










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBiocell
Print version ISSN 0327-9545
Biocell vol.34 no.3 Mendoza Sept./Dec. 2010
ORIGINAL ARTICLES
Influence of estradiol analogue on cell growth, morphology and death in esophageal carcinoma cells
Thandi Mqoco1, Sumari Marais1 and Annie Joubert
Department of Physiology, University of Pretoria, Pretoria, South Africa
*Address correspondence to: Annie Joubert. E-mail: annie.joubert@up.ac.za
ABSTRACT: 2-Methoxyestradiol-bis-sulphamate is a bis-sulphamoylated derivative of the naturally occurring 17-beta-estradiol metabolite namely 2-methoxyestradiol. 2-Methoxyestradiol-bis-sulphamate is regarded as a potential anticancer drug with increased antiproliferative activity when compared to 2-methoxyestradiol. The aim of this pilot in vitro study was to determine the influence of 2-methoxyestradiol-bis-sulphamate on cell growth, morphology and possible induction of certain types of cell death in the SNO esophageal carcinoma cell line. A dose-dependent study (0.2-1.0µM) was conducted with an exposure time of 24 hours. Data revealed that 2-methoxyestradiol-bis-sulphamate reduced cell numbers statistically significantly to 74% after exposure to 0.4µM of the drug. Morphological studies including light microscopy demonstrated hallmarks of apoptosis, while fluorescent microscopy revealed both the presence of apoptosis and autophagy as types of cell death being induced in SNO cells after 24 hours of exposure to 0.4µM 2-methoxyestradiol-bis-sulphamate.
Key words: Cancer; Apoptosis; Autophagy
Introduction
Esophageal squamous cell carcinoma is one of the most frequently occurring cancers in South Africa with the highest incidence observed in South African black males (Davies-Coleman et al., 2005). Due to limited efficacy that current chemotherapeutic agents offer, new drugs with increased anticancer potential are being researched. 2-Methoxyestradiol is a naturally occurring 17-b-estradiol metabolite which is currently undergoing phase II human clinical trials for the treatment of breast- and prostate cancer (Fotsis et al., 1994; Lakhani et al., 2003; Thaver et al., 2009). 2-Methoxyestradiol exerts anti-tumour and anti-angiogenic properties in vitro and in vivo (Dingli et al., 2002; Lippert et al., 2000; Thaver et al., 2009; Schumacher and Neuhaus, 2001; Cushman et al., 1995). However, 2-methoxyestradiol is rapidly inactivated by 17b-hydroxysteroid dehydrogenase type 2, an enzyme that is mainly expressed in both the gastrointestinal tract and liver, leading to 2-methoxyestradiol exerting a low bioavailability (Foster et al., 2008; Utsumi et al., 2005; Newman et al., 2006; Ireson et al., 2004; Sano et al., 2001). Therefore, an analogue of 2-methoxyestradiol, namely 2-methoxyestradiol-bis-sulphamate (2-MeOE2bisMATE) is currently being researched as a potential anticancer drug. 2-MeOE2bisMATE is a potent inhibitor of steroid sulphatase, an enzyme that converts estrone sulphate (E1S) to estrone. Estrone can be converted to a biologically active estrogen, namely estradiol. The latter binds to estrogen receptors (ER) and stimulates growth of hormone-dependent tumours (Purohit et al., 2001; Purohit et al., 2003).
Recent in vitro studies from our laboratory and other researchers have found that 2-MeOE2bisMATE inhibits cell growth in cancer cell lines namely the human breast adenocarcinoma ER+ cell line (MCF-7), human breast adenocarcinoma ER- cell line (MDA-MB-231), human umbilical vein endothelial cells (HUVEC), human prostate cancer cell line (PC3) and ovarian cancer cells (Foster et al., 2008; Purohit et al., 2003; Day et al., 2003; Raobaikady et al., 2003; Ireson et al., 2004; Tagg et al., 2008; Suzuki et al., 2003). Some cell types were found to be more sensitive to 2-MeOE2bisMATE, suggesting differential signal transduction exerted by this compound. It has been postulated that 2-MeOE2bisMATE causes microtubule disruption that culminates in apoptosis (Purohit et al., 2003; Day et al., 2003; Tagg et al., 2008). The aim of this novel in vitro study was to determine the influence of 2-MeOE2bisMATE on growth, morphology and its possible induction of two types of cell death namely apoptosis and autophagy in SNO esophageal carcinoma cells that has never been reported previously.
