










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBiocell
versão impressa ISSN 0327-9545
Biocell vol.35 no.1 Mendoza jan./abr. 2011
ORIGINAL ARTICLES
Bovine parthenogenotes produced by inhibition of first or second polar bodies emission
Romina J. Bevacqua, Rafael Fernandez-Martin, Daniel F. Salamone
Laboratorio de Biotecnología Animal, Facultad de Agronomía, Universidad de Buenos Aires. Av. San Martín 4453. C1417DSE. Argentina.
*Address correspondence to:
Daniel Salamone. E-mail: salamone@agro.uba.ar
Abstract: Parthenogenetic embryos are an ethically acceptable alternative for the derivation of human embryonic stem cells. In this work, we propose a new strategy to produce bovine parthenogenetic embryos inhibiting the emission of the first polar body during in vitro maturation, and allowing the extrusion of the second polar body during oocyte activation. Cytochalasin B, an inhibitor of actin microfilaments, was employed during in vitro maturation to inhibit first polar body emission or during parthenogenetic activation to block second polar body emission. Only one polar body was inhibited in each strategy in order to keep the diploid chromosome set. In experiment 1, the effect of cytochalasin B on in vitro maturation of bovine oocytes was evaluated. Most oocytes (77%) were arrested at a meiotic stage characterized by the presence of a large internal metaphase plate and absence of polar body. In experiment 2, development of embryos exposed to cytochalasin B during in vitro maturation (CytoB-IVM) or during activation (CytoB-ACT) was compared. Developmental rates did not differ between diploidization strategies, even when three agents were employed to induce activation. Both groups, CytoB-IVM and CytoB-ACT, tended to maintain diploidy. CytoB-IVM parthenogenesis could help to obtain embryos with a higher degree of homology to the oocyte donor.
Key words: Meiotic maturation; Microfilaments; Cytochalasin B; Parthenogenesis
Introduction
Parthenogenesis is the growth and development of embryos out of oocytes that have not been fertilized by sperm, and it occurs naturally in many invertebrates as well as in some vertebrates. It can be induced chemically (Susko-Parrish et al., 1994; Presicce and Yang, 1994), and the produced embryos have been widely used to understand early development events (Liu et al., 1998a,b). Interest in parthenogenetic embryos has increased after demonstrating their potential as a source of embryonic stem cells (Kim et al., 2007). Recently, parthenogenetic embryonic stem cells have been isolated from blastocysts on mice and primates (Allen et al., 1994; Cibelli et al., 2002), opening new possibilities on regenerative medicine. These embryonic stem cells have the ethical advantage of not involving the destruction of viable embryos (Cibelli et al., 2006), but the limitation of being homozygous for most genes (Robertson et al., 1983; Surani et al., 1984).
The most common strategy to produce bovine parthenogenetic embryos consists on chemical activation of metaphase II oocytes. During in vitro maturation, oocytes arrested at the diplotene stage of the first prophase, meiotic stage commonly known as germinal vesicle, resume meiosis until second metaphase (Pincus and Enzmann, 1935; Edwards, 1965). The most relevant events of maturation are: germinal vesicle breakdown, segregation of homologous chromosomes, extrusion of the first polar body and progression trough the first meiotic division to metaphase II (revised in Mehlmann, 2005; revised in Wassarman and Albertini, 1994). Mammalian oocytes, with one pair of homologous chromosomes, composed by two sister chromatids, remain arrested at metaphase II until fertilization or parthenogenetic activation (Pincus and Enzmann, 1935).
With the aim to obtain parthenogenetic diploid embryos, metaphase II oocytes are chemically activated in the presence of a drug which inhibits second polar body emission (Balakier and Tarkowski, 1976). Embryos produced in this way contain both sister chromatids of one of the homologous chromosomes, and are predominantly homozygous for the majority of their genes (Kubiak et al., 1991). Cytochalasin B, a microfilament polimerization inhibitor, is usually employed to block second polar body emission (Landa and Hájková, 1990; Niemierko, 1975).
