










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBiocell
versión impresa ISSN 0327-9545
Biocell v.29 n.2 Mendoza abr./ago. 2005
DNA injury induced by 5-aminouracil and caffeine in G2 checkpoints path of higher plant cells
A. Del Campo*, M. Bracho, L. Marcano, J. Guíñez, and C. De la Torre**
* Universidad del Zulia. Facultad Experimental de Ciencias. Departamento de Biología. Maracaibo. Estado Zulia. Venezuela.
** Consejo Superior de Investigaciones Científicas. Centro de Investigaciones Biológicas. Departamento de Biología Celular y del Desarrollo. Madrid, España.
Address correspondence to: Dr. Antonio Del Campo. Departamento de Biología. Facultad Experimental de Ciencias. Universidad del Zulia. Maracaibo, Estado Zulia. VENEZUELA. Telefax: (+58-261) 7598109. E-mail: delcampoa@terra.com
Abstract: This work evaluated the qualitative and quantitative cellular changes induced by treatment with 5-aminouracil (5-AU) and a combination of 5-AU and caffeine in plant cells in relation to DNA damage, repaired damage, and residual damage. As biological material, Allium cepa L. root tips were used, grown in filtered water, in darkness, with aeration at constant temperature of 25 °C ± 0.5. Cell populations were synchronized using 5 mM caffeine in order to study the effects of 5-AU and caffeine/5-AU combined treatment on the DNA content and their incidence in the entrance to mitosis. The results showed a delay in the G2 period due to induced DNA damage by the 5-AU and caffeine/5-AU combined treatment, shown by aberrant metaphases, anaphases and telophases. The effect of caffeine in the combined treatment was heightened in spite of lengthening the checkpoints route that retains the cells in G2. The existence of G2 checkpoints was shown in the cell population studied, inducing lesions in the DNA, chromosomic aberrations and cellular instability.
Keywords: 5-aminouracil. G2 checkpoints. Caffeine.
Introduction
Proteins responsible for the regulation of the cellular division cycle are under strict control of systems that trigger or block the machinery of the cycle in the correct order and once per cycle, responding to intra - and extra- cellular information which assure that the cycle is perfectly developed (Forrest et al., 2001; McGowan, 2002; Park and Lee, 2003).
When these control systems receive an inhibitory sign as the one caused by an incomplete process, the cycle is blocked by some elements of transition called negative regulators or checkpoints (Rao and Johnson, 1974; Nurse, 1994). There are three important checkpoints during the cell cycle: the first one located in G1/S, where the entrance to replication is blocked because of unfavorable environmental conditions or inappropriate cellular growth; the second one located in the entrance to mitosis, (G2/M), where the cycle is blocked if the replication of the DNA has not been completed or there is damaged DNA; and finally the third checkpoint (M/G1) activated during the change of metaphase to anaphase, when the chromosomes have not joined to the microtubules of the mitotic spindle or it has not been formed correctly (Harwell and Weinert, 1989; Murray, 1992).
The regulation of the cell cycle depends on the balance of positive stimuli that lead on and negative ones that stop it in the three points described previously. The existence of these checkpoints during the cell cycle assumes that the cells are able to recognize the incidence of DNA damage, impeding the onset of replication or mitosis by means of sign inhibition of Cdks/cyclins (Malumbres et al., 2000). At present, the positive mechanisms of regulation are better known than the negative ones; however, their implications in the regulation in normal and altered cells are similar in both cases (Park and Lee, 2003).
There are several reports that establish the effect of metabolic inhibitors on these cycle regulators that act by blocking either the positive or the negative stimuli (Baud and Karin, 2001; Bradley and Pober, 2001; Malumbres et al., 2000). Specifically in the G2 period, considered as a crucial point for the cycle progression (Forrest et al., 2003), the effect of different drugs has been established in animal cells and, to a smaller degree, plant cells (López-Sáez et al., 1966; Tobey, 1975; Murakami and Nurse, 2000; Nigg, 2001).
This work first determines the relationship between damage, duration of the stopped G2 phase, and DNA repair capability, measured as the difference between the produced and the remainder cell damage detected in mitosis. Secondly, the effect of caffeine behavior as preferential inhibitor of DNA repair that takes place during the blocked G2 (Del Campo et al., 1997), by two preferential mechanisms: prevention of damage detection and slowness of the reparative biosynthesis pathways.
