










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBiocell
versión impresa ISSN 0327-9545
Biocell v.28 n.3 Mendoza ago./dic. 2004
Differentiation and morphogenesis of Triatoma infestans (Klug 1834) female gonads. I – Post embryonic development
Cecilia I. Ibañez de Barrett, Juan Pablo Bozzini, and Marta Mariano de Bozzini*
A.N.L.I.S. Dr. Carlos G. Malbrán. Avda. Velez Sarsfield 563, (1281) Buenos Aires, Argentina.
*Member of the Research Career. National Research Council, Argentina.
Address correspondence to: Dra. Marta Mariano de Bozzini. A.N.L.I.S. "Carlos Malbrán", Virrey Cevallos 385 - 7º 40, (1077) Buenos Aires, ARGENTINA. FAX: (+54-11) 4303-1433.
E-mail: fliabozzini@hotmail.com
ABSTRACT: The post-embryonic development of the female gonads in Triatoma infestans (Hemiptera, Heteroptera), insects of importance in health affairs as harbors and vectors of different tripanosomatidea flagellates, is presented in a complete follow-up since insect hatches from the egg up to the last molt in the fifth instar stage. The detailed description of the morphological changes which occur in each instar as well as careful measurements evaluating its size increase have been analyzed by stereomicroscopy, phase contrast, dark field, and oblique illumination, in order to optimize the observations as well the photographic register of gonad morphology and structure.
The analysis was performed on gonad specimens obtained from broods no less than twenty (20) nymph bugs, reared at constant temperature and fed-up regularly.
According to the results of our study we can assert that gonad differentiation takes place in early phases of the insect development. Such is the case that first instars nymph’s present absolutely and easily recognizably male and female gonads.
From the third instar on beside the filament region, the three zones in each ovariole body is distinguished, a differentiation that is more noticeable during the fourth instar where a definite organization is present at the vitelarium. Such a clear cut zone development continues intensively during the fifth instar.
Finally at the end of such fifth nymph stage and when the last molt toward adults is prepared, clear signs of ovariole maturation take place since oocytes in early vitelogenesis are found.
Key words: Ovary, Development, Hemiptera, Insect Nymph
Abbreviations used in the illustrations:
est: stroma; f: suspensor filament; ft: terminal filament; ov: oocytes; ovd: oviduct; ovl: ovariole; pd: pedicel; tr: trachea (s); tro: tropharium; tz: transition zone; vt: vitelarium;
Introduction
There are few previous reports in the literature regarding the study of the morphogenesis and embryonic development of the female gonad or ovary in Hemiptera. In this sense the work of Bonhag (1955) on Oncopeltus fasciatus, of Case (1970), Huebner and Anderson (1972a) on Rhodnius prolixus, should be
mentioned.
Little is known about the morphogenesis of gonads, not only in Hemiptera but also in all the taxa with telotrophic ovaries. The complexity of the structure of this type of ovary is probably responsible for the limited number of studies available on differentiation, embryonic growth and morphologic variation of the ovary during the instar stages (Barth, 1973; Richard and Davies, 1983; Wigglesworth, 1951).
In order to gain a deeper knowledge of the morphogenesis of the gonad, we have carried out a thorough sequential study in Triatoma infestans, since we had previously studied the maturation of the female gamete in the adult specimen (Ibañez et al., 1999).
With regard to Hemiptera Reduvidae, group including among others Triatoma infestans and Rhodnius species, (Galliard, 1935; Gillet, 1935; Lutz and Huebner, 1978); the data reported by these authors remain unclear. Added to this fact, each species may have unique features, so it is not feasible to extrapolate the data obtained to other species.
Ever since the pioneer work of Del Ponte (1921) in Triatoma, the main interest with regards to the differences between male and female insects, was focused on the observation and description of the external sexual characteristics both in adult and instar stage insects (Canale and Carcavallo, 1985; Espínola, 1966; Gonçalves et al., 1985; Lent and Wygodzinsky, 1979).
This article presents a study of the morphogenesis changes of T. infestans´ female gonad during each of the five nymphal stages. The anatomic features and a brief albeit complete view of the histological structures are shown; a more detailed analysis, which includes the submicroscopic structure, is part of another study in progress.
Material and Methods
Biologic material
The nymphal gonads were obtained by dissection performed immediately after hatching or molting in each stage with no food intake between molting and dissection. In the 4th instar stage dissections were performed immediately after molting from the third, and few hours prior to the next instar molt taking into account the presence of a new cuticle under the old one to be abandoned by the insect reaching the new instar stage.