In this work, we explored the possibility to obtain bovine parthenogenetic embryos with a higher degree of identity to the oocyte donor. Cytochalasin B was employed during in vitro maturation to inhibit first polar body emission. Moreover, in vitro development of oocytes subjected to in vitro maturation, in presence or absence of cytochalasin B, was compared. Oocytes subjected to in vitro maturation with or without cytochalasin B were parthenogenetically activated in conditions that intended to maintain diploidy in both cases.
Materials and Methods
Experimental Design
Bovine cumulus oocyte complexes were subjected to in vitro maturation, in presence or absence of cytochalasin B, and their meiotic stages were analyzed. Additionally, developmental competence of these matured oocytes was compared. Each experiment was replicated three times. Experiments are detailed below:
Experiment 1: Bovine oocytes meiotic stage was evaluated during in vitro maturation in presence of cytochalasin B since 3 h or 10 h of starting the procedure, and the evaluation times were 6, 9, 10, 12, 15, 18, 21 and 24 h after starting.
Experiment 2: Comparison of developmental competence of bovine oocytes inhibited to emit a polar body during maturation (CytoB-IVM) or during activation (CytoB-ACT) (Fig. 1). Different chemical agents were used to induce parthenogenetic activation. Embryonic ploidy was analyzed.
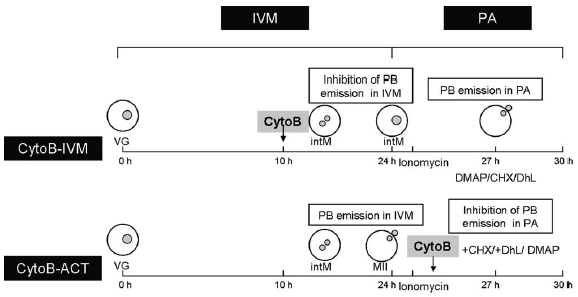
Figure 1. Experiment 2 design: For the group cytochalasin B-in vitro maturation, bovine oocytes were inhibited to extrude a polar body during in vitro maturation by addition of cytochalasin B since 10 h, and then they were activated allowing extrusion of a polar body (ionomycin+3h+dimethylaminopurine; ionomycin+cycloheximide or ionomycin+3h+ dehydroleucodine). For the group CytoB-activation, oocytes were in vitro matured in absence of cytochalasin B, allowing first polar body extrusion, and subsequently activated in conditions that inhibited second polar body extrusion (ionomycin+dimethylaminopurine; ionomycin+cycloheximide+cytochalasin B or ionomycin+dehydroleucodine+cytochalasin B). IVM: In vitro maturation; PA: parthenogenetic activation; PB: polar body; intM: internal metaphase; MII: metaphase II; Cyto B-IVM: cytochalasin B added during in vitro maturation; CytoB-ACT: cytochalasin B added during parthenogenetic activation; Io: Ionomycin; DMAP: 6-dimethylaminopurine; CHX: cycloheximide; DhL: dehydroleucodine.
Reagents
Except otherwise indicated, all chemicals were obtained from Sigma Chemicals Company (St. Louis, MO, USA).
Oocyte collection and in vitro maturation
Bovine ovaries were collected at slaughterhouses and transported to the laboratory in 0.9% NaCl-solution at 25-30ºC. Cumulus oocyte complexes were aspirated from small antral follicles (2-6 mm in diameter) with a 21-gauge needle attached to a 10 ml disposable syringe and were washed in Dulbecco's phosphate buffer saline (DPBS, 14287-072; Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS, 10499-044; Gibco, Grand Island, NY, USA) and 2% antibiotic-antimycotic (ATB, 15240-096; Gibco, Grand Island, NY, USA). Oocytes covered with at least 3 layers of granulosa cells were selected for in vitro maturation. Culture medium for in vitro maturation was bicarbonate-buffered Tissue Culture Medium 199 (TCM199, 31100-035; Gibco, Grand Island, NY, USA) supplemented with 2 mM glutamine (G-8540), 10% FBS, 10 mg/L follicle stimulating hormone (NIH-FSH-P1, Folltropin®, Bioniche, Australia), 0.3 mM sodium pyruvate (P2256), 100 μM cysteamine (M9768) and 2% ATB. For in vitro maturation, cumulus oocyte complexes were cultured in 100 ul droplets of maturation medium (20–25 oocytes/droplet) covered with mineral oil at 39°C in 6.5% CO2 and humidified air.
