sites. Based on a sample of 786 mist net captures and records from monitored roosts, the phyllostomid assemblage at RBU was found to include at least 19 species, two of which recognized as endangered at the national level (Lonchorhina aurita and Lonchophylla peracchii). The diversity profile for RBU was heavily influenced by common species and was one of the richest but less diverse in the region due to its uneven distribution. The diversity profiles suggested no seasonal differences in alpha diversity. Beta diversity profiles for spatial and seasonal analyses showed that samples behaved as subsets of a single assemblage, suggesting a relative homogeneity of resources in these dimensions. At the population level, however, some expected seasonal variation in abundance was confirmed (e.g., in Artibeus spp. and in Carollia perspicillata). The high relative frequency of Sturnira tildae at RBUdifferentiated this site, thus reinforcing its conservation importance.
Resumo
Diversidade e sazonalidade de uma assembleia de filostomídeos da Floresta Atlântica do sudeste do Brazil.
Morcegos filostomídeos são localmente abundantes, ecologicamente diversos, e formam ricas assembleias. Eles fornecem serviços ecossistêmicos essenciais, cuja disponibilidade pode mudar ao longo do ano por flutuações na sua abundância. Nos reportamos aqui a uma assembleia de filostomídeos estudada na Reserva Biológica União (RBU), uma área com grande diversidade no sudeste do Brasil. Apresentamos uma avaliação da variação sazonal de perfis de diversidade para os componentes alfa e beta, assim como da abundância das espécies mais comuns. Variação espacial na riqueza e diversidade de filostomídeos também foi investigada a partir de dados publicados de três sítios próximos. Baseado em uma amostra de 786 capturas em redes de neblina e registros a partir do monitoramento de abrigos, a assembleia de filostomídeos da RBU inclui pelo menos 19 espécies, duas das quais reconhecidas nacionalmente como ameaçadas de extinção (Lonchorhina aurita e Lonchophylla peracchii). O perfil de diversidade da RBU mostrou-se fortemente influenciado pelas espécies mais comuns, sendo um dos mais ricos da região mas pouco diverso em função da distribuição pouco equitativa. Os perfis de diversidade alfa indicaram a ausência de diferenças sazonais. Os perfis de diversidade beta para análises espaciais e sazonais mostraram que as assembleias locais se comportam como subconjuntos de uma única assembleia, sugerindo uma relativa homogeneidade de recursos nessas dimensões. Em nível populacional, entretanto, variações sazonais já esperadas foram confirmadas (e.g., Artibeus spp. e C. perspicillata). A frequência elevada de Sturnira tildae na RBU diferencia esse site dos demais aqui analisados, reforçando sua importância para a conservação.
Key words: Diversity partition; Diversity profiles; Reserva Biológica União; Rio de Janeiro; Sturnira tildae.
Palavras chave: Partição de diversidade; Perfis de diversidade; Reserva Biológica União; Rio de Janeiro; Sturnira tildae.
Recibido 12 abril 2017.
Aceptado 23 abril 2018.
Editor asociado: N. Giannini
INTRODUCTION
The Neotropical region harbors the most diverse mammalian fauna in the world although its area ranks 4th among major biogeographic regions (Ojeda 2013). At least 1550 species inhabit this region, corresponding to 30% of the extant mammal fauna (Solari et al. 2012). The proportion of endemic species is also remarkable: nearly 70%, considering only the land forms (Solari et al. 2012). An examination of the literature concerning mammalian conservation, however, revealed that this region, although presenting high levels of endemism and threats, has received relatively little attention (Amori & Gippoliti 2000). In order to improve both the quantity and quality of the conservation actions directed to Neotropical mammals, it is important to understand patterns and processes that may account for their extraordinary diversity. Spatial patterns, like those related to latitude and elevation gradients, have long been investigated (e.g., Voss & Emmons 1996; Maestri & Patterson 2016), and an increasing effort has been made to understand temporal variation, particularly when related to climatic changes (Davies et al. 2011).
Essential to the understanding of large scale macroecological patterns, is the availability of consistent local surveys (Bergallo et al. 2003). If we consider the Atlantic Forest, one of the world’s most threatened biomes, results from bat surveys conducted with mist nets have accumulated since the 1980’s, allowing important generalizations, such as the relatively high influence of temperature seasonality on species richness (see Stevens 2013). This conclusion was based on an analysis of phyllostomid bats, which are easy to catch with ordinary ground-level mist nets and are highly diverse, both taxonomically and ecologically (Simmons & Voss 1998).
Although the Atlantic Forest in the state of Rio de Janeiro is comparatively well-sampled for bats (Bergallo et al. 2003; Stevens 2013), few studies have been conducted in the north of the state, where Reserva Biológica União (RBU) and other important reserves are located (Jenkins et al. 2011). This area is of great zoogeographical importance, given that several bat species in the Emballonuridae and Phyllostomidae reach the southernmost limit on record of their distribution in northern Rio de Janeiro (Rhynchonycteris naso and Gardnerycteris crenulatum)or in theneighboring state ofEspírito Santo (Centronycteris maximiliani, Trinycteris nicefori, Rhinophylla pumilio, Dryadonycteris capixaba and Dermanura gnoma; Gardner 2008; Peracchi & Nogueira 2010). Here we present results of a phyllostomid survey at RBU, focusing on diversity, assemblage composition, and seasonal abundance patterns of these bats. Temporal heterogeneity of resources such as food may result in different assemblage compositions along the year (Tschapka 2004; Rex et al. 2008; Pereira et al. 2010). The occurrence of seasonal changes is a reasonable expectation at least for frugivorous phyllostomids (Mello 2009), because their food seems to be more abundant in the wet season, a pattern confirmed for our study region (Mello et al. 2004; Lapenta 2007). The opposite is probably true for hematophagous species (Zortéa & Alho 2008), whose food (vertebrate blood) availability is not expected to change seasonally (Freitas et al. 2006). We also performed diversity comparisons including three other sites from the same region (São João river basin), to gain insight into bat diversity at the regional spatial scale. The various phyllostomid assemblages at São João river basin were suggested to present marked differences (Mello & Schittini 2005), and a comparative approach incorporating diversity profiles and the partition into alpha and beta components (Jost 2007; Marcon et al. 2012) may shed light into possible differences of assemblage composition at the basin scale.