Materials and Methods
Materials
The SNO esophageal carcinoma cell line was purchased from Highveld Biological Ltd. (Pty) (Sandringham, SA). These cells are described as non-keratinizing squamous epithelial cells. Penicillin, streptomycin, fungizone and trypsin were obtained from Highveld Biological (Pty) Ltd. (Sandringham, SA). Dimethyl sulphoxide (DMSO), propidium iodide, Hoechst 3342 (HO) and acridine orange (AO) were supplied by Sigma-Aldrich Co. (St Louis, USA). Haematoxylin, eosin, ethanol and xylol fixative were purchased from Merck (Darmstadt, Germany). Dulbecco's Modified Eagle's Medium with glucose, sodium pyruvate and L-glutamine (DMEM), fetal calf serum (FCS), sterile cell culture flasks and plates were obtained from Separations (Randburg, Johannesburg, SA). Since 2-MeOE2bisMATE is currently not commercially available, the compound was synthesized by Prof. Vleggaar from the Department of Chemistry (University of Pretoria, Pretoria, SA).
Cell culture
To conduct cell growth studies, cells were propagated as monolayers in growth medium at 37ºC in a humidified atmosphere containing 5% CO2. A stock solution of 2-MeOE2bisMATE with a concentration of 2.0x10-3M was prepared in DMSO and stored at -20ºC. Appropriated controls were included for each experiment. One of the controls contained cells propagated in growth medium; while the other control was defined as the vehicle-treated control where cells were treated with the same volume of DMSO used to expose cells with 2-MeOE2bisMATE. The DMSO content of the final dilutions never exceeded 0.05% (v/v). Positive controls for the experiments included cells which were exposed to actinomycin D (0.1µg/ml) and starved cells (cells which were propagated in a solution which contained both growth medium and PBS at a ratio of 1:2 respectively). Dose-dependent studies (0.2-1µM) were conducted with analysis of the effects of 2-MeOE2bisMATE on cell growth in SNO cells after 24 hours of exposure. This concentration range was chosen since previous studies have shown that it is within this range that 2-MeOE2bisMATE is most efficient as an antiproliferative agent (Raobaikady et al., 2003).
Determination of cell numbers (crystal violet staining)
Exponentially growing SNO cells were seeded in 96-well tissue culture plates at a cell density of 5 000 cells per well. Cells were incubated at 37ºC for a period of 24 hours to allow for attachment. Medium was discarded and cells were exposed to a dilution series ranging from 0.2-1µM 2-MeOE2bisMATE, including controls as described previously. The experiment was terminated by the addition of 100µl of 1% glutaraldehyde in water for 15 minutes. Crystal violet (100µl) (1%, in PBS) was added for 30 minutes, after which the culture wells were immersed in running tap water for 15 minutes. The plates were left overnight to dry. Then 200µl of Triton X-100 (0.2% in water) was added to each well. Plates were incubated for 30 minutes and 100µl of the liquid content was transferred to 96-well plates. The absorbance of the samples was analysed using an ELX800 Universal Microplate Reader (Bio- Tek Instruments, Inc., Analytical Diagnostic Products, Weltevreden, SA).
Polarization-optical differential interference contrast
Exponentially growing SNO cells were seeded at 500 000 cells per well in a 6-well plates. After a 24 hour incubation period at 37ºC to allow for cell adherence, cells were exposed to 0.4 µM of 2-MeOE2bisMATE. A concentration of 0.4 µM of 2-MeOE2bisMATE was chosen as focus dosage, since this concentration demonstrated pronounced inhibition of cell growth. Appropriate controls were included as described previously. Polarization-optical differential interference contrast images were obtained using the Zeiss Axiovert-40 microscope (Göttingen, Germany) after 24 hours of exposure to 2-MeOE2bisMATE and to the controls respectively.
