










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBiocell
versão impressa ISSN 0327-9545
Biocell v.27 n.2 Mendoza abr./ago. 2003
Proteoglycans production by aortic vascular smooth muscle cells from hypertensive rats
Norma Risler, Claudia Castro, Montserrat Cruzado, Susana González and Roberto Miatello
Cell Culture Laboratory, Departments of Pathology and Morphophysiology, School of Medicine. National University of Cuyo (UNC), Mendoza, Argentina.
Address correspondence to: Dra. Norma Risler. Area de Fisiología Patológica, Departamento de Patología, Facultad de Ciencias Médicas, Universidad Nacional de Cuyo, Mendoza, ARGENTINA. E-mail: nrisler@fcm.uncu.edu.ar
Abstract: Remodeling of large and small arteries contributes to the development and complications of hypertension. Artery structural changes in chronic sustained hypertension include vascular smooth muscle cells (VSMC) proliferation and extracellular matrix (ECM) modifications. Extracellular constituents such as proteoglycans (PGs), may modulate vascular stiffness and VSMC growth and differentiation. We examined the effect of growth factors on secreted and membrane-bound PGs synthesis by cultured aortic smooth muscle cells (SMC) from 12- to 14- week-old spontaneously hypertensive rats (SHR) and age-matched Wistar rats. After stimulation with platelet-derived growth factor (PDGF-BB), 10% fetal calf serum (FCS) or 0.1% FCS as control, PGs synthesis (dpm/ng DNA) was evaluated in the medium (M-ECM) and in the cell layer (P-ECM) by a double-isotopic label method using both [3H]-glucosamine and [35S]-sodium sulfate which are incorporated into all complex carbohydrates or only into sulfated dysaccharides, respectively. Data are presented as percent of the control (0.1% FCS). SHR VSMC displayed a significantly greater synthesis of M-ECM [3H]-PGs than Wistar rat cells, with both treatments, but no differences in M-ECM [35S] uptake were found in any case. In the P-ECM, both PDGF-BB and 10% FCS produced a greater effect on [3H]-PGs and sulfated PGs synthesis in VSMC from SHR. An important change seen in SHR cells was a significant decreased sulfation, assesed by [35S]/[ 3H] ratio, in basal and stimulation conditions. Present results indicate the existence of changes in PGS synthesis and modulation in VSMC from a conduit-artery of SHR and support the pathophysiological role proposed for matrix proteoglycans in the vascular wall changes associated to hypertension and related vascular diseases as atherosclerosis.
Key words: Vascular Smooth Muscle Cells. Extracellular Matrix Proteoglycans. Hypertension. Spontaneously Hypertensive Rats.
Introduction
Structural changes in the arterial wall in chronic sustained hypertension include smooth muscle proliferation and extracellular matrix (ECM) modifications. ECM is the structural and functional support to which cells adhere and on which they grow, migrate and differentiate. The ECM of the normal artery wall is a collection of fibrous proteins and associated glycoproteins, embedded in a hydrated ground substance of glycosaminoglycans (GAGs) and proteoglycans (PGs). These distinct molecules are organized into a highly ordered network, that are closely associated with the vascular cells producing them. In addition to providing the architectural framework for the artery wall that impacts mechanical support and viscoelasticity, the ECM can regulate the behaviour of vascular cells, including their ability to migrate, proliferate and survive injury (Raines, 2000). Blood vessel wall ECM contains elastin, collagen and proteoglycans (Carey, 1991) and it behaves as a biologically active system, being an adequate balance of its components necessary for the normal vascular function (Tuñon et al., 2000). PGs are important nonfibrous matrix components of the arterial wall that carry unusual carbohydrates, the GAGs, which are composed by repeated disaccharide units and exist in different forms. Some are protein bound (the natural form); these contain sulfate associated to the carbohydrate. The other form is hyaluronic acid, which is made as a free GAG and lacks sulfate. GAGs, the sulfated ones in particular, have a strong negative charge. This charge makes it possible for GAGs to bind many substances, including growth factors. Further interesting features of this system are that the synthesis of some PGs by various kind of cells, such as vascular smooth muscle cells (VSMC) (Emoto et al., 1998) or glomerular mesangial cells (Dubey et al., 1997), is stimulated by growth factors; that many of the PGs function as modulators of growth factors (Ruoslahti and Yamaguchi, 1991) and that some PGs-growth factor interactions are mediated by the GAG components of the PGs (Yayon et al., 1991). Thus, ECM PGs play an important role in VSMC growth and differentiation (Hein et al., 1996).