MATERIAL AND METHODS
Study site
Reserva Biológica União (RBU; 22º27¢30² S and 42º02¢15² W) is a 2547 ha conservation unit (recently increased to 7756 ha) composed of Atlantic Forest fragments located in the municipality of Rio das Ostras, state of Rio de Janeiro, southeastern Brazil (Fig. 1). Most of its area is covered by submontane ombrophilous forest (47%) and lowland ombrophilous forest (29%), with the rest of the Reserve covered by secondary shrubby vegetation (capoeira) and Eucaliptus plantations (ICMBIO 2008). Climate in the region is hot and humid, with an average annual rainfall varying between 1500 and 2000 mm (Primo & Völker 2003). The less rainy months, from May to August (average monthly rainfall 59 mm), are also those with lower temperatures and are here recognized as a dry season. The wet season, from September to April presents an average monthly rainfall of 177 mm. The average daily temperatures are high throughout the year, ranging between 22°C in the dry season to 26 °C in the wet season (ICMBIO 2008).

Fig. 1. Map showing the location of Reserva Biológica União (RBU) and other forest remnants in the São João river basin used for comparison: Morro de São João (MSJ), Reserva Biológica de Poço das Antas (RBPA), and RPPN Fazenda Bom Retiro (FBR). Thick black lines indicate Federal Reserve Contours (RBU and RBPA). Shapefile for forest remnant areas obtained from SOS Mata Atlântica (http://mapas.sosma.org.br/).
The three other sites close to RBU, and for which there is published information on phyllostomid captures used in our analyses (see below), are Morro de São João (MSJ; Esbérard et al. 2013), Reserva Biológica de Poço das Antas (RBPA; Baptista & Mello 2001), and Reserva Particular do Patrimônio Natural Fazenda Bom Retiro (FBR; Menezes-Jr et al. 2015a). These sites are separated from RBU at distances of, respectively, 8, 20, and 25 km (Fig. 1), and their sizes are, respectively, 2000 ha, 5052 ha, and 494 ha. FBR is the farthest site from RBU and presents the smallest area in our samples, but it is located north to the BR101 federal road, in a patch of fragments that is more connected (via small forest corridors) to this latter site than to RBPA and MSJ. The most isolated site is MSJ, which is surrounded by pastures (Fig. 1).
Bat sampling
Sampling points were chosen along trails and roads (possible routes of bats’ flights) in the reserve interior, trying to cover both lowland and submontane forest areas. Following the map of internal roads of the reserve management plan, the four sampling points we selected were Estrada do Buracão, Trilha Interpretativa do Pilão, Estrada do Lava Pé and Estrada das Três Pontes (ICMBIO 2008). Bat sampling was conducted every two months between August 2012 and September 2015, with at least three capture sessions in each visit to the reserve. Sporadic net sessions were made from 2006 to 2011 during field courses promoted by the Graduate Program in Ecology and Natural Resources, Universidade Estadual do Norte Fluminense. These samples, and data from roosts, were not included in the statistical analyses, but they were used to complement our species list.
In each capture session we used 3 to 6 ground-level mist nets of 6, 9 or 12 m long and 3 m high. In addition to exploring trails and roads, we also placed these nets over water bodies and close to roosts. In general, nets were set before sunset and remained open for at least 5 hours. Nets were monitored every 20 minutes, to preserve physical integrity of individuals and prevent net damage. We restricted the sampling to the first half of the night, when phyllostomid bats are expected to be more active (Fleming et al. 1972; Fleming 1988; Aguiar & Marinho-Filho 2004). We avoided repeating sampling points in consecutive days and sampling in full-moon periods; reportedly, both situations reduce capture success (Bergallo et al. 2003; Esbérard 2006; Saldaña Vázquez & Munguía-Rosas 2013). Net effort was calculated according to Straube & Bianconi (2002), with a unit effort corresponding to one square meter of net exposed for one hour (h.m²). Vouchers were prepared as skin and skull or as fluid-preserved specimens with skull removed; all specimens were deposited in the Coleção de Mamíferos da Universidade Estadual do Norte Fluminense and at Coleção Adriano Lúcio Peracchi (Laboratório de Mastozoologia, Universidade Federal Rural do Rio de Janeiro). Classification follows Baker et al. (2016) at subfamily level; below subfamily, nomenclature follows Nogueira et al. (2014).
Data analysis
Intersite comparisons - The phyllostomid assemblage from RBU was compared to those sampled by other authors in the same region using diversity profiles decomposed into alpha, beta and gamma components (Marcon et al. 2014). The general function for diversity as effective numbers of species (Jost 2007) is:
 ,
,
where ps is the probability in the given assemblage that a sampled individual belongs to species s, and the exponent q is the order of the diversity index. Known indices are related to specific orders, such as species richness (q=0), Shannon entropy (q=1) and Simpson (q=2). The function is actually undefined at q=1, but its limit is equal to the exponential of Shannon entropy (Jost 2006). Larger values of q are more sensitive to most common species, whereas smaller values of q are more sensitive to rare species (Jost 2007). Considering a number of sites forming a meta-assemblage in a region (Marcon et al. 2014), the gamma diversity qDg, based on all individuals sampled, irrespective of collection site, can be decomposed into the components of alpha and beta diversities, as qDg=qDa * qDb. The alpha diversity is a within-site component, calculated as a weighted average diversity over all sites, and the beta diversity is a between site component (Marcon et al. 2012). Confidence intervals for the profiles were estimated with 1000 replicates sampled from the original distribution of the species (Marcon & Hérault 2015). The effective number of species, or elements, has slightly different interpretations depending on the component. For alpha and gamma diversities, they are effective numbers of species (Jost 2006), the diversity equivalent to an assemblage with the same number of equally abundant species. In the case of beta diversity, the effective number of elements ranges from 1 (the meta-assemblage behaves as a single set) to the maximum number of assemblages being compared (if each one has a completely different species composition from the others).
