










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBiocell
versión impresa ISSN 0327-9545
Biocell v.28 n.2 Mendoza abr./ago. 2004
Mouyu Yang1 and V.Benno Meyer-Rochow1,2
1. School of Engineering and Science, International University Bremen, D-28725 Bremen, Germany.
2. Department of Biology, University of Oulu, P.O.Box 3000, SF-90014 Oulu, Finland.
Address correspondence to: Dr. V.B. Meyer-Rochow. School of Engineering and Science, International University Bremen (IUB), P.O.Box 75 05 61, D-28725 Bremen, GERMANY. Fax: (+49-421) 200 3249. Tel: (+49-421) 200 3242. E-mail: b.meyer-rochow@iu-bremen.de
Abstract: Only one insect (the scale insect Eriococcus sp.) is known, in which photoreceptive lamellae appear to have replaced the usual arthropod rhabdom microvilli. We are now reporting the presence of photoreceptive membranes, which also appear to resemble lamellae rather than microvilli, but they are in the ocellus of the tiny wasp Centrodora sp., which parasitizes scale insect eggs. The apparently optically homogenous lens of the Centrodora ocellus measures approximately 10 µm in diameter and, thus, operates at the limits of diffraction. We calculated that the lens is capable of focusing a parallel bundle of rays on the retina of the ocellus.
Key words: Insect retina. Eye. Ocellus. Parasitoid. Visual membrane.
Introduction
Arthropod compound eyes have extensively been studied and it seems that all of them possess photoreceptors that, according to Eakin (1979) are "rhabdomeric" (= microvillar) in origin. Arthropod dorsal ocelli (Toh and Tateda, 1991; Mizunami, 1994; Yoon et al., 1996) and stemmata (= larval eyes: Meyer-Rochow, 1974; Cole, 1994) have received considerably less attention. Yet, it is clear that they too, with one exception (Duelli, 1978), possess photoreceptive membrane specializations that can be considered microvillar in nature.
The one exception, mentioned above, concerns the ocelli of the male scale insect Eriococcus sp., in which membrane stacks "whose configuration is analogous to the stacked plates of vertebrate cones" (Duelli, 1978) replace the customary microvilli. Duelli (1978) speculated on the reasons for the presence of these stacked membrane lamellae and came to the conclusion that the lamellar arrangements did not provide a more extensive area than that made available by microvilli for the accommodation of photosensitive pigments. The small size of the ocelli also did not seem to be an adequate explanation for the presence of membrane stacks. Why the latter occurred in the ocellus of the male scale insect Eriococcus sp. had to remain an open question.
We have now examined the ocelli of the tiny, only about 0.5 mm long parasitic wasp Centrodora sp. It parasitizes eggs of various scale insects and appears to show some similarity in ocellar ultrastructure to that of its host.
Materials and Methods
Numerous tiny specimens of the parasitic wasp Centrodora sp (fam. Aphelinidae), measuring no more than 0.5 mm in total body length (Fig. 1), were fixed whole in cold 3% phosphate buffered glutaraldehyde primary fixative. Because the specimens were fixed in their totality and the removal of just the eyes was not possible, the fixation results are not entirely optimal. Following several rinses in buffer, the specimens were then postfixed for 2 h in 1% OsO4 at room temperature. After a renewed brief rinse the specimens were dehydrated in a graded series of acetone and embedded in Spurrs resin. Sections, 1 µm thick for light microscopy, and golden (i.e., 90-150 nm) for electron microscopy were cut with a glass knife on a Reichert OM U2 ultramicrotome and stained either with toluidine blue (for light microscopy) or uranyl acetate and lead citrate (electron microscopy) for a few minutes. Carbon-coated 300-mesh grids were used to pick up the ultrathin sections.
FIGURE 1. Illustration of a typical parasitic wasp of the genus Centrodora (in this case of C. speciosissima: From Borror et al., 1989).
Results and Discussion
The ocelli of Centrodora sp. are equipped with a transparent, biconvex lens, the diameter of which measures approximately 10 µm and whose outer and inner radii of curvature are Rd = 5.4 µm and Rp = 5.5 ìm, respectively (Figs. 2, 3). Beneath the corneal lens, large amounts of dark screening pigment, contained in oval granules of approximately 0.7 x 0.3 µm dimension surround centrally-placed photoreceptive membranes (Figs. 2, 4). The membranes closely resemble the stacked plates known from the ocellus of the scale insect Eriococcus sp. (Fig. 5) and, moreover, also contain central cores of cytoplasm just like the scale insect ocellus (compare Figs. 4 and 5).