An increase in tissue resistance, caused in part by an increase in aortic wall thickness, characterize chronic hypertension in both spontaneously hypertensive rats (SHR) and humans. Modification of aortic thickness is caused by hypertrophy of VSMC and a simultaneous modification in the amount of extracellular proteins. The relationship between VSMC and the matrix elements is thought to be a controlling factor in the elastic properties of the arterial wall. The alteration of this relationship is likely to play a role in the physiopathology of diseases characterized by vascular proliferative disorders, such as atherosclerosis, restenosis (McLeod et al., 1994) and hypertension. Several studies (Intengan et al.,1999) have demonstrated that disturbances in the matrix composition, in particular an increase in the collagen/elastin ratio, were responsible for the hypertrophy of the arterial wall that occurs during the rise in blood pressure. Remodeling of large and small arteries contributes to the development and complications of hypertension. Altered VSMC growth and increased deposition of ECM proteins are important processes in vascular remodeling in hypertension. ECM components, including PGs, represent an important regulatory mechanism that modulates the nature of VSMC growth response (Berk, 2001). Several changes in vascular ECM components have been described, such as increase in collagen and fibronectin, due to an augmented synthesis or a diminished degradation by matrix metaloproteinases activity (Intengan and Schiffrin, 2001). VSMC under dynamic mechanical stresses from arterial pressure, may modify their biomechanical enviroment in a manner that limits its potential biomechanical injury. ECM PGs are molecules that can play a prominent role in tissue mechanics. Mechanical deformation of cultured aortic SMC increases specific PGs synthesis and aggregation, indicating a highly coordinated ECM response to biomechanical stimulation (Lee et al., 2001) which may serve as an initial defense against mechanical stress, but it also could be detrimental because of PGs participation in the subendothelial retention of atherogenic lipoproteins.
Previously, we have reported that growth factors induce different profiles of PGs synthesis in VSMC obtained from resistance arteries (mesenteric vascular bed), of adult 12- to 14-week-old spontaneously hypertensive rats (SHR) with an established chronic blood pressure elevation or age-matched normotensive Wistar rats (Castro et al., 1999). We have also found that changes in sulfated PGs synthesis by VSMC of mesenteric arteries precede the vascular dysfunction associated to the development of hypertension (Risler et al., 2002).
In this study, we investigated the effect of growth factors on the synthesis of secreted and membrane-bound PGs by cultured vascular smooth muscle cells of a conduit artery, the thoracic aorta, from SHR and normotensive Wistar rats.
Material and Methods
Male SHR of 12-14- weeks- old and their age-matched Wistar normotensive control rats (n=8 in each group) were used for this study. SHR were acquired from the Veterinary School, University of La Plata, Argentina. They were housed under standardized conditions of controlled temperature (20ºC) and humidity (60%) and a 12-hour light/dark cycle. Animals were fed regular commercial pelleted rat chow and given tap water ad libitum. All procedures were performed in accordance with institutional guidelines for animal experimentation (Animal Experimentation Committee, School of Medicine, Universidad Nacional de Cuyo). Systolic blood pressure was monitored indirectly in conscious prewarmed slightly restrained rats by the tail-cuff method and recorded on a Grass model 7 polygraph (Grass Medical Instruments, MS, USA). The average of three pressure readings was recorded and systolic pressure was measured four times in each rat. SHR systolic pressure (183 ± 3 mmHg) was significantly greater than that of Wistar rats (114 ± 3 mmHg) (P<0.001), without significant difference in body weight.
Cell Culture
Unless otherwise noted, all reagents were obtained from Sigma Chemical Co (St Louis, MO, USA). The animals were killed by decapitation under ether anesthesia and arterial vessels were aseptically excised and placed in chilled Hank's Buffered Saline Solution (HBSS) and antibiotic mixture for further dissection. Aortic cells were isolated according to a technique previously described (Cruzado et al., 1998). Aortic SMC were obtained from two to three thoracic aortas by enzyme dispersion with 1.5 mg/mL collagenase (Class II Worthington Biochemical Corp, NJ, USA) in F-12 modified Eagle's medium (MEM) with 10% fetal calf serum (FCS) (Gen S.A., Buenos Aires, Argentina). After a 2 to 3 h period in an oscillating water bath at 37ºC, isolated cells were grown in 10% FCS/F-12 MEM, incubated at 37ºC under humid 5% CO2-air conditions, and passaged every 5 to 7 days. Aortic SMC between the thirth and sixth passages, obtained from three different primary cultures, were used for the experiments.