Diversity profiles were calculated for each site’s alpha and for the partition into a combined alpha, beta and gamma (Marcon et al. 2014). Profiles plot estimated diversity against the order (q) of the estimate. Because the changing orders give different weights to rare or common species, the profile of an assemblage can be compared and ranked against other assemblages with a more complete assessment of species distribution and the influence of dominant species. All diversity estimates were corrected for bias due to undersampling (Marcon & Hérault 2015), using an estimate of sample coverage (C) calculated from the number of singletons (species captured only once) in sample I, denoted S1i, and sample size ni as Ci=1 – S1i/ni. The sample coverage multiplies the species frequencies in the calculation of entropies and diversities, as presented in the correction formula in Marcon et al. (2014). Because the number of specimens captured was different among areas, we weighted the calculation of combined (total) alpha diversity using the proportion of their sample contribution to the meta-assemblage sample (ni/N).
Intrasite (seasonal) comparisons – We based our seasonal analyses on dry vs. wet season comparisons within RBU only (the other sites did not have seasonal data). We used capture rates (captures per unit effort—see net effort calculation above), and the partition of diversity profiles (described above) into alpha (within season) and beta (between season) components. For the following analyses, each capture session was treated as a sample unit, and bias related to differences in net effort was avoided by using capture rates, instead of raw capture numbers. Capture rates for the whole assemblage in different seasons were analyzed via a graphical comparison using box plots, where notches in the boxes provide a 95% confidence interval for the median (McGill et al. 1978) The proportion of captures of the most common species in the wet season was compared to a random expectation (the proportion of effort allocated to the same period) using 95% confidence intervals. Finally, we used distance-based redundancy analysis to assess seasonal differences in assemblage composition (Legendre & Anderson 1999). This ordination technique based on distances was chosen because we found a high proportion of zeros (80%) in the abundance matrices for species per sample (Zuur et al. 2007). Also, because we detected an over-dispersed distribution of abundances, we chose to transform the species abundances into presence/absence data and use the Jaccard similarity index (more precisely, its distance counterpart: Distance=1 – Similarity) because it is asymmetric (only presences are considered evidence of similarity) and metric (Legendre & Legendre 2012).
Software - Statistical analyses and graphics were performed using the R environment (R Development Core Team 2015). We calculated the diversity profiles with the R package entropart (Marcon & Hérault 2015). The redundancy analysis was performed in R package vegan (Oksanen et al. 2015).
RESULTS
Diversity at RBU and intersite comparisons
With a total effort of 15069 h.m², we captured 786 individuals belonging to 15 genera and 18 species of Phyllostomidae, grouped in 5 ensembles (Table 1). Frugivores corresponded to 93% of all captures, mainly due to the wide dominance of Carollia perspicillata in the assemblage (73%). The second most common species (9%), Artibeus lituratus, was also a fruit-eating bat. Except for Trachops cirrhosus, which raised our total richness to 19, all species were captured in mist nets away from known roosts. Most species (60%) were recorded in the beginning of our samplings (within the first 100 captures), but Gardnerycteris crenulatum was recorded only after more than 700 individuals were captured. Chrotopterus auritus was sampled in our first capture session and never recorded again.
Table 1
List of phyllostomid bats from Reserva Biológica União, municipality of Rio das Ostras, state of Rio de Janeiro, southeastern Brazil, including number of individuals by season, relative abundances, and ensemble classification. Individuals obtained from roosts are treated separately and were not used in statistical analysis.

Diversity profiles comparing RBU with other forest remnants in the São João river basin indicated that the order of diversity (the exponent q) strongly influenced the estimates, with a pattern of decreasing diversity when the most dominant species were given larger weights (Fig. 2). The alpha diversity within sites was non-comparable because the profiles intersect (Fig. 2, upper right panel), showing that ranking of diversity depended on q. Considering q=0 (species abundance is disregarded, equivalent to richness), the diversity of RBU was the largest. On the other hand, as q approached 1 (diversity based on Shannon entropy), the hyperabundance of C. perspicillata greatly decreased RBU diversity and it fell to a second-to-last position in the site ranking. The profiles of MSJ and FBR were less steep than RBU and RBPA, indicating a more even distribution of species abundances. These results suggested that the influence of C. perspicillata in the estimated richness and diversity measurements were larger for RBU than for other sites in the same region. The beta diversity was higher for smaller values of q, suggesting that the most relevant species composition differences among assemblages were due to rare species (Fig. 2, lower left panel). The 95% confidence interval ranged from approximately 1.3 to 1.7 when q=0, showing uncertainty due to sampling of rare species. The most common species were basically the same in all assemblages and the profile of beta diversity approached 1 as q approached 2. The regional diversity (gamma) was, therefore, mostly a function of the combined alpha diversity for local assemblages, as shown by the profile similarities (Fig. 2, upper left and lower right panels).

Fig. 2. Diversity profiles of assemblages at different locations within the São João River Basin. The diversity components (in number of effective elements) are plotted against order of diversity (the exponent q); 95% confidence intervals are shown as grey areas. Profiles for different sites are identified by acronyms and line type. RBU: solid line; RBPA: dotted line; MSJ: dashed line; FBR: dot and dashed line.
Seasonal variation in abundance and diversity at RBU
Capture rates for the whole phyllostomid assemblage in RBU did not differ between seasons (Fig. 3), as the boxplot comparison indicated an overlap of median confidence intervals. When the most common species were considered separately, groundstory frugivores (C.perspicillata and S. tildae)were more frequent than expected in the dry seasonand some fig specialists sensu Dumont (2003; specifically, Artibeus lituratus and A. obscurus) were more frequent than expected in the wet season (Fig.4). The confidence intervals of wet season capture proportions of sanguivorous Desmodus rotundus and the fig specialist Platyrrhinus recifinus, included the expected proportion of 53%, indicating no seasonal changes in capture.
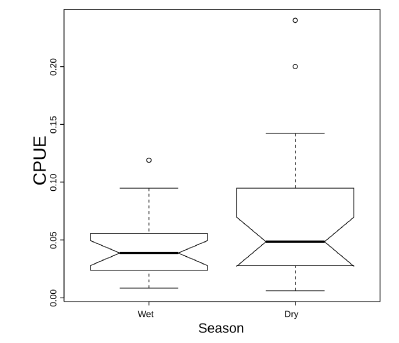
Fig. 3. Boxplot comparing bat captures per unit effort (individuals/h.m2) in sampling nights during different seasons at Reserva Biológica União. The notch indicates a 95% confidence interval for the median (thick horizontal line).