Cumulus oocyte complexes were removed from culture droplets after 3 h or 10 h of in vitro maturation and incubated in the presence of 10 mg/L cytochalasin B (C6762).
After maturation, cumulus cells were removed by vortexing for 3 min in hyaluronidase (H-4272) (1 g/L DPBS) and washed three times in Tyrode Albumin lactate piruvate buffered with Hepes, (TALP-Hepes) (Bavister and Yanagimachi, 1977).
Meiotic stage analysis
Oocytes were stained with Hoechst Bisbenzimide 33342 (B-2261) (1 mg/L) in TCM-199 for 10 min. Visualization of meiotic stage was performed under UV Light with inverted Nikon Eclipse Diaphot TE-300 epifluorescence microscope (Nikon, NY, EEUU).
Chemical oocyte activation
Oocytes were exposed to 5 μM ionomycin (I24222; Invitrogen, California, USA) in TALP-Hepes for 4 min and washed in DPBS supplemented with FBS and ATB. For the group CytoB-IVM, oocytes were placed in TCM-199 for 3 h to permit extrusion of a polar body and subsequently treated with 2 mM 6-dimetilaminopurine (D2629) in TCM-199 for 3 h or directly incubated with cycloheximide (C8855) 10mg/L in TCM-199 for 5 h or dehydroleucodine 5 uM (Embryology and Histology Institute donation, Mendoza, Argentina) for 3 h. For the group CytoB-ACT, exposure to ionomycin was followed by further activation with 6- dimetilaminopurine, cycloheximide +cytochalasin B and dehydroleucodine+ cytochalasin B. For activation, cytochalasin B was used in 5 mg/L concentration. The inhibitors were removed by washing three times in TALP-H and cultures were continued as described below.
In vitro culture
Presumptive zygotes were returned to the original maturation media and co-cultured with cumulus cells. Cleavage was evaluated at day 2, number of morulae and blastocysts at days 5 and 7 respectively.
Cytogenetic Analysis
At 48 h post-activation, embryos were prepared and examined for their cytogenetic composition, as described by King and Basrur (1979) with minor modifications. For this, embryos were synchronized at metaphase by transferring to medium containing 0.05 mg/L democolcine (KaryoMAX Colcemid, GIBCO BRL, USA) for 6 h. They were treated with 100 ul pronase (P-8811) (15mg/ml) in 2 ml TL-Hepes to disrupt pellucidae zones and subsequently transferred into hypotonic solution (0.8% Na-citrate (F71497), w/v) for 15 min at 37°C. Individual embryos were placed on pre-cleaned microscope slides and fixed by addition of ethanol (1.00983.1000; Merck, Darmstadt, Germany): acetic acid (401422; Carlo Erba, BA, Argentina) (3:1,v/v) mixture. After drying, slides were stained with 5% (v/v) Giemsa solution (Lowens, BA, Argentina) for 10 min and washed. The stained chromosome spreads and nuclei were evaluated at x400 and x1000 magnification with oil-immersion optics. Embryos were classified as being haploid, diploid, tetraploid or others (polyploid or mixoploid).
Statistical Analysis
The data were pooled from at least three replications. Differences in the percentage of oocytes developing to a particular stage were determined by chi-square procedures. The SAS program was used (SAS Institute Inc., 1989). Probability results of P<0.05 were considered to be statistically significant.
Results
Meiotic progression of bovine oocytes during in vitro maturation in presence of cytochalasin B
Meiotic progression of bovine oocytes incubated in cytochalasin B since 3 or 10 h of in vitro maturation beginning was evaluated by Hoechst 33342 staining (Fig. 2).