Characterization of Cultured SMC
Although cultured aortic SMC exhibited the characteristic hill-and-valley growth pattern on reaching confluence (Chamley-Campbell et al., 1981), cultures were identified by the presence of positive staining with anti-smooth muscle a-actin (Pang, 1989). A negative staining with anti-factor VIII antibodies assessed a complete removal of endothelial cells from aortic arteries.
Proteoglycan Evaluation
PGs were extracted and measured using techniques already described (Yanagishita et al., 1989) in the cultured VSMC medium and cell layer. A double _ isotopic label method with both [35S] sodium sulfate and [ 3H] glucosamine (New England Nuclear, Boston,MA) was used. The specific radioactivity of [ 3H] glucosamine is incorporated into all complex carbohydrates (total PGs) and the second radioisotope, [35S] sodium sulfate, labels only sulfated disaccharides.
ECM synthesis
Aortic SMC were plated on 12-well plates (4 x 104 cell/well) and cultured for 3 days in MEM/ F-12 supplemented with 10% FCS at 37ºC under humid 5% CO2-air conditions, until they reached subconfluence. VSMC were then serum-starved for 48 h in 0.1% FCS- MEM/ F-12 before stimulation. Quiescent cells were then incubated with the test materials: 10% FCS, 10 ng/ml platelet -derived growth factor (PDGF-BB), or as control group 0.1% FCS (n=8 in each), in the presence of both [35S]sodium sulfate ](10 mCi/well) and [ 3H] glucosamine (10 mCi/well) for 48 h.
Isolation of proteoglycans
Medium Extracellular Matrix (M-ECM): Secreted into the medium PGs
The medium was removed from each plate and the total volume was applied to a disposable Sephadex G-50 gel filtration column equilibrated and run in 4 mol/L guanidine hydrochloride, 0.05 mol/L sodium acetate (pH 6.0) and 2% (w/v)Triton X100. The corresponding eluted fraction was collected and analyzed for radioactivity.
Pericellular Matrix (P-ECM): Cell-associated PGs
The cell layer was rinsed in HBSS and extracted in 1 mL of 4 mol/L guanidine hydrochloride, 0.05 mol/L sodium acetate (pH 6.0) and 2% (w/v)Triton X 100. The solution containing the detached cells was chromatographed on Sephadex G-50 columns as described above.
In every experiment DNA synthesis for each treatment was determined by the Hoechst method.
The radioactivity of each medium or cell extract was counted simultaneously for [35S] and [3H] with a three over two dual label counting method in a liquid scintillation counter (BetaRack LKB Wallac, Finland).
Statistical and Data Analysis
Data (mean ± SEM) are presented as percent of the control incubation condition (0.1% FCS). Normal distribution of data was asssesed by Kolmogorov-Smirnov test so the statistical significance was assessed with two-way analysis of variance and the multiple comparisons tau test. A P value lesser than 0.05 was considered significant.
Results
The effect of the stimulation with different growth factors on secreted and membrane-bound total and sulfated PGs production in cultured aortic SMC from SHR and Wistar rats was examined. Incubation with 0.1% FCS was used as non-stimulated control in each group.
Figure 1A shows the results when [3H]-PGs were measured in the medium (medium ECM: M-ECM). In VSMC from both groups of rats the stimulation with PDGF-BB and 10% FCS increased significantly (P<0.001) extracellular [3H]-PGs compared with basal level (100%). VSMC from SHR displayed a significant greater synthesis of [3H]-PGs than those from Wistar rat cells, with both treatments. The results obtained when M-ECM [35S]-PGs production was evaluated are shown in Figure 1B. Both PDGF-BB and 10%FCS significantly increased [35S]-PGs in VSMC from SHR and control rats, without significative difference in [35S]-PGs production between SHR and Wistar cells in any case.
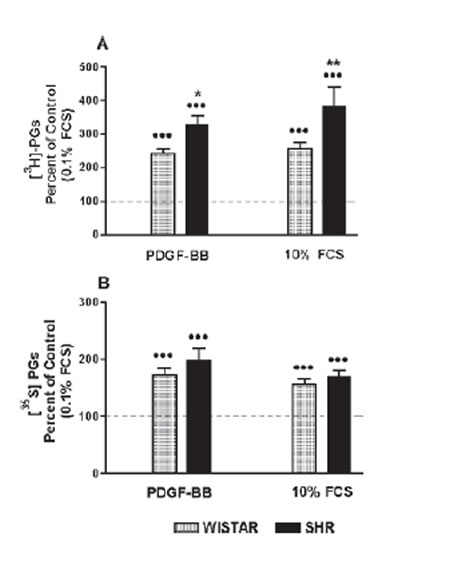
Figure 1. Effect of growth factors on the synthesis of PGs secreted into the medium (M-ECM). Uptake of [3H]-glucosamine (A) and [35S]-sodium sulfate (B) by cultured aortic SMC from SHR and Wistar rats incubated with 10 ng/mL PDGF-BB and 10% FCS. Data (mean ± SEM) presented as percent of a control group incubated with 0.1% FCS (100%). n= 8 in all groups. ••• P<0.001 vs. 0.1% FCS group. *P<0.05 and ** P<0.01 vs. Wistar rat cells.