Materials and Methods
Biological materials and study conditions
Allium cepa L. bulbs were placed in recipients with filtered water and changed every 24 h. Roots grew at a constant temperature of 25ºC ± 0.5, in darkness and with constant aeration (10-20 ml/min). Root tips were used when they reached the dynamic balance of growth and proliferation, when all the parameters of the cell cycle remained constant.
Treatments
Roots were treated for 2 h with 5 mM caffeine (Merck), in order to inhibit cytokinesis since the caffeine impedes the coalition of the Golgi vesicles which are responsible for the formation of cellular wall, originating binucleated cell populations (López-Sáez et al., 1966), (Fig. 1). When the roots were 2-3 cm long, they were submerged, along with the bulbs, in the different treatment solutions, prepared with filtered water and without altering the environmental conditions set for the development of the root tips.

FIGURE 1. Bimetaphase cell after caffeine treatment. X 400
Determination of 5AU optimal concentration
Different 5-AU concentrations (10-3 M, 5 x 10-4 M and 10-4 M) were used. The 5-AU effect was valuated by the mitotic index alteration (MI) as a parameter of root growth rate.
5-AU Effect on the G2 period progression
5-AU optimal concentration (5 x 10-4 M) was used to treat the binucleated population for 3 h during the period that corresponds to the end of the DNA replication (from the 7th hour to the 10th hour after S phase initiation, Fig. 2).

FIGURE 2. Experimental protocol to evaluate the 5-AU effect and the double treatment 5AU/ caffeine effect to binucleated A. cepa cells.
Influence of caffeine treatment on the 5-AU effects
A secondary, 5mM caffeine treatment was given for 2 h (10th -12th hour) in order to determine caffeine effect on the produced genome damage in relation with controls grown under the same conditions of temperature, darkness and constant aeration (Fig. 2).
Cell recovery capability to drugs effect
To determine cells recovery capacity, meristems were placed in filtered water after each treatment, under the same growing conditions for 25 and 40 h. During this time cells developed 1 or 2 complete cell cycles. Recuperation was evaluated by mitotic index (MI), percentage of chromosomic aberrations and duration of G2 phase.
Nuclei in replication detection
To determine the duration of the S period, roots were separated from the bulb and incubated for 10 min with H3-thymidine (Amersham, UK, 370 KBq.ml-1, 925 GBq.mmol-1 specific activities) at 2, 4, 6, 8, 10 and 12 h. The slides with cell preparation were covered with radiographic emulsion NTB-2 (Kodak) diluted 1:1 (v/ v) in water and incubated for 9 days at -70ºC. Slides were treated with developing Kodak D-19 solution and fixed with Kodak ultra fast fixer.
Cytological techniques
Roots were separated from the bulb and fixed in a mixture of glacial acetic acid - ethanol (3:1 v/v). Afterward, the meristematic tissues were dehydrated in serial alcohol concentrations (50º, 75º, 95º and 100º), stained by the Feulgen technique and extended in monolayer on glass slides.
Statistical analysis
Student's test was used to establish comparisons between averages of the treated cells and the controls with a significance level less than 0.05.
Results
Optimal concentration of 5-AU
5-AU at 5x10-4 M was selected as optimal concentration since 5x10-3 M 5-AU considerably depresses cell growth. Although 5x10-4 M 5-AU depresses growth, new proliferate balances are acquired that come close to the control values and remain stable during the treatment time (MI = 8.4%, Fig. 3).
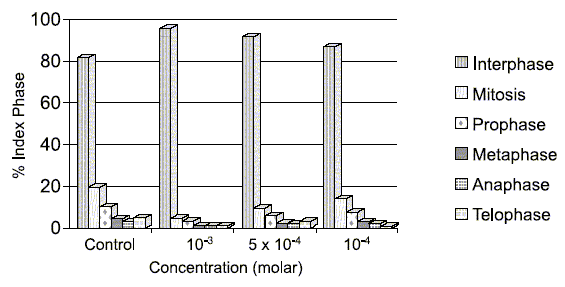
FIGURE 3. Effect of the 5-AU on index phase (IP) of cell cycle in Allium cepa meristems.