Fig. 4. Proportion of captures in the wet season with 95% confidence intervals for different species at Reserva Biológica União. Dotted line corresponds to proportion of capture effort in the wet season (0.53), considered as a null expectation of no difference in captures between seasons. Species acronyms are: Cp=Carollia perspicillata, Dr=Desmodus rotundus, Al=Artibeus lituratus, Ao=Artibeus obscurus, Pr=Platyrrhinus recifinus, St=Sturnira tildae.
When diversity profiles of different seasons were compared for RBU, the wet season presented a slightly higher diversity regardless of q, but the difference is contained in the confidence interval for the combined total alpha diversity (Fig. 5, upper right and left panels). Uncertainty regarding alpha and beta diversities was higher for lower values of q, probably due to the resampling of rare species, as can be seen in the larger confidence intervals (Fig. 5, upper and lower left panels). The multi-season diversity in RBU (gamma) had a large alpha component, as the between seasons (beta) profile showed small diversity, approaching 1 when more weight was given to common species (larger q). These results suggested that a single assemblage, instead of two (one for each season) seemed to be present in the area along the whole year.

Fig. 5: Diversity profiles of assemblages in different seasons at Reserva Biológica União. The diversity components (in number of effective elements) are plotted against order of diversity (the exponent q); 95% confidence intervals are shown as grey areas. Profiles for different seasons are identified by names and line type. Wet season: solid line; Dry season: dashed line.
The distance-based redundancy analysis showed significant (permuted P=0.0239) but weak (only c. 5% of similarity explained) seasonality in species composition between capture events. The association between species and seasons was due to higher relative abundances of C. perspicillata in the dry season and A.lituratus in the wet season, a tendency also detected in the capture rate data.
Discussion
Diversity at RBU and intersite comparisons
Phyllostomid assemblages have been extensively sampled along the Atlantic Forest, with richness values ranging between 4 and 30 species, and averaging 14 (N=60 sites; Stevens 2013). With at least 19 phyllostomid species, RBU is slightly above the average for the biome, with an assemblage similar to those sampled in the three other localities in the same region. These areas can be viewed as subsamples of a single general assemblage anteriorly spread over the São João river basin (Grativol et al. 2008). Given the heterogeneity of the regional landscape (e.g., MSJ is more isolated from other forests and RBPA is more fragmented), this similarity in species composition deserves further consideration. Although a pattern of species abundance distribution with a few very abundant species and many rare species characterizes all areas (McGill et al. 2007), the abundance distributions do not follow similar models and can be said to be non-comparable (Tóthmérész 1995). This happens because it is possible to find diversity indices that rank the assemblages in different ways (depending on q). The diversity profiles for RBU and RBPQ indicated a strong sensitivity to dominant species, whereas for MSJ and FBR, the abundance distributions were more equitable and the diversity profiles were less influenced by the most common species. This pattern was observed because a strong dominance of C. perspicillata is shared only between RBU (73% of all phyllostomids) and RBPA (78%). The lower abundance of C. perspicillata at MSJ (37%) and FBR (46%) may be related to local plant assemblages, more specifically to a reduced availability of Piper fruits (Esbérard et al. 2013), which form the core of the diet of this species (Fleming 1986). This association can be viewed as a more general pattern for frugivorous phyllostomids (Rex et al. 2008).
The high abundance of C. perspicillata is determinant for the strong dominance of the frugivore ensemble at RBU, but this pattern seems to be widespread in Neotropical bat assemblages, and not only when just phyllostomids are considered (Lim & Engstrom 2001). Several authors have emphasized possible methodological biases leading to this pattern (Simmons & Voss 1998), but as predicted by energy availability (Robinson & Redford 1986), phytophagous (frugivorous and nectarivorous) bats are expected to be more abundant at a particular site than those in higher trophic levels, such as insectivorous and carnivorous (Arita 1993). At our study site, phyllostomid nectarivores did not constitute an abundant ensemble (just 1.9% of all captures in this family), and the same was observed in RBPA and in FBR (0.7%). A relatively higher abundance of nectarivores was recorded at MSJ (4.7%); in this respect, this site, departs from the regional tendency, but this may be an artifact of sampling bats close to banana plantations (Esbérard et al. 2013).
The diversity profile for MSJ indicated a more even distribution of abundances and higher diversity when common species were given greater weight. This was a surprising result, considering that some detrimental aspects to bat diversity, such as isolation from other forested areas and insertion in a harsh matrix (Meyer et al. 2016), are particularly noticeable at this site. It can be argued, however, that MSJ presents a more favorable shape to maintain bat diversity, because it is more compact and rounded than the other sites, reducing potential negative effects of habitat edges (Meyer et al. 2016).This argument is also congruent with our finding that RBPA, which is composed of a more fragmented landscape (Brito et al. 2004; Figueiredo & Fernandez 2004), presented the lowest phyllostomid diversity.
The presences at RBU of Lonchorhina aurita and Lonchophylla peracchii, species categorized as endangered at the national level (MMA 2014), are noteworthy and reinforcethe importance of this natural reserve for conservation. Maybe even more remarkable is the unusual abundance pattern we recorded for Sturnira lilium and S. tildae. These species are widely distributed in eastern Brazil (Velazco & Patterson 2013), but the former is usually much more common in local inventories than the latter (Esbérard 2006; Luz et al. 2013). The reverse trend recorded at RBU—S. tildae four times more abundant than S. lilium—should be investigated under a macroecological approach, but it might be the case that factors such as dependence of lowland humid forests with low fragmentation levels contributed to this pattern (Brosset & Charles-Dominique 1990; Simmons & Voss 1998). Support to this hypothesis comes also from the absence of this species at RBPA, a more fragmented area (Baptista & Mello 2001) and from the more isolated MSJ (Esbérard et al. 2013). In fact, S. tildae is currently recorded in only a few localities in the state of Rio de Janeiro, and in all of them few individuals were captured (Esbérard et al. 2006; Luz et al. 2011, 2013; Menezes-Jr et al. 2015b; Souza et al. 2015). In states like Rio de Janeiro, where most lowland forests were removed and conservation units of integral protection are largely circumscribed to mountain areas (Tanizaki-Fonseca & Moulton 2000), S. tildae populations may be facing a substantial decline.