Cell morphology (haematoxylin and eosin staining)
For light microscopy, exponentially growing SNO cells were seeded at 250 000 cells per well in 6-well plates on heat-sterilized coverslips. After a 24 hours incubation period at 37ºC, cells were exposed to 0.4 µM of 2-MeOE2bisMATE for 24 hours at 37ºC. Coverslips were removed and transferred to a staining dish. Cells were fixed in Bouin's fixative for 30 minutes. The fixative was discarded and 70% ethanol was added to the coverslips (20 minutes), rinsed in tap water and subsequently left for 20 minutes in Mayer's hemalum. After rinsing with running tap water for 2 minutes, coverslips were washed with 70% ethanol before being subjected to 1% eosin for 2 minutes. This was followed by rinsing twice for 5 minutes with 70% ethanol, 96% ethanol, 100% ethanol and xylol respectively. Coverslips were mounted with resin and left to dry before they were evaluated with a Zeiss Axio reflected light microscope. Mitotic indices were obtained by counting 1000 cells on each slide of the biological replicates and expressing it as a percentage of cells in mitosis and apoptosis.
Cell morphology (triple staining technique)
For fluorescent microscopy, SNO cells were seeded according to the same procedure describe for light microscopy. Cells which were exposed to actinomycin-D and starved cells were included as positive controls. The amount of 0.5ml HO solution (3.5µg/ml in PBS) was added to the medium to give a final concentration of 0.9µM and cells were incubated for a period of 30 minutes at 37ºC. After 25 minutes into incubation, 0.5 ml of AO solution (4 µg/ml in PBS) was added to the medium to give a final concentration of 1 µg/ml and 0.5 ml of PI solution (40 µg/ml in PBS) was also added to the medium to give a final concentration of 12 µM and incubated for another 5 minutes at 37ºC. Medium was discarded and cells were rinsed twice with PBS. Samples were examined with a Zeiss inverted Axiovert CFL40 microscope and Zeiss Axiovert MRm monochrome camera using Zeiss Filter 2 for Hoechst 33342-stained cells (blue emission), Zeiss Filter 9 for acridine orange-stained cells (green emission) and Zeiss filter 15 for propidium iodide-stained cells (red emission). In order to prevent fluorescent dye quenching; all procedures were performed with plates and reagents covered with foil.
Statistical analysis of data
The ANOVA Students't-test was used to determine the analytical variation in experimental procedures and biological variations within each experiment. Cell growth studies were repeated three times with a sample size of 6 in each experiment. A P-value of <0.05 was regarded as statistically significant and indicated with an asterisk (*) on the graph. Fluorescent microscopy and Polarization-optical differential interference contrast provided qualitative data, whereas the haematoxylin and eosin staining procedure provided both qualitative and quantitative (mitotic indices) results.
Results
Determination of cell numbers
The effects of SNO cells exposed to 0.2-1µM 2-MeOE2bisMATE for 24 hours were evaluated and compared to appropriate controls. Cells treated with vehicle (DMSO), as well as those treated with concentrations lower than 0.4µM 2-MeOE2bisMATE did not show a significant decline in cell growth (Fig. 1). A statistically significant decrease in cell growth (74%) was observed at a concentration of 0.4µM 2-MeOE2bisMATE (P-value <0.05). Cells exposed to 0.5 µM and 0.6µM 2-MeOE2bisMATE respectively revealed a slight decrease in cell growth, while those cells exposed to 0.7-1µM 2-MeOE2bisMATE decreased cell growth to approximately the same extend when compared to the effect exerted by 0.4µM 2-MeOE2bisMATE (Fig. 1).
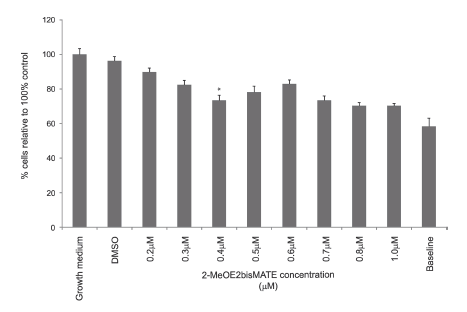
FIGURE 1. SNO cell numbers expressed as a percent of cells relative to the control (cells propagated in growth medium only) after being exposed to 2-MeOE2bisMATE (0.2-1μM) for 24 hours. A statistically significant inhibition of cell growth was observed after a 24 hours exposure period to 0.4μM 2-MeOE2bisMATE (74%). ANOVA Students't-test was used to determine the analytical variation in experimental procedures and biological variations within each experiment to determine the P-value. An asterisk indicates a P-value < 0.05.