The results obtained when cell-associated PGs (pericellular matrix: P-ECM) were measured are shown in Figure 2 A and B. In both VSMC groups, a significant increase in membrane-bound [3H]-PGs and [35S]-PGs compared with the control treatment (100%) was produced by PDGF-BB and 10% FCS. Both treatments elicited a significantly greater [3H]-PGs and [35S]-PGs production in VSMC from SHR than in Wistar rat cells.

Figure 2. Effect of growth factors on the synthesis of PGs secreted in the cell layer (P-ECM). Uptake of [3H]-glucosamine (A) and [35S]-sodium sulfate (B) by cultured aortic SMC from SHR and Wistar rats incubated with 10 ng/mL PDGF-BB and 10% FCS. Data (mean ± SEM) presented as percent of a control group incubated with 0.1% FCS (100%). n= 8 in all groups. •P<0.05, •• P<0.01 and ••• P<0.001 vs. 0.1% FCS group. **P<0.01 and *** P<0.001 vs. Wistar rat cells.
The sulfation of GAG chains was assesed by the sulfated/total PGs relationship, calculated by the [35S]-sulfate/[3H]-glucosamine ratio expressed as percent ([35S]dpm/ng DNA / [3H] dpm/ng DNA x 100) (Fig. 3). In Wistar rat and SHR cells, both PDGF-BB and 10% FCS treatments produced a decrease in sulfation of the medium PGs compared with the corresponding basal control group (0.1% FCS). The sulfated/total PGs ratio was significantly smaller in the medium from SHR VSMC, compared to Wistar rat cells, either in basal and stimulation conditions. A different pattern was observed in the case of the membrane-bound PGs , in which PDGF-BB and 10% FCS reduced sulfation ratio in Wistar cells, without effect on SHR cells. Besides, sulfation ratio was significantly smaller in control and stimulated cells from SHR than in Wistar rat cells.

Figure 3. Effect of growth factors on sulfation of PGs secreted into the medium (A) and bound to the membrane (B), as assessed by the [35S]-sulfate / [3H]-glucosamine ratio x 100, in cultured aortic SMC from SHR and Wistar rats, incubated with 0.1% FCS (control group), 10 ng/mL PDGF-BB and 10% FCS. Data are mean ± SEM, n=8 in all groups. •• P<0.01 and ••• P<0.001 vs. 0.1% FCS group. **P<0.01 and *** P<0.001 vs. Wistar rat cells.
Discussion
The mechanical properties of large arteries play a major role in cardiovascular haemodynamics through the buffering of stroke volume and the propagation of the pressure pulse (Safar and Frohlich, 1995). It is well recognized that mechanical properties of large arteries are primarily determined by the composition of the arterial wall and are dependent on the level of interaction between arterial wall components (cells and ECM). The ECM proteins, mainly collagen and elastin, influence the "passive" mechanical properties of the arterial wall whereas its "active" properties depend on the activation of VSMC.
It has been generally accepted that hypertension produced an increase in large-artery stiffness (Safar and Frohlich, 1995). The functional and structural modifications of the arterial wall have important effects on the cardiovascular system, increasing the incidence of fracture, rupture and aneurysm formation in arteries and, potentially, the development of atherosclerosis (Benetos et al., 1997). Sustained hypertension is associated with a rearrangement of the arterial wall material, including qualitative or quantitative changes in arterial components leading to the mechanical adaptation of the arterial wall. The arterial wall is not homogeneous and is composed of various elements, including VSMC and ECM components. All of these elements contribute to the mechanical behaviour of the wall material through their own elastic modulus and the way in which they are arranged. Local hormonal changes in hypertensive subjects may have pressure-independent effects on the arterial wall, mainly by modifying cell growth or synthesizing extracellular matrix. Nevertheless it is difficult to know whether the changes in arterial stiffness are pressure-related or due to intrinsic changes in the arterial wall or a combination of both factors. It seems that, for a given blood pressure level, arterial distensibility of the central arteries in hypertensives may be altered as a function of various enviromental and/or genetic factors which affect the composition of the extracellular matrix of the arterial wall.