From a zoogeographic standpoint, a remarkable species in our phyllostomid list was Gardnerycteris crenulatum. This bat ranges from Mexico to southeastern Brazil (Williams & Genoways 2008), and its southernmost known records came from localities near our study site (Mello & Pol 2006). Capture data obtained at RBU are in agreement with those summarized by Mello & Pol (2006), including the association of G. crenulatum with water bodies (small dam) and its occurrence in low abundances. In Mexico, this species presents a restricted distribution and is rare, leading Ceballos (2014) to classify it as endangered. In Rio de Janeiro, G. crenulatum is classified as vulnerable, due to the small size of its populations and destruction of its habitat (Bergallo et al. 2000). Our record, nearly 17 years after the captures reported by Mello & Pol (2006), add a third locality for this species in Rio de Janeiro and reinforces the importance of forests at São João river basin to protect its apparently meridional edge populations in eastern Brazil—this species has never been captured in more southern and intensively sampled regions of Rio de Janeiro and São Paulo states (e.g., Esberard 2003; Bolzan et al. 2010; Garbino 2016).
Seasonal variation in abundance patterns
The phyllostomid assemblage did not present significant changes in composition throughout the year at RBU, but behind the similar general capture rates at both seasons, consistent evidences of seasonal shifts in a few species were detected, corroborating previous studies in the Atlantic Forest (Mello 2009). The most conspicuous pattern was the higher capture rate of C. perspicillata and S. tildae in drier months, contrasting with the trend of Artibeus spp. to be more frequent in the wet season. Fruit availability seems to be a main driver of general abundance patterns in frugivorous species (Rex et al. 2008), and seasonal changes are also likely to respond to this factor (Fleming 1988; Giannini 1999; Mello 2009). In the case of Artibeus spp., there is local evidence that some seasonality exists in their core fruit plants (sensu Fleming 1986). Lapenta (2007), for example, found that more species of Cecropia bear ripe fruits during the wet season in RBU.
The higher capture rates of groundstory bats in the drier season is contrary to our expectation, mainly in the case of C. perspicillata, whose core food plants (Piper spp.; Fleming 1986) produce fruit more abundantly in the wet season (Mello et al. 2004). As a possible explanation, one could argue that this bat needs to travel less between roosts and feeding areas during this season, which could reduce its chance of being captured in mist nets spread over the study area. In Costa Rica, C. perspicillata travels greater distances in the dry season, when resources are less abundant and more heterogeneously distributed (Fleming 1988). Also important to support this hypothesis is the high site fidelity previously recorded for C.perspicillata (Bianconi et al. 2006), since species that forage over large areas, like Artibeusspp. (Morrison 1978), probably go beyond our sampling stations or move to different forest fragments during the dry season. For S. tildae, whose core food plants are species of Solanum (Lobova et al. 2009), variation in capture rates are difficult to interpret given the lack of local phenological data.
The lack of seasonality in capture rates of D. rotundus was in accordance with our expectation and literature records (Young 1971; Freitas et al. 2006; Zortéa & Alho 2008), and is probably explained by the continuous availability of mammals as blood source for this species at RBU and its immediate surroundings. Not only cattle is available in the area; the reserve also harbors abundant populations of collared peccary (Tayassu pecari) and capybaras (Hydrochoerus hydrochaeris; L. R. Monteiro, unpublished data), both of which can be considered potential targets for this bat (Bobrowiec et al. 2015). The other aseasonal species in our sample, Platyrrhinus recifinus, is a canopy fruit-eating bat, which departed from results obtained for other Moraceae specialists, Artibeus spp. Ficus and Cecropia are used frequently by these bats (Lobova et al. 2009), but P. recifinus is a much smaller species, which may be a limiting factor for moving to other foraging areas in periods of food scarcity (Kalko et al. 1996; but see also Albrecht et al. 2007 and Villalobos-Chaves et al. 2017).
The seasonal diversity profiles at RBU were largely affected by the most common species, but the assemblages in the two seasons presented a similar pattern of dependence of diversity on q. These results can be viewed as evidence of the potential of the dry season at RBU to support a rich phyllostomid assemblage. This is in agreement with results from other assemblage-level analyses performed here, such as the redundancy analysis, which showed small effects or no difference related to season. Investigating phyllostomids in an Atlantic Forest site in Rio de Janeiro, Gomes et al. (2015) found no seasonal differences in species richness or assemblage composition, but higher general abundance in the wet season. Although these authors did not analyze species separately, it can be derived from their data that this abundance pattern was largely determined by the same species that are dominant at RBU (A. lituratus and C. perspicillata). While the ecological role of these species, mainly seed dispersal, might be more intense depending on the season (Mello 2009), the year-round maintenance of similar assemblages, including their ensemble composition, might also prove important for ecosystem functioning and regeneration (Silveira et al. 2011).
Concluding remarks
Analyses of local assemblages are essential tools for the description of world’s biodiversity, establishing the basic knowledge for its conservation (Balmford & Gaston 1999). Here we highlighted the importance of RBU for the conservation of Phyllostomidae, the most ecologically diverse bat family, analyzing our data set both qualitatively and quantitatively, and including both spatial (inter-site) and temporal (seasonal) components of variation. Although at a regional scale, results presented here pointed to low among-site diversity, singularities, as revealed by species such as S.tildae, show that RBU may conceal ecological opportunities that are masked when assemblage metrics are considered alone. The metric richness, and alpha and beta diversity, are also insensitive to differences in functional diversity, and further analyses in the same region should formally incorporate this aspect. Our qualitative data in this respect showed that nearly all the same five ensembles (see Table 1) are widely distributed, being gleaning animalivorous the only absence from FBR. This similarity is surprising, considering the level of fragmentation in the landscape, and suggest some resilience of this bat family, which is important for the maintenance of its ecological function.