Effect of the 5-AU on the progression of the G2
Results showed a G2 duration of 4 h, with a delay of 1.8 h for the beginning of prophase in comparison with the controls (Table 1). This 1.8 h delay indicates operability of the G2 checkpoint which is the controller of the synchronous entrance to prometaphase (Del Campo et al., 1997). Figure 4 a and b shows that after this delay, some nuclei enter mitosis earlier. This effect is shown as heterophasic bimitosis, indicating that the negative G2 control was weakened during the 5-AU treatment.
TABLE 1.
Effect of the 5-AU and combined 5-AU/caffeine treatment on the development of G2., delay mitotic and induction of chromosomic aberrations

FIGURE 4. Chromosomal anomalies (ChA) induced by 5-AU/caffeine in tips roots onion (Allium cepa L). X 400 a. Binucleated cell with micronuclei (arrows). b. Cells with agglutinated chromosomes (arrows). c. Cells with chromosomal segregated chromosomes (arrows). d. Bitelophasic cell with chromosomal bridge (arrow).
These results indicate the presence of a negative checkpoint control responsible for the synchronous entrance to prometaphase and for the detection of the relationship between the produced damage and the one repaired, valued as the percentage of chromosomic aberrations detected at 25 and 40 h. The difference between the mean of these percentages and the percentage of chromosomic aberrations at the end of the treatment gives the residual (or not repaired) damage (Table 1).
Effect of combined 5-AU/caffeine treatment on the induction of chromosomic aberrations and development of G2
A significant induction (P < 0.01) of chromosomic aberrations in exposed cells is detected (Table 2), and the effect was amplified with the caffeine treatment. In spite of the mitotic delay induced by exposure of 5-AU being given, a considerably increased of abnormal metaphase, anaphase and some telophase were observed (Table 1). Therefore, the DNA repair pathway is not completely showed as an increase in the apparition of cells with micronuclei and nuclei with agglutinated chromosomes (Table 2; Fig. 4 a, b, c and d) and to a less extent, cells with chromosomal break, chromosomal bridge and segregated chromosomes.
TABLE 2.
% of chromosomal aberrations at different concentrations of 5AU and 5AU + caffeine

The effect of this combined treatment is evidenced by the loss of the nuclei synchronicity when entering prometaphase (Fig. 5). After a delay, in comparison to the controls, of 1.8 h in the case of the 5-AU and of 4.1 h for the combined treatment, some nuclei start a premature mitosis, observed as heterophasic bimitosis, indicating checkpoint G2 alterations (Del Campo et al., 2003).

FIGURE 5. Loss of the nuclei synchronicity and bimitosis heterophasic are present. a. Binucleated cell, with nuclei in prophase (p) and one in metaphase (m). X 400 b. Binucleated cell, with nuclei in pro-anaphase (a) and one in metaphase (m). X 400
According to the results obtained for 5-AU/caffeine treatment, doubling the effect of the single 5-AU treatment could be explained as the existence of 2 cooperative routes: increase of the mitotic delay and inhibition of some DNA repair routes.
With the purpose of establishing the additive effect of the chromosomal damage exercised by caffeine, the potential factor (Fp) was determined increasing severely the frequency of chromosomic aberrations induced by 5-AU. The Fp expresses the relationship between the combined treatment, 5-AU/cafeína, Σ(i + p) and the sum of the inducing agent, Σ i = 5AU and the additing agent, Σ p = caffeine.
Then:
Fp = Σ (i + p) / Σ i + Σ p
For the case of the Table 3, it is:
Fp = Σ (i + p) / Σ i = 0.39 / 0.22 = 1.7
TABLE 3.
Additive effect of the chromosomal damage exercised by caffeine evaluated by the determination of the potencial factor.

Fp expresses the magnitude of the increased chromosomic aberrations induced by the 5-AU and caffeine, practically duplicating the frequency of abnormal metaphase and anaphase, confirming that caffeine is able to produce chemical damage to cells in G2 (Table 3).