The VSMC play a prominent role in development and maintenance of arterial structure. VSMC are the primary source of arterial ECM, including collagen, elastic fibers, and several PGs. In the normal media, VSMC are surrounded by ECM molecules and the interaction between VSMC and matrix components can significantly influence their ability to respond to growth factors and/or chemoattractans and can promote the transition of VSMC from a contractile to a synthetic phenotype (Hedin et al., 1999), which may play an essential role in the development of atherosclerotic and restenotic lesions (Rivard and Andrés, 2000).Conduit-artery stiffness in genetically hypertensive rats is influenced not only by hypertension but also by differences in the contents of collagen subtypes (Chamiot- Clerc et al., 1999). Extracellular constituents other than collagen and elastin, such as proteoglycans, may modulate vascular stiffness and VSMC functions. PGs serve several functions in the artery wall, including cell adhesion, migration, proliferation and differentiation (Ruoshlati and Yamaguchi, 1991; Jacob et al., 2001). PGs can also interact with cytokines, modifying its binding to their cell surface receptors and influencing the behaviour of cells (Schonherr and Hausser, 2000). The major PG in the arterial ECM synthetized by arterial VSMC is the large chondroitin sulfate proteoglycan versican, that bind hyaluronan and form large aggregates, which contribute to tissue mechanical properties, providing a hydrated sponge-like matrix that resists or cushions against deformation. Arteries smaller PGs contain dermatan sulfate glycosaminoglycans such as decorin and byglican, which interact with other ECM proteins and with macromolecules that enter the vascular wall such as low density lipoproteins (Chait and Wight, 2000). In addition, blood vessels contain perlecan, which is an heparan sulfate PG. In the normal adult human thoracic aorta, the main structural ECM components of the media include elastin, collagen types I, III, IV, V and VI, fibronectin, fibrillin-1, and proteoglycans, predominantly collagen-associated dermatan sulfate PG, cell-associated heparan sulfate PG, and interstitial chondroitin sulfate PG (Dingemans et al., 2000).
The present study was designed to examine the effect of growth factors on both secreted and membrane-bound total and sulfated PGs synthesis in cultured arterial smooth muscle cells obtained from a conduit artery, the thoracic aorta, of adult SHR and Wistar rats.
The present study demonstrates that the stimulation with PDGF-BB or 10% FCS induced a significantly greater amount of extracellular and pericellular total PGs, measured by [3H]-glucosamine incorporation, in aortic SMC from 12- to 14-week-old SHR than in cells from age-matched normotensive control rats. When sulfated PGs synthesis was analyzed, both treatments produced a significantly greater effect on membrane-bound PGs production in VSMC of SHR. However, there was no difference between SHR and Wistar rat cells in secreted to the medium sulfated PGs. Both PDGF-BB and 10% FCS induced a significant greater synthesis by VSMC from both rat strains in all cases, compared to non-stimulated basal values. These results indicate that VSMC obtained from thoracic aorta of genetically hypertensive rats, in a situation of growth stimulation, synthetize more total and sulfated proteoglycans than cells from normotensive animals.
We performed the experiments with cells from SHR because this animal is the most representative experimental model of human essential hypertension. Several changes in the VSMC of SHR has been described. Abnormal growth of aortic SMC in vitro is manifest as an accelerated entry into S phase of the cell cycle as well as a hyperresponsiveness to growth factors (Hadrava et al., 1989, 1991). Changes in apoptosis and proliferation of VSMC have been found in small intramyocardial arteries from the left ventricle of adult SHR (Diez et al., 1998). Aortic SMC migration induced by 10% FCS and PDGF was enhanced in young and adult SHR (Hsieh and Lau, 1998).
PGs were measured in both the cell layer and medium, as the medium does not necessarily reflect the conditions of the cell layer. According to Thogersen (Thogersen et al., 1996) the cell layer of cultured arterial SMC together with its associated extracellular matrix can be considered as a model of the arterial wall. Besides, the use of cultured VSMC allows us to study PGs synthesis by the tunica media, excluding synthesis by other vascular wall components. The VSMC growth stimulation was induced by two different treatments. As a complete growth factor, we used 10% FCS, which stimulatory effect on proliferation is clear under the same experimental conditions (Cruzado et al., 1998). On the other hand, PDGF is a critical autocrine and paracrine factor for VSMC mediating hyperplasia, hypertrophy and remodeling of the vascular wall (Berk, 2001; Lewis et al., 2001). It is also a potent chemotactic agent for VSMC and stimulates the production of extracellular matrix which, in turn, may regulate the proliferative and migratory capacity of the cells (Nelson et al., 1997; Raines et al., 2000). Evanko et al. (2001) have shown that PDGF modulates the synthesis and organization of VSMC pericellular coat-forming molecules such as versican, hyaluronan and link protein, which leads to ECM expansion and alterations in VSMC phenotype.