ACKNOWLEDGEMENTS
The authors would like to thank W. Costa-Jr (RBU administrator), K. V. Maia and M. M. Morais-Jr for support and providing infrastructure for the field work. We also thank C. Esbérard and M. T. Nascimento for providing essential information about the forest remnants. C. R. Ruiz-Miranda, M. T. Nascimento, N. Giannini and an anonymous reviewer provided useful insights in previous versions of this manuscript. This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Rio de Janeiro (FAPERJ), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
LITERATURE CITED
1. Aguiar, L. M. S., & J. Marinho-Filho. 2004. Activity patterns of nine phyllostomid bat species in a fragment of the Atlantic Forest in southeastern Brazil. Revista Brasileira de Zoologia 21:385-390. [ Links ]
2. Albrecht, L., C. F. Meyer, & E. K. Kalko. 2007. Differential mobility in two small phyllostomid bats, Artibeus watsoni and Micronycteris microtis, in a fragmented neotropical landscape. Acta Theriologica 52:141-149. [ Links ]
3. Amori, G., & S. Gippoliti. 2000. What do mammalogists want to save? Ten years of mammalian conservation biology. Biodiversity & Conservation 9:785-793. [ Links ]
4. Arita, H. T. 1993. Rarity in neotropical bats: correlations with phylogeny, diet, and body mass. Ecological Applications 3:506-517. [ Links ]
5. Baker, R. J., S. Solari, A. Cirranello, & N. B. Simmons. 2016. Higher level classification of phyllostomid bats with a summary of DNA synapomorphies. Acta Chiropterologica 18:1-38. [ Links ]
6. Balmford, A., & K. J. Gaston. 1999. Why biodiversity surveys are good value. Nature 398:204-205. [ Links ]
7. Baptista, M., & M. A. R. Mello. 2001. Preliminary inventory of the bat species of the Poço das Antas Biological Reserve, RJ. Chiroptera Neotropical 7:133-135. [ Links ]
8. Bergallo, H. G. et al. 2000. Mamíferos. A fauna ameaçada de extinção do Estado do Rio de Janeiro (H. G. Bergallo, C. F. D. Rocha, M. A. S. Alves & M. Van Sluys, eds.). Editora da Universidade do Estado do Rio de Janeiro, Rio de Janeiro. [ Links ]
9. Bergallo, H. G. et al. 2003. Bat species richness in Atlantic Forest: What is the minimum sampling effort? Biotropica 35:278-288. [ Links ]
10. Bianconi, G. V., S. B. Mikich, & W. A. Pedro. 2006. Movements of bats (Mammalia, Chiroptera) in Atlantic Forest remnants in southern Brazil. Revista Brasileira de Zoologia 23:1199-1206. [ Links ]
11. Bobrowiec, P. E. D., M. R. Lemes, & R. Gribel. 2015. Prey preference of the common vampire bat (Desmodus rotundus, Chiroptera) using molecular analysis. Journal of Mammalogy 96:54-63. [ Links ]
12. Bolzan, D. P. et al. 2010. Morcegos da região da Costa Verde e adjacências, litoral sul do estado do Rio de Janeiro. Chiroptera Neotropical 16:585-595. [ Links ]
13. Brito, D., L. C. Oliveira, & M. A. R. Mello. 2004. An overview of mammalian conservation at Poço das Antas Biological Reserve, southeastern Brazil. Journal for Nature Conservation 12:219-228. [ Links ]
14. Brosset, A., & P. Charles-Dominique. 1990. The bats from French Guiana: a taxonomic, faunistic and ecological approach. Mammalia 54:509-560. [ Links ]
15. Ceballos, G. 2014. Mammals of Mexico. Johns Hopkins University Press, Baltimore. [ Links ]
16. Davies, T. J., L. B. Buckley, R. Grenyer, & J. L. Gittleman. 2011. The influence of past and present climate on the biogeography of modern mammal diversity. Philosophical Transactions of the Royal Society B: Biological Sciences 366:2526-2535. [ Links ]
17. Dumont, E. R. 2003. Bats and fruit: an ecomorphological approach. Bat ecology (T. H. Kunz & B. Fenton, eds.). The University of Chicago Press, Chicago. [ Links ]
18. Esbérard, C. E. L. 2003. Diversidade de morcegos em área de Mata Atlântica regenerada no sudeste do Brasil. Revista Brasileira de Zoociências 5:189-211. [ Links ]
19. Esbérard, C. E. L. 2006. Efeito da coleta de morcegos por noites seguidas no mesmo local. Revista Brasileira de Zoologia 23:1093-1096. [ Links ]
20. Esbérard, C. E. L. et al. 2006. Morcegos da Ilha Grande, Angra dos Reis, RJ, Sudeste do Brasil. Revista Brasileira de Zoociências 8:147-153. [ Links ]
21. Esbérard, C. E. L., L. de Moraes-Costa, & J. L. Luz. 2013. Morcegos de Morro de São João, estado do Rio de Janeiro, sudeste do Brasil. Bioscience Journal 29:449-457. [ Links ]
22. Figueiredo, M. S. L., & F. A. S. Fernandez. 2004. Contrasting effects of fire on populations of two small rodent species in fragments of Atlantic Forest in Brazil. Journal of Tropical Ecology 20:225-228. [ Links ]
23. Fleming, T. H. 1986. Opportunism versus specialization: the evolution of feeding strategies in frugivorous bats. Frugivores and seed dispersal (A. Estrada & T. H. Fleming, eds.). Dr W. Junk Publishers, Dordrecht. [ Links ]
24. Fleming, T. H. 1988. The short-tailed fruit bat: a study in plant-animal interactions. The University of Chicago Press, Chicago. [ Links ]
25. Fleming, T. H., E. T. Hooper, & D. E. Wilson. 1972. Three Central American bat communities: structure, reproductive cycles, and movement patterns. Ecology 53:556-569. [ Links ]
26. Freitas, M. B., A. F. Welker, & E. C. Pinheiro. 2006. Seasonal variation and food deprivation in common vampire bats (Chiroptera: Phyllostomidae). Brazilian Journal of Biology 66:1051-1055. [ Links ]
27. Garbino, G. S. T. 2016. Research on bats (Chiroptera) from the state of São Paulo, southeastern Brazil: annotated species list and bibliographic review. Arquivos de Zoologia 47:43-128. [ Links ]
28. Gardner, A. L. 2008. Mammals of South America, Volume 1. The University of Chicago Press, Chicago. [ Links ]
29. Giannini, N. P. 1999. Selection of diet and elevation by sympatric species of Sturnira in an Andean rainforest. Journal of Mammalogy 80:1186-1195. [ Links ]