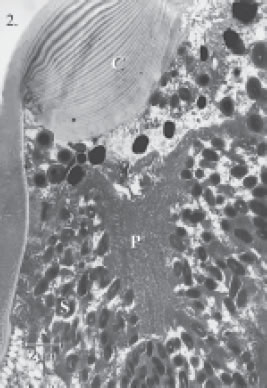
FIGURE 2. Longitudinal section through the dorsal ocellus of Centrodora sp., showing biconvex corneal lens (C), photoreceptive membranes (P), and grains of screening pigment (S).

FIGURE 3. Diagrammatic representation of corneal lens with radii of curvature entered and three focal points (A, B, and C) calculated for lens refractive indices of 1.45, 1.50, and 1.55, respectively.
Each membrane disc is approx. 0.045 µm thick and separated from its neighbours by a space of 0.012 µm. Screening pigment granules, like those of the scale insect ocellus, are not invading the photoreceptive membrane layers, but occupy places below and around the photoreceptor outer segment layer. The depth, i.e. the distance the photoreceptive membranes occupy from their most distal to most proximal ends is approximately 5 µm and again resembles that reported from the ocellus of the scale insect. However, the total volume of visual membrane material in Centrodora is very considerably smaller than that of the scale insect, in which outer segments are not confined to the focal region of the lens, but occupy the entire inside surface area of the spherical lens to a thickness of 3 µm. The lens in Centrodora also seems less specialized than that of the scale insect, in which an F-number as small as 0.55 was calculated. The differences could be a reflection of the fact that Centrodora in addition to the ocelli possesses lateral compound eyes (with obvious microvilli rather than lamellae) and, thus, depends less on the ocelli with regard to vision than the scale insect, which has no compound eyes and, therefore, relies only on the ocelli for photoreception.
Using geometrical optics for paraxial rays according to the Müller-Matrix method (cf. Saleh and Teich, 1991), we calculated for the Centrodora ocellus focal distances for three optical conditions involving its optically homogenous, biconvex lens of known dimension. Assuming refractive indices of NA = 1.45, NB = 1.50, and NC = 1.55 for the corneal cuticle and N = 1.35 for the more aqueous cytoplasm on the proximal side of the lens, we find the corresponding focal points A, B, and C at approx. 9, 7.5, and 6.5 µm behind the inner surface of the lens. The resultant F-numbers (focal distance/lens diameter) would be 0.9, 0.75, and 0.65. Considering that there is an approximately 1-2 µm thick membrane-free space beneath the corneal lens, occupied by clear cytoplasm, at least focal points B and C would fall upon photoreceptive membranes. That a figure of even 1.55 for the assumed refractive index of the corneal lens material is not too high an estimate, becomes obvious, when we compare the figure with measurements that were obtained on fresh insect material and determined with the aid of an interference microscope (N = 1.545 in the larval eye of the sawfly Perga: Meyer-Rochow, 1974; N = 1.534 in the cornea of the beetle Euoniticellus: Meyer-Rochow, 1978).
That focal points exist at all and lie within the layer of the photoreceptive membranes is an indication of the quality of the lens and the need of the eye to perceive a certain degree of focused light. It does not, however, allow us to conclude that the insect makes use of image formation, because to resolve even a crude image probably requires significantly more receptors than those that are present in the Centrodora ocellar retina. In fact, with a diameter of only 10 µm the lens of Centrodora, like that of some other small arthropods (Meyer-Rochow and Reid, 1996; Meyer-Rochow and Bobkova, 2002), operates near the limits of distortion through diffraction (Barlow, 1952). However, some larger parasitic hymenopteran parasitoids (for example Microplitis croceipes) have been shown to involve vision during processes of learning (Wäckers and Lewis, 1999).

FIGURE 4. Higher magnification of photoreceptive membrane layers (P) alternating with cytoplasmic strands resembling what has been described from the ocellus of the scale insect host (cf. Fig. 5).
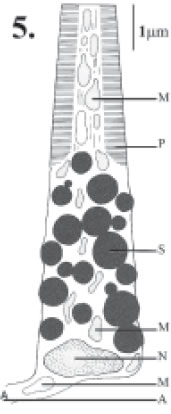
FIGURE 5. Schematic longitudinal section of a photoreceptor cell in the ocellus of the male scale insect Eriococcus sp. (after Duelli, 1978).
The similarity between the ocellar fine structure of the eye of the male scale insect Eriococcus sp. and that of its minute egg parasite is intriguing. Can it be more than mere coincidence? Both are very small insects and since the eyes and ocelli of large numbers of very small insects have not been scrutinized to any extent, it may well be that similar structural organizations turn up in the eyes of other tiny insect species. Yet, some of the tiniest arthropods whose eyes have been examined ultrastructurally, for example those of an only 0.35 mm long mite, do possess photoreceptive membranes of microvillus nature (Meyer-Rochow and Bobkova, 2002). Clearly, the answer to why photosensitive lamellae rather than microvilli might be developed in a few isolated arthropod taxa eludes us, but could the similarity between host and parasite photoreceptor be an indication of the possibility of horizontal gene transfer?