Before dissecting the insects were anesthetized for a few seconds with chloroform, then placed with their back upwards on a paraffin bottom Petri dish and immobilized with entomology pins.
Fixation
Once the chitin cuticle of the back had been removed (under stereoscopic binocular microscope) and the internal organs were exposed, an in situ fixation was performed with freshly prepared Karnovsky´s fixative, at a temperature below 10ºC. The specimen was then placed in a small flask containing the same fixative for no less than 30 minutes, in a bath at approximately 8ºC, so that immersion fixation may proceed. Once the fixation stage was completed the gonads were separated on a Petri dish and the necessary incisions performed, mainly at the level of the suspending filament and/or terminal duct (the dissection should be done with great care in order to avoid damaging the extremely fragile gonads). The separated gonads were placed in small flasks with the same fixative, and kept in a cold bath for two hours. Post fixation with 1% osmium tetra-oxide also achieves a contrasting effect on the sample.
Taking advantage of the experience we have accumulated in the recognition of gonads in adult specimens and considering that the size of the insect and the organ facilitates its identification, we were able to perform a sequential study, in a decreasing order, of each instar stage and recognize the corresponding structure.

FIGURE 1
Inclusion, sectioning, staining and embeding
The specimens were dehydrated using a series of graded alcohols (ethanol), then rinsed twice in propylene oxide. Samples of each instar stage were embedded in Poly-Bed Araldite resins, according to Mollenhauert (1964).
Semi-thin sections (2mm) obtained by the ultramicrotome were stained with a mixture of Azure II- methylene blue and mounted with the same inclusion resin for optic microscope observation.
Results
The procedure used to recognize the gonads consisted in a comparative study of these organs in a decreasing order, each instar stage with those of the previous one, that is, from the last stage (5th instar) to the first one.
Although the gonad is a structure that can easily be mistaken with the surrounding tissue, particularly in the first stages, it is possible to identify it as an independent structure if certain parameters are taken into account: location, shape and relative size.
Located between the digestive tract wall and the tegument, the gonad is found associated to the fatty body and components of the respiratory system, both present in all of the insects’ body cavity. On the one hand, the texture of the fatty body seen as a loose web of fatty lobes and, on the other, the bigger or smaller tracheal tubes, are criteria that help us to recognize a different structure, somewhat compact although very fragile, that looks like a flattened sac and has a different light refraction index when compared to the surrounding tissues. When the dissected specimen is observed under a stereoscopic microscope - with a magnification higher than 30x - the gonads can be identified as a pair of structures located symetricaly in the abdomen. The examination of these structures under phase contrast, dark field and oblique illumination optical microscopy, will show the following images:
1.- The structures that belong to the same specimen have seven equivalent formations, of the same size and characteristic general morphology.
2.- The structures from distinct specimens can be separated into two groups, according to the morphologic differences found in their components. Female and male gonads are thus recognized. In Triatoma we have established such differences from the 1st instar stage on (Fig. 2A, 2B).

FIGURE 2. T. infestans (Klug 1834) Photomicrograph exposed at the same magnification giving a relationship of male (A) and female (B) gonads size at first nymph instar few hours after hatching. (A) The arrow points one of the seven testicular follicles conforming the male gonad Phase contrast. Female gonad (B) shown under dark field illumination with a superimposed negative object micrometer (1mm ¸ 100sections). The seven primordial ovarioles are easily distinguished. Each micrometer division match up 10 mm. Male gonad has always a little larger size.
The size variations of each component (ovariole) of the female gonad (ovary) in each of the five nymphal stages are shown in the graphic (Fig. 1).
First instar stage – (Fig 3A)
An elongated gonad is observed, its size is shown in Figure 1. The body of the ovary is formed by seven ovarioles, laid out in an almost parallel way, clearly separated from one another. All the ovariole terminal filaments converge at the front end to form the suspensor filament. The caudal extremity of the ovarioles is less defined; nevertheless, each of them is oriented towards the back and extend to form a primary duct (oviduct) (Fig. 3B, 3C) In the space between ovarioles the cells that make up the stroma of the gonad are found.
FIGURE 3. T. infestans (Klug 1834). A: first instar nymph 10:1. B: The whole ovary of the first instar nymph shown under oblique illumination. C: Another ovary as it appears under phase contrast, the seven ovarioles can be differentiated. D: Second instar ovary photomicrograph under phase contrast showing the seven ovarioles. E: Another ovary shown under oblique illumination. F: second instar nymph 8,5:1.