Figure 2. Meiotic progression of bovine oocytes in vitro matured in cytochalasin B 10 mg/L, since 3 h or 10 h of in vitro maturation. Cyto B: cytochalasin B; MII: metaphase II; intM: internal metaphase; GV: germinal vesicle.
Germinal vesicle - metaphase I transition mainly occurred between 6 and 9 h of in vitro maturation in control and cytochalasin B 10h groups (p<0.05). A different behavior was observed for cytocha lasin B 3h group, which showed this transition between 9 and 12 h (P<0.05).
For both groups incubated in cytochalasin B during in vitro maturation, a high oocyte proportion did not emit the first polar body after 24 h in vitro maturation (76.6% and 77.1% for cytochalasin B 3h and cytochalasin B 10h respectively). In these groups, the most abundant meiotic species consisted on two internal metaphase plates at 21 h, which seemed to rejoin in a large metaphase plate at 24 h (Fig. 3). No differences in metaphase II oocytes proportion were observed between evaluation times for both cytochalasinB groups. In the control group, most of the oocytes reached metaphase II at 18 h (72.2%) (Fig. 2).
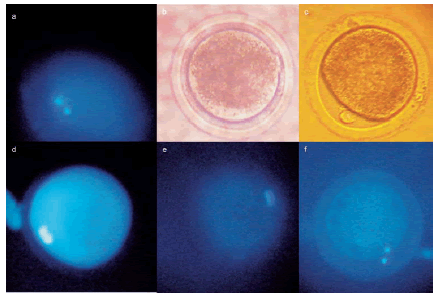
Figure 3. Meiotic progress of bovine oocytes in vitro matured with cytochalasin B (since 10 h in vitro maturation) (a,b,d,e) or without it (c,f). a, d,e) In vitro maturation with cytochalasin B since 10 h and visualization under UV light (488 nm) at different times: a) 15 h; b) 18 h; d) 24 h. f) In vitro maturation without cytochalasin B for 24 h and visualization under UV light. b) and c) Oocytes shown at panels e) and f) after removal of UV light.
Developmental competence of bovine oocytes with polar body emission blocked during maturation or during activation
Developmental competence of bovine oocytes in vitro matured with cytochalasin B since 10h or in vitro matured without cytochalasin B but chemically activated in its presence was compared. Oocytes were selected previous activation based on polar body emission. In CytoB-IVM group, only oocytes inhibited to emit the first polar body were activated. In CytoB-ACT group, only metaphase II oocytes were used (Table 1). No differences were observed in embryo development between diploidization strategies (Table 1). However, when we induced parthenogenetic activation of bovine oocytes incubated with cytochalasin B since 3h or 5h of the beginning of in vitro maturation,embryo development was compromised (data not shown).
Preliminary cytological analysis of embryos showed a high degree of diploid blastomeres in both groups (Table 2), with the exception of activation with ionomycin+ 6-dimethylaminopurine for the group CytoB-ACT. However, no statistical differences were found.
Table 1. Comparative development of oocytes inhibited to emit their first (cytochalasin B-IVM) or second polar bodies (cytochalasin B-ACT), employing chemical activation with 6-dimetilaminopurine, cycloheximide or dehidroleucodine.

Table 2. Cytological analysis of embryos inhibited to emit their first (CytoB-IVM) or second polar bodies (CytoB-ACT), employing chemical activation with 6-dimethylaminopurine, cycloheximide or dehidroleucodine.

Discussion
Our work demonstrates that the presence of cytochalasin B during in vitro maturation provokes a blockage in internal metaphase-metaphase II transition. Moreover, we prove that cytochalasin B causes inhibition of first polar body extrusion during in vitro maturation, but it does not affect subsequent in vitro embryo development.
Our first experiment demonstrates a delay in the transition germinal vesicle-metaphase I when cytochalasin B is added since 3h of in vitro maturation. These results suggest a role for microfilaments on the capture of chromosomes during germinal vesicle breakdown, indicating that micofilaments in concertation with microtubules, lead to proper congression of chromosomes onto the metaphase plate at the metaphase I spindle (Lènart et al., 2005).