30. Gomes, L. A. C., A. D. S. Pires, M. A. Martins, E. C. Lourenco, & A. L. Peracchi. 2015. Species composition and seasonal variation in abundance of Phyllostomidae bats (Chiroptera) in an Atlantic Forest remnant, southeastern Brazil. Mammalia 79:61-68. [ Links ]
31. Grativol, A. D., C. R. Ruiz-Miranda, A. M. De Godoy-Teixeira, & M. A. R. Schmidt. 2008. Abordagem de metapopulação para a conservação dos micos-leões-dourados na paisagem fragmentada da bacia do rio São João. Conservação do mico-leão-dourado: enfrentando os desafios de uma paisagem fragmentada (A. D. Grativol, C. R. Ruiz-Miranda, A. M. G. Teixeira & M. A. R. Schimidt, eds.). Universidade Estadual do Norte Fluminense, Campos dos Goytacazes, RJ. [ Links ]
32. ICMBIO. 2008. Plano de manejo da Reserva Biológica União, Encarte 3: Análise da Unidade de Conservação. Ministério do Meio Ambiente, Brasil. [ Links ]
33. Jenkins, C. N., S. L. Pimm, & M. D. S. Alves. 2011. How conservation GIS leads to Rio de Janeiro, Brazil. Natureza & Conservação 9:152-159. [ Links ]
34. Jost, L. 2006. Entropy and diversity. Oikos 113:363-375. [ Links ]
35. Jost, L. 2007. Partitioning diversity into independent alpha and beta components. Ecology 88:2427-2439. [ Links ]
36. Kalko, E. K., C. O. Handley, & D. Handley. 1996. Organization, diversity, and long-term dynamics of a Neotropical bat community. Long-Term Studies of Vertebrate Communities (M. L. Cody & J. A. Smallwood, eds.). Academic Press, San Diego. [ Links ]
37. Lapenta, M. J. 2007. Frugivoria, dispersão primária e secundária de sementes consumidas por micos-leões-dourados (Leontopithecus rosalia) na Reserva Biológica União, RJ. PhD Thesis. Universidade de São Paulo, São Paulo. [ Links ]
38. Legendre, P., & M. J. Anderson. 1999. Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecological Monographs 69:1-24. [ Links ]
39. Legendre, P., & L. Legendre. 2012. Numerical ecology. Elsevier, Amsterdam. [ Links ]
40. Lim, B. K., & M. D. Engstrom. 2001. Bat community structure at Iwokrama forest, Guyana. Journal of Tropical Ecology 17:647-665. [ Links ]
41. Lobova, T. A., C. K. Geiselman, & S. A. Mori. 2009. Seed dispersal by bats in the Neotropics. New York Botanical Garden, New York. [ Links ]
42. Luz, J. L., L. D. M. Costa, E. C. Lourenço, & C. E. L. Esbérard. 2011. Bats (Mammalia, Chiroptera) from Reserva Rio das Pedras, Rio de Janeiro, Southeastern Brazil. Biota Neotropica 11:95-101. [ Links ]
43. Luz, J. L., L. M. Costa, T. Jordão-Nogueira, C. E. L. Esbérard, & H. G. Bergallo. 2013. Morcegos em área de Floresta Montana, Visconde de Mauá, Resende, Rio de Janeiro. Biota Neotropica 13:190-195. [ Links ]
44. Maestri, R., & B. Patterson. 2016. Patterns of species richness and turnover for the South American rodent fauna. PLoS One 11:e0151895. [ Links ]
45. Marcon, E., & B. Hérault. 2015. entropart: An R package to measure and partition diversity. Journal of Statistical Software 67. [ Links ]
46. Marcon, E., B. Hérault, C. Baraloto, & G. Lang. 2012. The decomposition of Shannon’s entropy and a confidence interval for beta diversity. Oikos 121:516-522.
47. Marcon, E., I. Scotti, B. Hérault, V. Rossi, & G. Lang. 2014. Generalization of the partitioning of Shannon diversity. Plos One 9:e90289. [ Links ]
48. Mcgill, B. J. et al. 2007. Species abundance distributions: moving beyond single prediction theories to integration within an ecological framework. Ecology Letters 10:995-1015. [ Links ]
49. Mcgill, R., J. W. Tukey, & W. A. Larsen. 1978. Variation of box plots. The American Statistician 32:12-16. [ Links ]
50. Mello, M. A. R. 2009. Temporal variation in the organization of a Neotropical assemblage of leaf-nosed bats (Chiroptera: Phyllostomidae). Acta Oecologica 35:280-286. [ Links ]
51. Mello, M. A. R., & A. Pol. 2006. First record of the bat Mimon crenulatum (É. Geoffroy, 1801) (Mammalia: Chiroptera) in the state of Rio de Janeiro, Southeastern Brazil. Brazilian Journal of Biology 66:6295-299. [ Links ]
52. Mello, M. A. R., & G. M. Schittini. 2005. Ecological analysis of three bat assemblages from conservation units in the lowland Atlantic Forest of Rio de Janeiro, Brazil. Chiroptera Neotropical 11:1-2. [ Links ]
53. Mello, M. A. R., G. M. Schittini, P. Selig, & H. G. Bergallo. 2004. A test of the effects of climate and fruiting of Piper species (Piperaceae) on reproductive patterns of the bat Carollia perspicillata (Phyllostomidae). Acta Chiropterologica 6:309-318. [ Links ]
54. Menezes-Jr, L. F. P., A. C. Duarte, M. D. Contildes, & A. L. Peracchi. 2015a. Comparação da quiropterofauna em área florestada e área aberta da RPPN Fazenda Bom Retiro, Rio de Janeiro, Brasil. Iheringia, Série Zoologia 105:271-275. [ Links ]
55. Menezes-Jr, L. F. P., A. C. Duarte, M. D. Contildes, & A. L. Peracchi. 2015b. Lista de Morcegos (Mammalia Chiroptera) do Parque Natural Municipal da Serra do Mendanha, Município do Rio de Janeiro, Rj, Brasil. Biodiversidade Carioca: segredos revelados (J. A. L. Pontes, ed.). Technical Books, Rio de Janeiro. [ Links ]
56. Meyer, C. F. J., M. J. Struebig, & M. R. Willig. 2016. Responses of tropical bats to habitat fragmentation, logging, and deforestation. Bats in the Anthropocene: conservation of bats in a changing world (C. Voigt & T. Kingston, eds.). Springer, New York. [ Links ]
57. MMA. 2014. Portaria nº 444 de 17 de dezembro de 2014. Diário Oficial da União 245:121-126. [ Links ]
58. Morrison, D. W. 1978. Lunar phobia in a neotropical fruit bat, Artibeus jamaicensis (Chiroptera: Phyllostomidae). Animal Behaviour 26:852-855. [ Links ]
59. Nogueira, M. R., I. P. De Lima, R. Moratelli, V. Da-Cunha Tavares, R. Gregorin, & A. L. Peracchi. 2014. Checklist of Brazilian bats, with comments on original records. Check List 10:808-821. [ Links ]
60. Ojeda, R. A. 2013. Diversity and conservation of Neotropical mammals. Encyclopedia of Biodiversity 2:582-594. [ Links ]