When the sulfation ratio of the GAG chains was analyzed in the medium, the increase in PG synthesis by aortic cells was accompanied by undersulfation, assesed by a minor [35S]/[3H] ratio, in the presence of PDGF-BB or a complete growth factor, such as 10% FCS, and in cells from both groups of rats. The same variation was observed in P-ECM PG synthesis in the Wistar rat cells but not in those from SHR. In both basal and growth stimulation conditions, cells from SHR displayed a lower [35S]/[3H] ratio in the medium and membrane-bound PGs than those of control rats. The relation between an increase in PG synthesis and undersulfation has been observed in other tissues, such as human fetal mesangial (Moran et al., 1991) and immature testis (Thiebot et al., 1997) cells. Present results differ from ours previously found in VSMC obtained from resistance mesenteric arteries (Castro et al., 1999), in which undersulfation appeared in both secreted and membrane-bound PG synthesis in SHR cells incubated with growth factors, but not in VSMC from Wistar rats. This difference in results could be explained by the concept of vascular heterogeneity. Indications that VSMC may be morphologically and functionally heterogeneous have recently appeared (Archer, 1996). Differences in proliferative capacity, electrophysiology, phenotypic marker protein content, matrix synthesis and expression of cell-specific transcription factors between individual VSMC and a diversity in contractile and pharmacologic properties of different smooth muscles are well recognized, and this diversity of VSMC may be a critical factor determining specific responses of smooth muscles to a number of physiological and pathophysiological stimuli (Halayko et al., 1997). According to Daemen and De Mey (1995) several types and levels of vascular heterogeneity may be distinguished, such as intervascular and intravascular heterogeneity. Intravascular cellular heterogeneity is present in normal and injured arteries, and its characteristics and mechanisms of production has been extensively reviewed (Shanahan and Weissberg, 1999; Frid et al., 1997). Previously we have described a different response to the growth promoting effect on insulin on VSMC isolated from aorta and mesenteric arteries from normal rats (Cruzado et al., 1998). The difference in undersulfation of the synthetized PGs by non-stimulated and stimulated VSMC from such functionally and structurally different arteries as mesenteric bed and aorta, obtained from SHR and Wistar rats, is a further evidence on intervascular heterogeneity. The concept of inter and intravascular heterogenity, that is, the variety of VSMC populations from segment to segment within an artery or among vascular trees of different organs, could explain the fact that vessels from different vascular beds exhibit different responses to a common stimulus, and it would be useful to explain the heterogeneity of arterial structural changes seen in hypertension and injury.
In summary, the present study indicate that cultured aortic SMC from spontaneously hypertensive rats synthetized more PGs in growth stimulating conditions than cells from normotensive rats. Results also show that this increase of PGs synthesis was accompanied with a greater undersulfation of GAG chains in both non- and stimulated conditions, indicating a different modulation of proteoglycan metabolism in this strain. The relationship between VSMC and the matrix elements, including proteoglycans, is thought to be a controlling factor in the elastic properties of the arterial wall. Besides, matrix proteoglycans play fundamental roles in the vessel wall, regulating vascular cell proliferation, migration and survival, modulating vascular stiffness and participating in cytokines activity. Present results may indicate that, by these or other actions, the changes in proteoglycans synthesis and modulation in smooth muscle cells of a conduit artery may be one of the pathophysiological mechanisms leading to vascular wall changes occuring in hypertension and other associated vascular diseases as atherosclerosis.
Acknowledgements
This study was supported by a grant from SeCyT-UNC (06/J131).