Discussion
The eukaryotic cells in proliferation possess a safety mechanism that interrupts their passage from G2 to mitosis if the DNA has not been totally replicated or repaired (Murray, 1992; Park and Lee, 2003). This negative control is blocked when DNA base analogs are used which can cause cell pass to mitosis with alterations in its genetic material (Lydall and Weinert, 1995; Nigg, 2001; Robinson et al., 2001). The use of these analogs, 5AU and caffeine, disable these safety mechanisms either in animal (Malumbres et al., 2000) or plant cells (Murakami and Nurse, 2000). The existence of these controls during the cellular cycle assumes that the cells can recognize damage in its DNA and impede the beginning of replication or mitosis.
Genetic experiments in exposed bacteria to UV radiations (Selby and Sancar, 1990), and biochemical studies on DNA replication and repair (Tobey, 1975; Forrest et al., 2003), have given detailed information on the events. However, the studies in eukaryotic cells have not been very successful due to the higher cell complexity and organization. The existence of a chromatin with a great structural and functional versatility determines that a genetic lesion might cause an intricate net of molecular alterations.
With the purpose of studying the recovery capacity of cells in G2 as well as the passage for this phase of the cycle, two aberrant agents have been used: 5-AU and caffeine. These two clastogenic compounds, inaddition to producing chromosomic aberrations, cause a delay in the kinetics to mitosis in injured cells.
The results allow for establishment that the 5-AU has two related effects: 1) it inhibits the late replication responsible for the premetaphasic synchronization which agrees with other reports (Del Campo et al., 1997) and 2) it promotes delay in the post-replicative phase due to induced genomic lesions with duration in G2 of 4 h in comparison with 2.2 h observed in controls. These values are triplicate when a secondary treatment was given with 5 mM caffeine (Table 1). The results allow to infer that the cells tried with the 5-AU presented a damage that makes them sensitive to the caffeine treatment and that an important fraction of the lesions produced by the 5-AU is repaired when the cells are in the last phase of G2. The caffeine inhibits the repair mechanisms in G2 (Del Campo et al., 2003). In presence of this drug the cells arrive to mitosis with the types of unrepaired lesions before mentioned (Table 2).
As it is observed in Figure 4a, one of the nuclei suffered a premature chromosomal condensation, observed in a state similar to prometaphase, with its nuclear membrane seemingly broken. The chromosomal breaks that appear correspond to segments, replicated or not, that could eventually originate micronucleus (Marcano et al., 1999). Observation of these aneuploid nuclei and other chromosomic aberrations like breaks, anaphasic bridges, agglutinated chromosomes and left-behind chromosomes, implies the effect of the treatments on the spindle microtubules, reflected in segregation problems, as has been observed in yeasts (Lydall and Weinert, 1995). The micronucleus observation in different phases of mitosis (Fig. 5), indicates the presence of nucleolus organizing, that are able to capture the diffusible factor which trigger the entrance to replication and once the nuclei are repaired, the entrance to mitosis. Similar results have been reported in animal and plant cells (Gonzalez-Fernández et al., 1971; Rao and Johnson, 1974; Hervás et al., 1982; Del Campo, 2003).
Two synergetic actions are set out during the DNA repair mechanisms: mitotic retardation associated to genomic damage and removal of the lesions with recovery of the DNA normal structure, or on the contrary, programmed cellular death when recovery is not possible.
The results of the combined 5-AU/caffeine treatment, establish that the action of caffeine could reside in the existence of 2 cooperative effects: increase of the mitotic delay and inhibition of some of the repair routes. The results shown in table 1 evidence a repaired damage average of 9.2% for the case of the 5-AU treatment and of 11.35% for the double 5AU/caffeine treatment, with a residual damage of 12.8 for the first case and of 27.8 for the second one. This is why it is presumed that the damage caused by the drug is irreversible starting from the second cellular cycle. Either the 5-AU or the combined action of this one with the caffeine, alters the controller capacity of the negative regulators G2 described in different cell systems (Tobey, 1975; Murakami and Nurse, 2000; Nigg, 2001; Del Campo et al., 2003).
Conclusions
5-AU has 2 related effects: inhibition of late DNA replication, responsible for the premetaphasic synchronization and induction of post-replicate delay with the purpose of repairing damage to the genome caused by the drug treatment.
Caffeine increased the damage caused by the 5-AU treatment, duplicating the number of aberrant anomalies, and in consequence, the duration of G2, inhibiting some DNA repair routes.