At the histological level, we can distinguish in each ovariole three regions: the filament, which is quite big, a middle region or body of the ovariole and a terminal region or pedicel (Fig. 6A).
As already mentioned, in this stage we can differentiate the male and female gonads and the first observable difference, as shown by the stage micrometer, is the slightly bigger size of the former (Fig. 2A, 2B).
Second instar stage – (Fig. 3F)
The form of the gonad is slightly oval and elongated, becoming thinner at both extremes. As in the first stage, the body of the seven ovarioles, each of them surrounded by their basal lamina, is perfectly outlined. Each group of seven ovarioles is oriented and converge towards the genital duct. The separation between them has decreased (Fig. 3D, 3E).
Under a phase contrast microscope we can observe that the ovarioles present a thinner filament region. In each ovariole body two regions can already be distinguished: the tropharium and a not yet differentiated region; the pedicel has approximately the same size as that of the 1st stage The stroma of the gonad is best seen at the level of the ovariole tropharium. (Fig. 6B) The whole organ is surrounded and encircled by branched small tracheas (tracheoles). The quantity of tracheoles is moderate and their relationship to the organ is modified during the different stages.
Third instar stage – (Fig. 4A)
The shape of the ovary is similar to that of the previous one. The well outlined ovarioles are no longer separated and their convergence at the cephalic and caudal ends is more evident and increases the compact aspect of the ovary (Fig. 4B), with a well defined stroma. At the histological level the body of the ovariole has changed considerably and can be divided in regions:
tropharium, intermediate or transition zone and pedicel. The transition zone begins to be visible between the developing tropharium and the pedicel or ovariole peduncle (Fig. 6C). The tracheoles encircle the gonad with no visible change in their number as compared to the 2nd stage (Fig. 4B).
FIGURE 4. T. infestans (Klug 1834). A: Third instar nymph 7,5 : 1. B: The complete ovary of the third instar
photographed under dark field illumination. Six ovarioles are easily distinguished. C: Phase contrast micrograph
of an ovary of the fourth instar nymph. D: Fourth instar nymph 5 : 1.
Fourth instar stage
The general shape of the gonad is similar to that of the 3rd stage on a just molted insect, but the compact aspect of the ovary has increased. The ramified tracheoles which now completely surround the gonad have grown considerably and form a dense coating (Fig. 4C). At the histological level each ovariole is notably differentiated, showing a clear separation of the tropharium, the transition region and the incipient vitelarium (Fig. 6D). The filament zone increases its length and reduces its diameter, the pedicel is totally developed (Fig. 6G).
It can be observed that the tropharium cells have increased in number and reduced their size as compared to the previous stages; moreover, cell divisions can also be seen.
Fifth instar stage – (Fig. 5A).
The general shape and histology of the ovary in just molted nymphs is similar to that of the end of the 4th stage. In the ovarioles one can again observe a slight tendency to separation so the ovary looks less compact. The tracheal web which surrounds and encircles the ovary is very conspicuous and penetrates the organ (Fig. 5B, 5C, 5E).
FIGURE 5. T. infestans (Klug 1834). A: Fifth instar nymph 2,7 : 1. B: Fifth instar ovary as seen "in situ" after dissection, a profusion of tracheal tubes each of different diameter hide the gonad fine anatomy. C: Fifth instar ovary dissected and pealed out of the large diameter tracheal tubes in order to visualize the gonad anatomy. Four ovarioles and part of a fifth are distinguished. D: Adult female just molted 2,3 : 1. E: Tracheal sac frequently found on the tracheal tubes near the ovarioles. F: Ovary of a just molted adult which still is covered by part of the tracheal system. Only part of the respiratory ducts have been removed in order to show an oocytes{arrow}; (probably a vitelogenic one) Notice the size of the gonad and the presence of a nearly mature gamete!!.
In a specimen at the end of the 5th stage a process of reorganization of the tropharium cells occurs, which apparently exhibit features of adult nutritious cells, even before molting to the adult stage takes place (Fig. 6G). At the histological level we observe an obvious change in the growing vitelarium, where grown oocytes can be seen, showing signs of maturation in initial vitelogenesis, at the time when molting to the adult stage soon will take place, even when no feeding took place (Fig. 6H).