Polarization-optical differential interference contrast
This was used to visualise the effect of 0.4µM 2-MeOE2bisMATE in SNO cells. SNO-treated cells showed an increase in the number of cells in mitosis (Fig. 2C), indicating that the cells are blocked in mitosis. The cell density of treated-SNO cells was compromised when compared to the density of the controls (Fig. 2A-D).

FIGURE 2. Polarization-optical differential interference contrast photos of SNO cells propagated in growth medium (A), vehicle-treated control (B), 0.4μM 2-MeOE2bisMATE-treated cells (C) and actinomycin-D treated cells (D) after an exposure period of 24 hours. SNO cells treated with 2-MeOE2bisMATE revealed cells blocked in metaphase (round cells) while control-treated cells were in normal phases of mitosis (scale bar indicates 30μm).
Cell morphology (haematoxylin and eosin staining)
A haematoxylin and eosin cell staining technique was performed in order to reveal the morphological changes of the nuclear and cytoplasmic components of the SNO cells after 24 hours of exposure to 0.4 µM 2-MeOE2bisMATE (Fig. 3C). The SNO control showed interphase cells and normal cell division including a cell in telophase after 24 hours of exposure (Fig. 3A and B). Cells treated with 2-MeOE2bisMATE revealed apoptotic features namely hypercondensed chromatin, membrane blebbing and apoptotic bodies. Several cells were also blocked in metaphase (Fig. 3C). Actinomycin-D treated cells (Fig. 3D) served as a positive control for the induction of apoptosis.
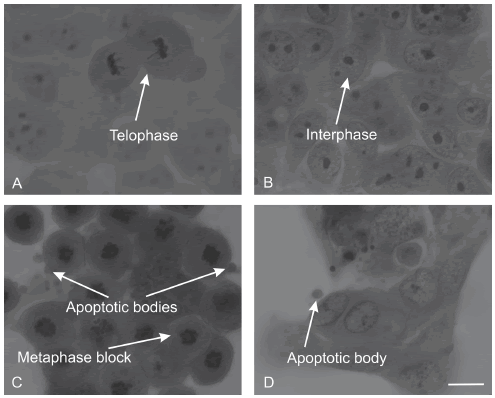
FIGURE 3. Haematoxylin and eosin staining of SNO cells propagated in growth medium (A), vehicle-treated control (B), 0.4μM 2-MeOE2bisMATE-treated cells (C) and actinomycin-D treated cells (D) after 24 hours of exposure. 2-MeOE2bisMATE-treated-SNO cells revealed cells blocked in metaphase, hypercondensed chromatin and the presence of apoptotic bodies (magnification 100x, scale bar indicates 30μm)
Cell morphology (triple staining technique)
Triple staining was used to determine possible induction of apoptosis and autophagy after 24 hours of exposure to 0.4µM 2-MeOE2bisMATE. Acridine orange stains lysosomes and vacuoles of cells undergoing autophagy with an increase in green fluorescence. Hoechst stains the nuclei of viable and apoptotic cells with a blue fluorescence. Propidium iodide stains cells with compromised membrane integrity (oncotic/necrotic processes) with a red fluorescence. Results showed an increased tendency for acridine orange in both 2-MeOE2bisMATE-treated SNO cells and starved cells (Fig. 4C and E) when compared to the controls (Fig. 4A and B). Cell membranes were not disrupted, since no red fluorescence (fluorescence emitted by propidium iodide) was observed in any of the cells. Results revealed that 2-MeOE2bisMATE caused cell death by apoptosis and autophagy, since an intense blue and green fluorescence (indicating increased lysosomal activity) were observed.