References
Archer SL (1996). Diversity of phenotype and function of vascular smooth muscle cells. J Lab Clin Med. 127: 524-529. [ Links ]
Benetos A, Kaurent S, Asmar RG, Lacolley P (1997). Large artery stiffness in hypertension. J Hypertens. 15 (suppl2): S89-S97. [ Links ]
Berk BC (2001). Vascular smooth muscle growth: autocrine growth mechanisms. Physiol Rev. 81: 999-1030. [ Links ]
Carey D (1991). Control of growth and differentiation of vascular cells by extracellular matrix proteins. Ann Rev Physiol. 53: 161-177. [ Links ]
Castro CM, Cruzado MC, Miatello RM, Risler NR (1999). Proteoglycan production by vascular smooth muscle cells from resistance arteries of hypertensive rats. Hypertension. 34 (part 2): 893-896. [ Links ]
Chait A, Wight TN (2000). Interaction of native and modified low-density lipoproteins with extracellular matrix. Curr Opin Lipid. 11: 457-463. [ Links ]
Chamiot-Clerc P, Renaud JF, Blacher J, Legrand M, Samuel JL, Levy BI, Sassard J, Safar ME (1999). Collagen I and III and mechanical properties of conduit arteries in rats with genetic hypertension. J Vasc Res. 36: 139-146. [ Links ]
Chamley-Campbell JH, Campbell JR, Ross R (1981). Phenotype-dependent response of cultured aortic smooth muscle to serum mitogens. J Cell Biol. 89: 379-383. [ Links ]
Cruzado M, Risler N, Castro C, Ortiz A, Rüttler M (1998). Proliferative effect of insulin on cultured smooth muscle cells from mesenteric resistance vessels. Am J Hypertens. 11: 54-58. [ Links ]
Daemen MJAP, De Mey JGR (1995). Regional heterogeneity of arterial structural changes. Hypertension. 25 (part I): 464-473. [ Links ]
Diez J, Fortuno MA, Zalba G, Etayo JC, Ravassa S, Beaumont J (1998). Altered regulation of smooth muscle cell proliferation and apoptosis in small arteries of spontaneously hypertensive rats. Eur Heart J. 19 Suppl G: G29-33. [ Links ]
Dingemans KP, Teeling P, Lagendijk JH, Becker AE (2000). Extracellular matrix of the human aortic media: an ultrastructural histochemical and immunohistochemical study of the adult aortic media. Anat Rec. 258: 1-14. [ Links ]
Dubey RK, Jackson EK, Rupprecht HD, Sterzel RB (1997). Factors controlling growth and matrix production in vascular smooth muscle and glomerular mesangial cells. Curr Opin Nephrol Hypertens. 6: 88-105. [ Links ]
Emoto N, Onose H, Yamada H, Minami S, Tsushima T, Wakabayashi I (1998). Growth factors increase pericellular proteoglycans independently of their mitogenic effects on A10 rat vascular smooth muscle cells. Int J Biochem. 30: 47-54. [ Links ]
Evanko SP, Johnson PY, Braun KR, Underhill CB, Dudhia J, Wight TN (2001). Platelet-derived growth factor stimulates the formation of versican-hyaluronan aggregates and pericellular matrix expansion in arterial smooth muscle cells. Arch Biochem Biophys. 394: 29-38. [ Links ]
Frid MG, Dempsey EC, Durmowicz AG, Stenmark KR (1997). Smooth muscle heterogeneity in pulmonary and systemic vessels. Importance in vascular disease. Arterioscler Thromb Vasc Biol. 17: 1203-1209. [ Links ]
Hadrava V, Tremblay J, Hamet P (1989). Abnormalities in growth characteristics of aortic smooth muscle cells in spontaneously hypertensive rats. Hypertension. 13: 589-597. [ Links ]
Hadrava V, Kruppa U, Russo RC, Lacourciere Y, Tremblay J, Hamt P (1991). Vascular smooth muscle cell proliferation and its therapeutic modulation in hypertension. Am Heart J. 122 (4 Pt2): 1198-11203. [ Links ]
Halayko AJ, Rector E, Stephens NL (1997). Characterization of molecular determinants of smooth muscle cell heterogeneity. Can J Physiol Pharmacol. 75: 917-929. [ Links ]
Hedin U, Roy J, Tran PK, Lundmark K, Rahman A (1999). Control of smooth muscle proliferation - the role of the basement membrane. Thromb Haemost. 82 Suppl 1: 23-26. [ Links ]
Hein M, Fisher J, Kim DK, Hein L, Pratt RE (1996). Vascular smooth muscle phenotype influences glycosaminoglycan composition and growth effects of extracellular matrix. J Vasc Res. 246: 279-292. [ Links ]
Hsieh CC, Lau Y (1998). Migration of vascular smooth muscle cells is enhanced in cultures derived from spontaneously hypertensive rat. Plugers Arch. 435(2): 286-292. [ Links ]