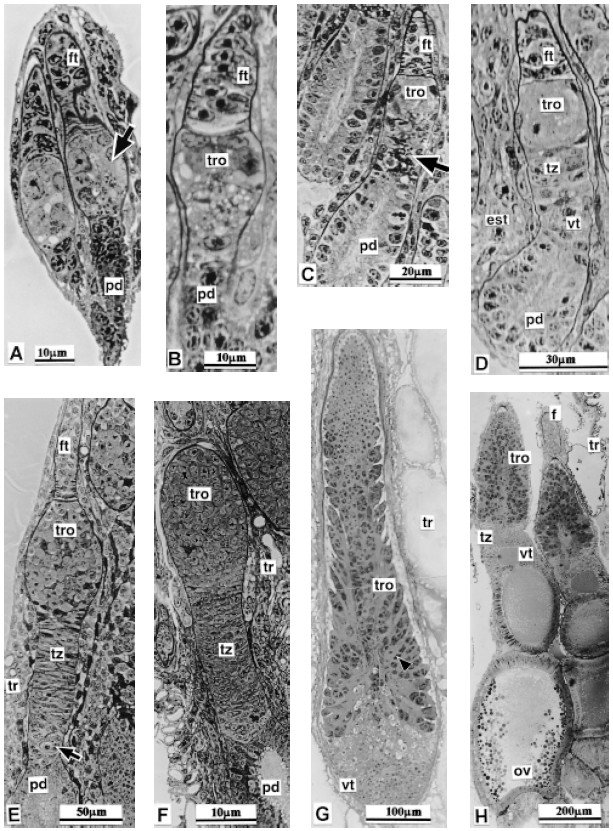
FIGURE 6. T. infestans (Klug 1834). Follow-up of the female gonad histology post embryo development as appears in 1 mm to 2 mm sections obtained after poly bed - araldite embedding. No more than longitudinal sections are presented in the eight photomicrographs. They portray ovarioles and not complete ovary. A: First instar nymph. Two ovarioles are distinguished, each one shows three regions. The central zone (arrow) of the ovariole that will give rise to the tropharium, vitelarium, nutritive chord etc. in the adult is pointed by the arrow. B: Second instar nymph. The ovariole that appear in the micrograph presents an early differentiation of tropharium. C: Third instar nymph. Three ovarioles have been sectioned at different levels. The arrow points an incipient transition region area. D: Fourth instar nymph dissected just after molted. No feeding took place after molting. Stroma tissue (est.) shows substantial increase. E: Fourth instar nymph at an advanced stage; (near to molt to the fifth instar). The arrow points the developing vitelarium site. F: Fifth instar nymph just molted. G: Fifth instar nymph at an advanced development stage. The future nutritive chords start their differentiation; (arrow). H: Adult female just molted. Two ovarioles are shown in this section. One of them presents an oocyte in initial vitelogenesis.
A specimen shortly before molting to the adult stage (when it already has a few layers of orange, non-hardened cuticle under the external cuticle, twelve to twentysix hours before ecdisis) shows ovarioles with features very similar to those of an adult mainly in relation with the tropharium cells, arranged centripetally with reference to the long axis of the ovariole. The irregular shape of their nuclei, with one or more prominent nucleoli, as well as the manifestation of structures similar to those described in adults as nutritious cells, showing also the initial branches of the developing nutritious chords.
Even though at the tropharium basis some cellular divisions appear, similar to those observed in adult ovarioles, a group of cells can be well differentiated both from the tropharium cells and those corresponding to the germarium region and/or transition zone. These cells are smaller than the nutritious cells and behave differently to staining, since their shade is lighter and their nuclei are almost spherical. Nuclei similar to these will appear later in cells that we identify in the adult as oogonia. This group of cells is neither found in egg-laying adult females nor in recently molted into 5th instar stage, at least matching the above description. On the other hand they can be found in recently molted female adults that were dissected immediately after molting with no previous food intake (Fig. 5F and Fig. 6H).
Discussion
An important part of the development of most insects takes place after the insect has hatched. This socalled post-embryonic period leads to the transformation or metamorphosis of the immature insect to assume the adult form. According to the different types of metamorphosis, the Reduvidae Hemiptera, which include the species we are studying, belong to the hemimetabolous insects (King, 1970; Richard and Davies, 1983; Wigglesworth, 1951).