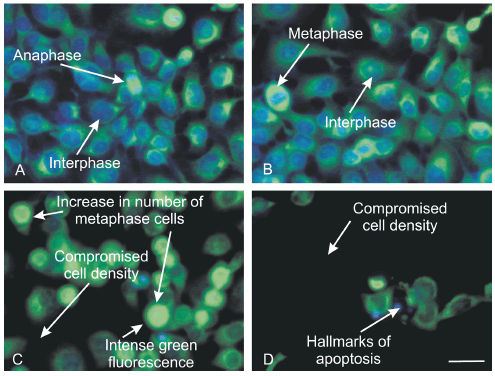
FIGURE 4. Triple-stained SNO cells propagated in growth medium (A), vehicle-treated control cells (B), 0.4μM 2-MeOE2bisMATE-treated cells (C), actinomycin-D treated cells (D) and starved cells (E). The latter served as a positive control for autophagy. 2-MeOE2bisMATE-SNO-treated cells revealed an extensive green and blue fluorescent indicating that both apoptosis and autophagy were induced by 2-MeOE2bisMATE. 2-MeOE2bisMATE-treated cells were rounded in appearance due to a metaphase block (magnification 400x, scale bar indicates 30 μm).
Discussion
The effects of 0.2-1µM 2-MeOE2bisMATE on the growth of SNO cells were investigated after an exposure period of 24 hours. It was observed that a concentration of 0.4µM of 2-MeOE2bisMATE decreased cell growth by 74%. Concentrations of 2-MeOE2bisMATE that were lower than 0.4µM did not reveal any statistically significant differences in the reduction of cell numbers when compared to the vehicle-treated controls and the cells that were propagated in growth medium. Cells exposed to 0.5 µM and 0.6µM of 2-MeOE2bisMATE respectively, demonstrated a slight decrease in cell growth while those cells which were exposed to 0.7-1µM of 2-MeOE2bisMATE showed a decrease in growth to approximately the same extend as those cell which were exposed to 0.4µM of 2-MeOE2bisMATE.
This observed biphasic effect of 2-MeOE2bisMATE is also a characteristic of the parental drug, which is 2-methoxyestradiol (Lottering et al., 1992; Pribluda et al., 2000). In some previous studies, a concentration of 0.4µM of 2-MeOE2bisMATE has been shown to cause a 50% growth inhibition (IC50) of MCF-7 cells (Utsumi et al., 2005; Raobaikady et al., 2003). Another preliminary study showed that 0.5 µM of 2-MeOE2bisMATE inhibited CAL51 growth in an MTS cell proliferation assay in a dose dependent manner (Wood et al., 2004), thus 2-MeOE2bisMATE does have anti-proliferative effects within this concentration range. Characteristics of apoptosis such as round cells, hypercondensed chromatin and apoptotic bodies were visible within the SNO cell line exposed to 0.4 µM of 2-MeOE2bisMATE and the cells' densities was also compromised.
These results were consistent with those of a preliminary study which was conducted by our laboratory using human breast epithelial cancer cells (MCF-7), the results revealed that 0.4 µM of 2-MeOE2bisMATE had induced high chromatin condensation of MCF-7 cells, apoptotic bodies were also observed and it was apparent that mitosis was inhibited in these cells after an exposure period of 48 hours (Visagie and Joubert, 2008). Similarly, Raobaikady et al. (2003) found that a significant increase in the number of rounded, shrunken, and detached MCF-7 cells after treating the cells for 24 hours with 2-MeOE2bisMATE at a concentration of 1 µM. 2-MeOE2bisMATE also induced cell cycle arrest and apoptosis in both the MCF-7DOX and MCF-7WT cells (Newman et al., 2008). The induction of apoptosis by 2-MeOE2bisMATE was further illustrated by an in vitro study which showed that 2-MeOE2bisMATE caused a G2/M cell-cycle arrest within 24 hours in both prostate and ovarian cell lines and a sub-G1 peak (indicative of apoptotic cells) (Day et al., 2003). The exact mechanism by which 2-MeOE2bisMATE induces apoptosis is still unknown, however, earlier studies have shown that 2-MeOE2bisMATE initiates apoptosis, possibly by phosphorylation of BCL-2, by inducing the activity of the pro-apoptotic protein p53, and by activating caspases 3 and 9 (Day et al., 2003; Wood et al., 2004; Newman et al., 2008). This pilot study revealed that 0.4µM of 2-MeOE2bisMATE significantly inhibited cell growth and induced cell death by both apoptosis and autophagy in SNO cells after 24 hours of exposure. These findings of autophagy induction were of novelty, since no previous studies have shown the induction of autophagy in cancer cells by 2-MeOE2bisMATE. However, Azad et al. (2009) have shown that the parental compound (2ME2) does induce autophagy; thus 2-MeOE2bisMATE could also have this effect on cells. This may also suggest a link between the two types of cell deaths. Future studies will be conducted to further investigate the mechanism(s) of action of 2-MeOE2bisMATE.