Effects of parasitization by an ectoparasitic wasp have been described (Richards and Edwards, 1999) and even insect-to-insect gene transfers are known (Handler, 2001; Werren et al., 1995; Houck et al., 1991). All that is required for this to be successful is a vector, perhaps in the form of a virus and there are numerous insect viruses that could be potentially effective vectors (Boucias and Pendland, 1998). However, one ought to expect only minor changes in the phenotype as a consequence of this kind of gene transfer. Features controlled by multiple genes are not likely to be horizontally transmitted, but regulator genes affecting minor structural or functional changes are a different thing. A change from a tubelike microvillar membrane to a flattened lamella does not seem to require a great deal of modification. However, the lamellae created in this way would certainly not be homologous to the flattened stacks of membrane discs characteristic of eyes of Eakins ciliary line, i.e., vertebrates, echinoderms, chaetognaths, and coelenterates (Eakin, 1968).
Acknowledgements
We wish to thank Dr. Philippa Gerard for providing us with specimens for this study, Dr. Marina Bobkova for her help with the sectioning of the material, and Dr. Jozsef Gal for the optical measurements and calculations.
References
Barlow HB (1952). Size of ommatidia in apposition eyes. J Exp Biol 29: 667-674. [ Links ]
Boucias DG, Pendland JC (1998). Principles of insect pathology. Kluwer Academic, Boston. [ Links ]
Borror DJ, Triplehorn CA, Johnson NF (1989). An introduction to the study of Insects. Saunders College Publishers, Philadelphia. [ Links ]
Cole G (1994). Form and function of stemmata in larvae of holometabolous insects. Ann Rev Entomol 39: 323-349. [ Links ]
Duelli P (1978). An insect retina without microvilli in the male scale insect, Eriococcus sp. (Eriococcidae, Homoptera). Cell Tissue Res 187: 417-427. [ Links ]
Eakin RM (1968). Evolution of photoreceptors. In: General physiology of cell specializations. D Mazia and A Taylor, Eds, McGraw Hill, New York, pp 393-425. [ Links ]
Eakin RM (1979). Evolutionary significance of photoreceptors: in retrospect. Amer Zool 19: 647-653. [ Links ]
Handler AM (2001). A current perspective on insect gene transformation. Insect Biochem. Mol Biol 31: 111-128. [ Links ]
Houck MA, Clark JB, Peterson KR, Kidwell MG (1991). Possible horizontal transfer of Drosophila genes by the mite Proctolaelaps regalis. Science 253: 1125-1129. [ Links ]
Meyer-Rochow VB (1974). Structure and function of the larval eye of the sawfly Perga sp. (Pergidae, Hymenoptera). J Insect Physiol 20:1565-1591. [ Links ]
Meyer-Rochow VB (1978). Retina and dioptric apparatus of the dung beetle Euoniticellus africanus (Scarabaeidae). J Insect Physiol 24:165-179. [ Links ]
Meyer-Rochow VB, Bobkova MV (2002). Fine structure of the lateral eyes in the tiny marine Antarctic mite Halacarellus thomasi: an ultrastructural study. Polar Biol 25: 512-516. [ Links ]
Meyer-Rochow VB, Reid WA (1996). An eye for the extreme: photoreceptor fine structure in the Antarctic midge Belgica antarctica (Dipera: Chironomidae). Appl Entomol Zool 31: 631-634. [ Links ]
Mizunami M (1994). Information processing in the insect ocellar system: comparative approaches to the evolution of visual processing and neural circuits. Adv Insect Physiol 25, 152-265. [ Links ]
Richards EH, Edwards (1999). Parasitization of Lacanobia oleracea (Lepidoptera: Noctuidae) by the ectoparasitic wasp, Eulophus pennicornis ß effects of parasitization, venom and starvation on host haemocytes. J Insect Physiol 45: 1073-1083. [ Links ]
Saleh BEA, Teich MC (1991). Fundamentals of photonics. John Wiley & Sons, New York, Toronto. [ Links ]
Toh Y, Tateda H (1991). Structure and function of the insect ocellus. Zool Sci 8: 395-413. [ Links ]
Wäckers FL, Lewis WJ (1999). A comparison of color-, shape- and pattern-learning by the hymenopteran parasitoid Microplitis croceipes. J Comp Physiol 184: 387-393. [ Links ]
Werren JH, Zhang W, Guo LR (1995). Evolution and phylogeny of Wolbachia – reproductive parasites of arthropods. Proc Roy Soc Lond 261: 55-63. [ Links ]
Yoon C-S, Hirosawa K, Suzuki E (1996). Studies on the structure of the ocellar photoreceptor cells of Drosophila melanogaster with special reference to subrhabdomeric cisternae. Cell Tissue Res 284: 77-85. [ Links ]
Received on November 25, 2003.
Accepted on March 11, 2004.