Previous studies in Triatoma infestans have revealed the characteristics that determine shape and external anatomy, providing a chart of the typical 5 instar stages that make up the development of the nymphs (Canale and Carcavallo, 1985; Espínola, 1966; Gonçalves et al., 1985; Lent and Wygodzinsky, 1979). Other Reduviidae genera such as Rhodnius (Huebner and Anderson, 1972a and b) and Oncopeltus (Wick and Bonhag, 1955) have also been studied.
Previously published articles on sequential studies of the post-embryonic development of gonads are rare and obscure, particularly those referred to the morphogenesis of the ovary. However, a previous study on the formation and development of the male gonad or testicle in several Triatoma species has been published but only shows drawing, there are no photographs of the constituent parts (Gonçalves et al., 1987). No precise details are mentioned as regards the moment when the male and female gonads can be clearly differentiated. Our study clearly shows that this occurs in the 1st instar stage. As regards other Reduviidae genera, such as Oncopeltus sp. (Case, 1970) and Rhodnius (Lutz and Huebner, 1978), this aspect has not been observed; the differentiation has only been described for the last instar stage.
Our observations enable us to assert that during the 1st instar stage the male gonad is slightly bigger than the female one, a characteristic that has already been described by Mellambi (1936) in Oncopeltus, and Huebner and Anderson (1972b) in Rhodnius for later stages.
Considering the breeding conditions we can confirm that, at a constant temperature of approximately 27ºC and regular feeding, no fluctuations in the postembryonic development were observed, as was the case in other genera.
Our results make up a comparative chart of all the characteristics and/or changes the female gonad undergoes in each instar stage. Our observations correspond to microscopic examinations and photomicrographs, obtained with the above-mentioned techniques, that confirm the anatomic characteristics of the ovarioles and show how the general structure changes. Among these changes we can point out the differentiation in regions that can be observed at the histological level. Although the typical regions of the ovariole are outlined, and both filament and body of the ovariole can be distinguished since the 1st instar stage, we specifically show that the organization of the tropharium and the transition region proceed from the 3rd instar stage on, with an evident nuclear division activity in the 4th instar stage and a substantial increase in differentiation that heightens in the 5th instar stage. In the latter stage we can even observe the presence of an oocyte in initial vitelogenesis during molting to the adult stage, with no food intake. This functional behavior may correlate with the hormone activity and circulation present during these stages. (Furtado, 1976; Lutz, 1979; Stokâ, 1985; Wigglesworth, 1968). We emphasize the fact, sustained by photographic evidence, that the anatomic characteristics of the gonads are perfectly outlined when hatching occurs, and even before, during embryogenesis (manuscript under preparation). There is a similar statement for the 1st instar stage of Rhodnius, but for Triatoma´s different species it has been accepted that the morphogenesis of the ovary could be observed in the last instar stages (Stokâ and Salomon, 1985). Our results help to clarify previous data and confirm that the morphogenesis and differentiation of the female gonad in Triatoma infestans occurs in very early developmental stages.
Acknowledgements
We acknowledge to Mr. Agustin V. Chertcoff for technical assistance.
References
1. Barth R (1973). Estuds anatômicos e histológicos sobre a subfamília Triatominae (Heteroptera Reduvidae). Parte XIII. O ovario de Triatoma infestans. Mem Inst Oswaldo Cruz 71-137. [ Links ]
2. Bonhag PF (1955). Histochemical studies of the ovarian nurse tissue and oocytes of the Milkweed Bug Oncopeltus fasciatus (Dallas). J Morphol Vol 96(3): 381-421. [ Links ]
3. Canale DM, Carcavallo RV (1985). Factores Biológicos y Ecológicos En la Enfermedad de Chagas Cap. XVIII Triatoma infestans (Klug). Ed. Centro Panamericano de Ecología Humana y Salud de la OMS. pp. 237-250. [ Links ]