The existence of control mechanisms is evidenced by the analyzed biological pattern, similar to those described in other organisms.
Acknowledgments
This work was financed by the Fondo Nacional para la Ciencia y la Tecnología de Venezuela (FONACIT). Special thanks to Bradley W. Christian for correction on English grammar.
References
1. Baud V, Karin M (2001). Signal transduction by tumor necrosis factor and its relatives. Trends Cell Biol. 11: 372-377. [ Links ]
2. Bradley JR, Pober JS. (2001). Tumor necrosis factorreceptor-associated factors (TRAFs). Oncogene, 20: 6482-6491. [ Links ]
3. Del Campo A, et al (2003). G2 checkpoint targets late replicating DNA. Biology of the Cell. 95: 521-526. [ Links ]
4. Del Campo A, et al. (1997). Frailty of two cell cycle checkpoins which prevent entry into mitosis and progression through early mitotic stages in higher plant cells. Eur J Cell Biol. 74: 289-293. [ Links ]
5. Forrest AN, Taylor D, Grimmond S (2003). Exploration of the cell cycle genes fond within the RIKEN FANTOM2 data set. Genome Res. 13(6B): 1366-1375. [ Links ]
6. González-Fernández A, et al. (1971). Interphase development and biginning of mitosis in the different nuclei of polimerase homocaryotic cells. Chromosoma, 36: 101-112. [ Links ]
7. Harwell LH, Weinert TA (1989). Checkpoint controls that ensure the order of cell cycle events. Science, 246: 629-634. [ Links ]
8. Hervás JP, López-Sáez JF, Giménez-Martín G (1982). Multinucleate plant cells. II. Requirements for nuclear DNA synthesis. Exptl Cell Res. 139: 341-350. [ Links ]
9. López-Sáez JF, Risueño MC, Giménez-Martín G (1966). Inhibition of cytokinesis in plant cells. J ultrastruct Res. 14: 85-92. [ Links ]
10. Lydall D, Weinert TA (1995). Yeast checkpoint genes in DNAdamage processing implications for repair and arrest. Science, 270: 1488-1491. [ Links ]
11. Malumbres M, Ortega S, Barbacid M (2000). Genetic analysis of mammalian cyclin-dependent kinases and their inhibitors. Biol Chem. 381: 827-838. [ Links ]
12. Marcano de ML, Carruyo I, Montiel X, Bracho M, Soto LM (1999). Valoración del efecto tóxico del cadmio en células meristemáticas de cebolla Allium cepa, L. Rev Fac Agron, 16: 476-487. [ Links ]
13. McGowan CH (2002). Checking in on Cdsl (Chk2): A checkpoint kinase an tumor suppresor. Bioessays, 24: 502-511. [ Links ]
14. Murakami H, Nurse P (2000). DNA replication and damage checkpoints and meiotic cell cycle controls in the fiffion and budding yeast. Biochem J, 349: 1-12. [ Links ]
15. Murray AW (1992). Creative blocks cell-cycle checkpoints and feetback controls. Nature, 359: 599-604. [ Links ]
16. Nigg EA (2001). Mitotic kinases as regulators of cell division and its checkpoints. Nat Rev Mol Cell Biol, 2: 21-32. [ Links ]
17. Nurse P (1994). Ordering S phase and M phase in the cell cycle. Cell, 79: 547-550. [ Links ]
18. Park MT, Lee SJ (2003). Cell cycle and cancer. J Biochem Mol Biol, 36: 60-65. [ Links ]
19. Rao PN, Johnson RT (1974). Regulation of cell cycle in hybrid cells. In: Cell spring Harbor Lab (ed). Control of proliferation in animal cells. p. 785. [ Links ]
20. Robinson IB, Broude EV, Chang BD (2001). If not apoptosis, then what? Treatment-induced senescence and mitotic catastrophe in tumor cells. Drug Resist Updat, 4: 303-313. [ Links ]
21. Selby CP, Sancar A (1990). Mechanisms of caffeine inhibition of DNA repair in E. coli. Prog Clin Biol Res, 340: 179-193. [ Links ]
22. Tobey RA (1975). Different drugs arrested cells at a number of distinct stages in G2. Nature, 254: 245-247. [ Links ]
Received on April 22, 2004.
Accepted on March 16, 2005.