In our second experiment, we demonstrated that cytochalasin B induces a reversible meiotic blockage when it is added since 10 h of in vitro maturation beginning, as it allows subsequent in vitro embryo development. This is the first work reporting embryo development of oocytes inhibited to emit the first polar body in the bovine. The degree of heterozygosity of these diploid parthenogenic embryos could be higher than the one observed for embryos obtained after chemical activation of metaphase II oocytes. A recent study in the mice (Kim et al., 2007) demonstrated that activation of oocytes inhibited to go through the first meiotic division, ensured preservation of heterozygosity across the genome except in regions that converted to homozygous by recombination. The diagnostic pattern of a cell line derived from a parthenogenetic embryo obtained by activation of a metaphase II oocyte is pericentromeric homozygosity and an increasing frequency of heterozygosity as distance from the centromere increases. Embryonic stem cells obtained after chemical activation of mouse oocytes inhibited to extrude its first polar body is pericentromeric heterozygosity and increased homozygosity at locations distal to the centromeres (Kim et al., 2007). In our case, further genotypification studies of embryos developed in vitro from oocytes of known genotype would be neccesary to determine the degree of heterozygosity of bovine embryos obtained after both strategies.
Our karyotyping results showed a high degree of diploidies for both activation strategies. However, we also obtained a high proportion of aneuploid and tetraploid blastomeres, which agrees with previous bovine reports, showing that parthenotes exhibit more chromosomic anomalies than embryos obtained by in vitro fertilization (De La Fuente and King, 1998; Bhak et al., 2006).
In summary, our results demonstrate that bovine oocytes can be inhibited to emit their first polar body by microfilaments inhibition, and moreover, that oocytes in vitro matured in these conditions are capable to develop in vitro.
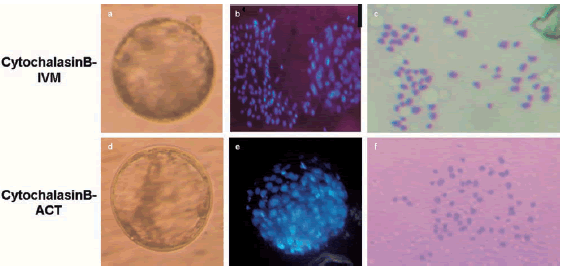
Figure 4. Top panels: Parthenogenesis strategy cytochalasin B-IVM, and activation with CHX; Low panels: Parthenogenesis strategy cytochalasin B-ACT, and activation with DMAP. a and d) Bovine blastocyst under bright light. b and e) Under UV light (488 nm), after Hoechst staining and mounting between slides to determine cell number [b: 170 cells; e) 100 cells]. c and f) Karyotype of day two blastomeres obtained after the same strategies as the blastocysts showed (60+2 chromosomes, magnification 400x).
Acknowledgements
The authors would like to thank slaughterhouses Amancay (Dr. Silva), Nueva Escocia (Dra. Saenz) and Merlo (Marcelo Danielli) for providing the biological material.