4. Case DC (1970). Post-embryonic development of the ovary of Rhodnius prolixus (Stahl). Master MSc Thesis Department of Zoology Mc Gill University, Montreal, Quebec, Canada. [ Links ]
5. Del Ponte E (1921). Contribución al estudio del Género Triatoma (Lap). Revista del Instituto Bacteriológico Vol. II (6): 1-64, Lam.I-XV. [ Links ]
6. Espinola HN (1966). Nota sobre diferencas sexuais em formas inmaturas de Triatominae (Hemiptera, Reduvidae). Rev Brasil Biol. Vol. 26(3): 263-267. [ Links ]
7. Furtado A (1976). Controle endocrine de l’ovogenése au cours de cinquieme stado nymphal de Panstrongylus megistus (Hemiptera Heteroptera Reduvidae). C. R. Acad. Sc. Paris, 561-564. [ Links ]
8. Galliard L (1935). Recherches sur les Reduvides Hématophages Rhodnius et Triatoma. II Abdome et evacues d l’ armature génitale des nimphes. Ann Parasit Hum Com, Vol XIII (4): 293-298. [ Links ]
9. Gillet JP (1935). The genital sterna of the immature stages of Rhodnius prolixus (Hemiptera). Trans R. Ent. Soc. Lond. Vol. 83(1): 1-5. [ Links ]
10. Gonçalves TC, Jurberg J, Costa MJ, de Souza W (1985). Estudo morfológico comparativo de ovos e ninfas de Triatoma maculata (Erichson 1848) e Triatoma pseudomaculata (Correa & Espínola 1964) (Hemiptera Reduvidea, Triatominae). Mem. Inst. Oswaldo Cruz Río J. Vol. 80(3): 263-276. [ Links ]
11. Gonçalves TC, Lent H, Ribeiro de Almeida J (1987). Estudo Anatómico e morfométrico dos folículos testiculares de algunas espécies de Triatominae (Hemiptera, Reduvidae). Mem Inst Oswaldo Cruz Rio de Janeiro Vol. 82(4): 543-550. [ Links ]
12. Huebner E, Anderson E (1972a). A cytological study of the ovary of Rhodnius prolixus. I The ontogeny of the follicular epithelium. J Morphol. 136: 459-563. [ Links ]
13. Huebner E, Anderson E (1972b). A cytological study of the ovary of Rhodnius prolixus. II Oocyte differentiation. J Morphol. 137: 385-416. [ Links ]
14. Ibañez de Barret CI, Bozzini JP, Mariano de Bozzini MI (1999). Cellular Interactions During the Female Gametogenesis of Triatoma infestans (Klug 1834). Biocell 23(2): 103-112. [ Links ]
15. King RC (1970). Ovarian Development in Drosophila melanogaster. Ed. Academic Press N. York, 227pp. [ Links ]
16. Lent H, Wygodzinsky P (1979). Revision of the Triatominae (Hemiptera Reduvidae) and their significance as vectors of Chagas’s disease. Bull Am Mus Nat Hist. 163: 127-520. [ Links ]
17. Lutz D, Huebner E (1978). Structural and physiological aspects of post-embryonic ovarian development in Rhodnius prolixus. J Cell Biol. 79 (2 part 2): G1026 (abstract). [ Links ]
18. Lutz DA (1979). Structural and physiological aspects of 5th instar ovarian development in Rhodnius prolixus (Insecta, Hemiptera). M Sch Thesis. Dept. Zoll. Univ. Manitoba. Canada. [ Links ]
19. Mellanby H (1936). The later embryology of Rhodnius prolixus. Q J Micros Sci. 79: 1-42. [ Links ]
20. Mollenhauer HH (1964). Plastic embedding mixtures for use in electron microscopy. Stain Technology 39: 111. [ Links ]
21. Richard OW, Davies RG (1983). El Sistema Reproductor. Cap. 17. In: Tratado de Entomología. Ed. Omega Madrid, Vol. 1: 305-336. [ Links ]
22. Stokâ AM, Salomón OD (1985). Desarrollo y maduración de los ovocitos en Triatominæ. Cap IX. In : Factores Biológicos y Ecológicos en la Enfermedad de Chagas Vol. I. Epidemiología y Vectores. Ed. Carcavallo R., Rabinovich J L, John B. [ Links ]
23. Stokâ AM (1985). Mecanismos endocrinos en la regulación del desarrollo de triatominos. Cap X: 103 - 111 in: Factores biológicos y ecológicos en la enfermedad de Chagas. Centro Panam. de Ecol. Humana y Salud de la OMS. Editores Canals D.A., Carcavalo R. [ Links ]
24. Wick JP, Bonhag PF (1955). Postembrionic development of the ovaries of Oncopeltus fasciatus (Dallas). J Morphol. 96: 31-66. [ Links ]
25. Wigglesworth UB (1951). Metamorphosis in Insects. Review. Proc. Roy. Ent. Soc. London Ser. C Vol. 15: 78-81. [ Links ]
26. Wigglesworth UB (1968). Summing up Growth Hormons and the Gene System in the Insect Rhodnius. Mem Soc Endoc. 15: 77-85. [ Links ]
Received on June 13, 2003
Accepted on April 27, 2004.


{kind=link}