Intengan HD, Schiffrin EL (2001). Vascular remodeling in hypertension: roles of apoptosis, inflammation and fibrosis. Hypertension. 38 (Pt2): 581-587. [ Links ]
Intengan HD, Thibault G, Li JS, Schiffrin EL (1999). Resistance artery mechanics, structure, and extracellular components in spontaneously hypertensive rats: effects of angiotensin receptor antagonism and converting enzyme inhibition. Circulation. 100: 2267-2275. [ Links ]
Jacob MP, Badier-Commander C, Fontaine V, Benazzoug Y, Feldman L, Michel JB (2001). Extracellular matrix remodeling in the vascular wall. Pathol Biol. 49: 326-332. [ Links ]
Lee RT, Yamamoto C, Feng Y, Potter-Perigo S, Briggs WH, Landschulz KT, Turi TG, Thompson JF, Libby P, Wight TN (2001). Mechanical strain induces specific changes in the synthesis and organization of proteoglycans by vascular smooth muscle cells. J Biol Chem. 276: 13847-13851. [ Links ]
Lewis CD, Olson NE, Raines EW, Reidy MA, Jackson CL (2001). Modulation of smooth muscle proliferation in rat carotid artery by platelet-derived mediators and fibroblast growth factor-2. Platelets. 12: 352-358. [ Links ]
McLeod DC, Strauss BH, de Jong M, Escaned J, Umans V, van Suylen RJ, Verkerk A, de Feyter PJ, Serruys PW (1994). Proliferation and extracellular matrix synthesis of smooth muscle cells cultured from human coronary atherosclerotic and restenotic lesions. J Am Coll Cardiol. 23: 59-65. [ Links ]
Moran A, Brown DM, Kim Y, Klein DJ (1991). Effects of IGF-I and glucose on protein and proteoglycan synthesis by human fetal mesangial cells in culture. Diabetes 40: 1346-1354. [ Links ]
Nelson PR, Yamamura S, Kent KC (1997). Platelet-derived growth factor and extracellular matrix proteins provide a synergistic stimulus for human vascular smooth muscle migration. J Vasc Surg. 26: 104-112. [ Links ]
Pang SC (1989). Proliferation of aortic smooth muscle cells of genetically hypertensive and normotensive rats in culture. J Pathol. 158: 167-173. [ Links ]
Raines EW (2000). The extracellular matrix can regulate vascular cell migration, proliferation, and survival: relationships to vascular disease. Int J Exp Pathol. 81: 173-182. [ Links ]
Raines EW, Koyama H, Carragher NO (2000). The extracellular matrix dynamically regulates smooth muscle cell responsiveness to PDGF. Ann New York Acad Sci. 902: 39-52. [ Links ]
Risler N, Castro C, Cruzado M, González S, Miatello R (2002). Early changes in proteoglycans production by resistance arteries smooth muscle cells of hypertensive rats. Am J Hypertens. 15: 416-421. [ Links ]
Rivard A, Andrés V (2000). Vascular smooth muscle cell proliferation in the pathogenesis of atherosclerotic cardiovascular diseases. Histol Histopathol. 15: 557-571. [ Links ]
Ruoshlati E, Yamaguchi Y (1991). Proteoglycans as modulators of growth factors activities. Cell. 64: 867-869. [ Links ]
Safar M, Frohlich E (1995). The arterial system in hypertension: a prospective view. Hypertension. 26: 10-14. [ Links ]
Schonherr E, Hausser HJ (2000). Extracellular matrix and cytokines: a functional unit. Dev Immunol. 7: 89-101. [ Links ]
Shanahan CM, Weissberg PL (1999). Smooth muscle cell phenotypes in atherosclerotic lesions. Curr Opin Lipidol. 10: 507-513. [ Links ]
Thiebot B, Bichoualne L, Langris M, Bonnamy PJ, Barbey P, Carreau S, Bocquet J (1997). IGF-1 stimulates synthesis of undersulfated proteoglycans and of hyaluronic acid by peritubular cells from immature rat testis. Biochim Biophys Acta. 1358: 127-141. [ Links ]
Thogersen VB, Keickendorff L, Ledet T (1996). Effect of insulin and growth hormone on the synthesis of radiolabelled proteoglycans from cultured human arterial smooth muscle cells. Eur J Endocrinol. 134: 326-330. [ Links ]
Tuñón J, Ruiz-Ortega M, Egido J (2000). Regulation of matrix proteins and impact on vascular structure. Current Hypertension Reports. 2: 106-113. [ Links ]
Yanagishita M, Salustri A, Hascall V (1989). Specific activity of radiolabeled hexosamines in metabolic labeling experiments. Methods in Enzimology. 179: 435-445. [ Links ]
Yayon A, Klagsbrun M, Esko JD, Leder P, Ornitz DM (1991). Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell. 64: 841-848. [ Links ]
Received on September 30, 2002.
Accepted on February 28, 2003.