Acknowledgements
Grants for this study were awarded to Prof. A.M. Joubert (University of Pretoria) from the Cancer Association of South Africa (AK246), the Medical Research Council (AG374, AK076), National Research Foundation, the RESCOM grant from the University of Pretoria and the Struwig-Germeshuysen Trust (AJ038).
References
1. Azad MB, Chen Y, Gibson SB (2009). Regulation of autophagy by reactive oxygen species (ROS): Implications for Cancer Progression and Treatment. Antioxidants & Redox Signaling 11 (4): 777-790. [ Links ]
2. Cushman M, He HM, Katzenellenbogen JA, Lin CM, Hamel E (1995). Synthesis, antitubulin and antimitotic activity, and cytotoxicity of analogs of 2-methoxyestradiol, an endogenous mammalian metabolite of estradiol that inhibits tubulin polymerization by binding to the colchicine binding site. Journal of Medical Chemistry 38: 2041-2049. [ Links ]
3. Davies-Coleman MT, Froneman W, Keyzers RA, Whibley C, Hendricks D, Samaal T, McQuaid C (2005). Anti-oesophageal cancer activity in extracts of deep-water Marion Island sponges. South African Journal of Science 101: 489-490. [ Links ]
4. Day JM, Newman SP, Comninos A, Solomon C, Purohit A, Leese MP, Potter BVL, Reed MJ (2003). The effects of 2-substituted oestrogen sulphamates on the growth of prostate and ovarian cancer cells. Journal of Steroid Biochemistry & Molecular Biology 84: 317-325. [ Links ]
5. Dingli D, Timm M, Russell SJ, Witzig TE, Rajkumar SV (2002). Promising preclinical Activity of 2-Methoxyestradiol in Multiple Myeloma. Clinical Cancer Research 8: 3948-3954. [ Links ]
6. Foster PA, Ho YT, Newman SP, Kasprzyk PG, Leese MP, Potter BVL, Reed MJ, Purohit A (2008). 2-MeOE2bisMATE and 2-EtE2bisMATE induce cell cycle arrest and apoptosis in breast cancer xenografts as shown by a novel ex vivo technique. Breast Cancer Research Treatment 111 (2): 251-260. [ Links ]
7. Fotsis T, Zhang Y, Pepper MS, Adlercreutz H, Montesano R, Nawroth PP, Schweigerer L (1994). The endogenous oestrogen metabolite 2-methoxyoestradiol inhibits angiogenesis and suppresses tumour growth. Nature 368: 237-239. [ Links ]
8. Ireson CR, Chander SK, Purohit A, Perera S , Newman SP, Parish D, Leese MP, Smith AC, Potter BVL, Reed MJ (2004). Pharmacokinetics and efficacy of 2-methoxyoestradiol and 2-methoxyoestradiol-bis-sulphamate in vivo in rodents. British Journal of Cancer 90: 932-937. [ Links ]
9. Lakhani NJ, Sarkar MA, Venitz J, Figg WD (2003). 2-Methoxyestradiol, a promising anticancer agent. Pharmacotherapy 23: 165-172. [ Links ]
10. Lippert C, Seeger H, Mueck AO (2000). The effect of endogenous estradiol metabolites on the proliferation of human breast cancer cells. Life Sciences 72: 877-883. [ Links ]
11. Lottering M-L, Hang M, Seegers JC (1992). Effects of 17-ß-estradiol metabolites on cell cycle events in MCF-7 cells. Cancer Research 52: 5926-5932. [ Links ]
12. Newman SP, Ireson CR,Tutill HJ, Day JM, Parsons MFC, Leese MP, Potter BVL, Reed MJ, Purohit A (2006). The role of 17ß-bydroxysteroid dehydrogenases in modulating the activity of 2-methoxyestradiol in breast cancer cells. Cancer Research 66: 324-330. [ Links ]
13. Newman SP, Foster PA, Stengel C, Day JM, Ho YT, Leese MP, Potter BVL, Reed MJ, Purohit A (2008). STX140 is efficacious in vitro and in vivo in taxane-resistant breast carcinoma cells. Clinical Cancer Research 14: 597-606. [ Links ]