61. Oksanen, J. et al. 2015. Vegan: Community Ecology Package <https://CRAN.R-project.org/package=vegan> [ Links ].
62. Peracchi, A. L., & M. R. Nogueira. 2010. Lista anotada dos morcegos do Estado do Rio de Janeiro, sudeste do Brasil. Chiroptera Neotropical 16:508-519. [ Links ]
63. Pereira, M. J. R., J. T. Marques, & J. M. Palmeirim. 2010. Ecological responses of frugivorous bats to seasonal fluctuation in fruit availability in Amazonian forests. Biotropica 42:680-687. [ Links ]
64. Primo, P. B., & C. M. Völker. 2003. Bacias hidrográficas dos rios São João e das Ostras: águas, terras e conservação ambiental. Consórcio Intermunicipal Lagos São João, Rio de Janeiro. [ Links ]
65. R Development Core Team. 2015. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org/. [ Links ]
66. Rex, K., D. H. Kelm, K. Wiesner, T. H. Kunz, & C. C. Voigt. 2008. Species richness and structure of three Neotropical bat assemblages. Biological Journal of the Linnean Society 94:617-629. [ Links ]
67. Robinson, J. G., & K. H. Redford. 1986. Body size, diet, and population density of Neotropical forest mammals. American Naturalist 128:665-680. [ Links ]
68. Saldaña-Vázquez, R. A., & M. A. Munguía-Rosas. 2013. Lunar phobia in bats and its ecological correlates: a meta-analysis. Mammalian Biology 78:216-219. [ Links ]
69. Silveira, M., L. Trevelin, M. Port-Carvalho, S. Godoi, E. N. Mandetta, & A. P. Cruz-Neto. 2011. Frugivory by phyllostomid bats (Mammalia: Chiroptera) in a restored area in Southeast Brazil. Acta Oecologica 37:31-36. [ Links ]
70. Simmons, N. B., & R. S. Voss. 1998. Mammals of Paracou, French Guiana, a Neotropical Lowland Rainforest fauna. Bulletin of the American Museum of Natural History 237:1-219. [ Links ]
71. Solari, S., P. Velazco, & B. Patterson. 2012. Hierarchical organization of Neotropical mammal diversity and its historical basis. Bones, clones, and biomes: The history and geography of recent Neotropical mammals (B. D. Patterson & L. P. Costa, eds.). University of Chicago Press, Chicago. [ Links ]
72. Souza, R. F. et al. 2015. Bats (Mammalia, Chiroptera) in a remnant of Atlantic Forest, Rio de Janeiro, Southeastern Brazil. Neotropical Biology and Conservation 10:9-14. [ Links ]
73. Stevens, R. D. 2013. Gradients of bat diversity in Atlantic Forest of South America: environmental seasonality, sampling effort and spatial autocorrelation. Biotropica 45:764-770. [ Links ]
74. Straube, F. C., & G. V. Bianconi. 2002. Sobre a grandeza e a unidade utilizada para estimar esforço de captura com utilização de redes-de-neblina. Chiroptera Neotropical 8:150-152. [ Links ]
75. Tanizaki-Fonseca, K., & T. P. Moulton. 2000. A fragmentação da Mata Atlântica no Estado do Rio de Janeiro e a perda de biodiversidade. A fauna ameaçada de extinção do Estado do Rio de Janeiro (H. G. Bergallo, C. F. D. Rocha, M. A. S. Alves & M. V. Sluys, eds.). Editora da Universidade do Estado do Rio de Janeiro, Rio de Janeiro, RJ. [ Links ]
76. Tóthmérész, B. 1995. Comparison of different methods for diversity ordering. Journal of Vegetation Science 6:283-290. [ Links ]
77. Tschapka, M. 2004. Energy density patterns of nectar resources permit coexistence within a guild of Neotropical flower-visiting bats. Journal of Zoology 236:7-21. [ Links ]
78. Velazco, P. M., & B. D. Patterson. 2013. Diversification of the yellow-shouldered bats, genus Sturnira (Chiroptera, Phyllostomidae), in the New World tropics. Molecular Phylogenetics and Evolution 68:683-698. [ Links ]
79. Villalobos-Chaves, D., M. Spínola-Parallada, K.Heer, E. K. Kalko, & B. Rodríguez-Herrera. 2017. Implications of a specialized diet for the foraging behavior of the Honduran white bat, Ectophylla alba (Chiroptera: Phyllostomidae). Journal of Mammalogy 98:1193-1201. [ Links ]
80. Voss, R. S., & L. H. Emmons. 1996. Mammalian diversity in Neotropical lowland rainforests: a preliminary assessment. Bulletin of the American Museum of Natural History 230:1-115. [ Links ]
81. Williams, S. L., & H. H. Genoways. 2008. Subfamily Phyllostominae Gray 1825. Mammals of South America, Volume 1: marsupials, xenarthrans, shrews, and bats (A. Gardner, ed.). The University of Chicago Press, Chicago. [ Links ]
82. Young, A. M. 1971. Foraging of vampire bats (Desmodus rotundus) in Atlantic wet lowland Costa Rica. Revista de Biologia Tropical 18:73-88. [ Links ]
83. Zortéa, M., & C. J. Alho. 2008. Bat diversity of a Cerrado habitat in central Brazil. Biodiversity and Conservation 17:791. [ Links ]
84. Zuur, A., E. N. Ieno, & G. M. Smith. 2007. Analysing ecological data. Springer, New York. [ Links ]