14. Pribluda VS, Gubish ER Jr, LaVallee TM, Treston A, Swartz GM, Green SJ (2000). 2 Methoxyestradiol: an endogenous antiangiogenic and antiproliferative drug candidate. Cancer Metastasis Reviews 19: 173-179. [ Links ]
15. Purohit A, Woo LWL, Chander SK, Newman SP, Ireson C, Ho Y, Grasso A, Leese MP, Potter BVL, Reed MJ (2001). Non-steroidal and steroidal sulfamates: new drugs for cancer therapy. Molecular & Cell Endocrinology 171: 129-135. [ Links ]
16. Purohit A, Woo LWL, Chander SK, Newman SP, Ireson C, Ho Y, Grasso A, Leese MP, Potter BVL, Reed MJ (2003). Steroid sulphatase inhibitors for breast cancer therapy. Journal of Steroid Biochemistry & Molecular Biology 86: 423-432. [ Links ]
17. Raobaikady B, Purohit A, Chander SK, Woo LWL, Leese MP, Potter BVL, Reed MJ (2003). Inhibition of MCF-7 breast cancer cell proliferation and in vivo steroid sulphatase activity by 2-methoxyoestradiol-bis-sulphamate. Journal of Steroid Biochemistry & Molecular Biology 84: 351-358. [ Links ]
18. Sano T, Hirasawa G, Takeyama J, Darnel AD, Suzuki T, Moriya T, Kato K, Sekine H, Ohara S, Shimosegawa T, Nakamura J, Yoshihama M, Harada N, Sasano H (2001). 17 betahydroxysteroid dehydrogenase type 2 expression and enzyme activity in the human gastrointestinal tract. Clinical Science (London) 101: 485-491. [ Links ]
19. Schumacher G, Neuhaus P (2001). The physiological estrogen metabolite 2-methoxyestradiol reduces tumor growth and induces apoptosis in human solid tumors. Journal of Cancer Research and Clinical Oncolology 127: 405-410. [ Links ]
20. Suzuki RN, Newman SP, Purohit A, Leese MP, Potter BVL, Reed MJ (2003). Growth inhibition of multi-drug-resistant breast cancer cells by 2-methoxyoestradiol-bis-sulphamate and 2-ethyloestradiol-bis-sulphamate. Journal of Steroid Biochemistry & Molecular Biology 84: 269-278. [ Links ]
21. Tagg SLC, Foster PA, Leese MP, Potter BVL, Reed MJ, Purohit A, Newman SP (2008). 2-Methoxyoestradiol-3,17-O,O-bissulphamate and 2-deoxy-D-glucose in combination: a potential treatment for breast and prostate cancer. British Journal of Cancer 99: 1842-1848. [ Links ]
22. Thaver V, Lottering M, van Papendorp D, Joubert A (2009). In vitro effects of 2-methoxyestradiol on cell numbers, morphology, cell cycle progression, and apoptosis induction in oesophageal carcinoma cells. Cell Biochemistry Function 24: 205-210. [ Links ]
23. Utsumi T, Leese MP, Chander SK, Gaukroger K, Purohit A, Newman SP, Potter BVL, Reed MJ (2005). The effects of 2-methoxyoestrogen sulphamates on the in vitro and in vivo proliferation of breast cancer cells. Journal of Steroid Biochemistry & Molecular Biology 94: 219-227. [ Links ]
24. Visagie MH, Joubert AM (2008). Influence of 2-methoxyestradiolbis-sulphamate on cell growth, morphology and cell death in MCF-7 and MCF-12A cells. Hons. Poster, Department of Physiology, University of Pretoria, South Africa. [ Links ]
25. Wood L, Leese MP, Mouzakiti A, Purohit A, Potter BVL, Reed MJ, Packham G (2004). 2-MeOE2bisMATE induces caspase-dependent apoptosis in CAL51 breast cancer cells and overcomes resistance to TRAIL via cooperative activation of caspases. Apoptosis 9: 323-332. [ Links ]
Received: April 29, 2010.
Acepted: September 19, 2010.