References
1. Allen ND, Barton SC, Hilton K, Norris ML, Surani MA (1994). A functional analysis of imprinting in parthenogenetic embryonic stem cells. Development 120: 1473-1482. [ Links ]
2. Balakier H, Tarkowski AK (1976). Diploid parthenogenetic mouse embryos produced by heat-shock and cytochalasin B. Journal of Embryology and Experimental Morphology 35: 25-39. [ Links ]
3. Bavister BD, Yanagimachi (1977). The effects of sperm extracts and energy sources on the motility and acrosome reaction of hamster spermatozoa in vitro. Biology of Reproduction 16: 228-237. [ Links ]
4. Bhak JS, Lee SL, Ock SA, Mohana Kumar B, Choe SY, Rho GJ (2006). Developmental rate and ploidy of embryos produced by nuclear transfer with different activation treatments in cattle. Animal Reproduction Science 92: 37-49. [ Links ]
5. Cibelli JB, Grant KA, Chapman KB, Cunniff K, Worst T, Green HL, Walker SJ, Gutin PH, Vilner L, Tabar V, Dominko T, Kane J, Wettstein PJ, Lanza RP, Studer L, Vrana KE, West MD (2002). Parthenogenetic stem cells in nonhuman primates. Science 295: 819. [ Links ]
6. Cibelli JB, Cunniff K, Vrana KE (2006). Embryonic stem cells from parthenotes. Methods Enzymology 418: 117-135. [ Links ]
7. De La Fuente R, King WA (1998). Developmental Consequences of Karyokinesis Without Cytokinesis during the First Mitotic Cell Cycle of Bovine Parthenotes. Biology of Reproduction 58: 952-962. [ Links ]
8. Edwards RG (1965). Maturation of mouse, sheep, cow, pig, reshus monkey and human ovarian oocytes. Nature 208: 349-351. [ Links ]
9. Kim K, Lerou P, Yabuuchi A, Daley G (2007). Histocompatible Embryonic Stem Cells by Parthenogenesis. Science 315: 482-486. [ Links ]
10. King WA, Basrur PK (1979). Ultrastructural changes in hereditary muscular hypertrophy in cattle. Acta Vet Scand. 20:245-257. [ Links ]
11. Kubiak J, Paldi A, Weber M, Maro B (1991). Genetically identical parthenogenetic mouse embryos produced by inhibition of the first meiotic cleavage with cytochalasin D. Development 111: 763-769. [ Links ]
12. Landa V, Hájková M (1990). Diploidization of bovine oocytes matured in vitro and parthenogenetically activated by electric shock. Folia Biologica (Praha) 36: 145-152. Abstract [ Links ]
13. Lénárt P, Bacher CP, Daigle N, Hand AR, Eils R, Terasaki M, Ellenberg J (2005). A contractile nuclear actin network drives chromosome congression in oocytes. Nature 436: 812-818. [ Links ]
14. Liu L, Ju JC, Yang X (1998a). Differential Inactivation of Maturation-Promoting Factor and Mitogen-Activated Protein Kinase Following Parthenogenetic Activation of Bovine Oocytes. Biology of Reproduction 59: 537-545. [ Links ]
15. Liu L, Ju JC, Yang X (1998b). Parthenogenetic development and protein patterns of newly matured bovine oocytes following chemical activation. Molecular Reproduction and Development 49: 298–307. [ Links ]
16. Mehlmann LM (2005). Stops and starts in mammalian oocytes: recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 130: 791-799. Review. [ Links ]
17. Niemierko A (1975). Induction of triploidy in the mouse by cytochalasin B. Journal of Embryology and Experimental Morphology 34: 279-289. [ Links ]
18. Pincus G, Enzmann EV (1935). The comparative behaviour of mammalian eggs in vivo and in vitro. Journal of Experimental Medicine 62: 655-675. [ Links ]
19. Presicce GA, Yang X (1994). Nuclear dynamics of parthenogenesis of bovine oocytes matured in vitro for 20 and 40 hours and activated with combined ethanol and cycloheximide treatment. Molecular Reproduction and Development 37: 61-68. [ Links ]
20. Robertson EJ, Evans MJ, Kaufman MH (1983). X-chromosome instability in pluripotential stem cell lines derived from parthenogenetic embryos. Journal of Embryology and Experimental Morphology 74: 297-309. [ Links ]
21. SAS Institute (1989). 'SAS/STAT: User's Guide. Version 6. Vol. 1. '4th Edn. (SAS Institute: Cary, NC). [ Links ]
22. Surani MA, Barton SC, Norris ML (1984). Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature 308: 548-550. [ Links ]
23. Susko-Parrish JL, Leibfried-Rutledge ML, Northey DL, Schutzkus V, First NL (1994). Inhibition of protein kinases after an induced calcium transient causes transition of bovine oocytes to embryonic cycles without meiotic completion. Developmental Biology 166: 729-739. [ Links ]
24. Wassarman PM, Albertini DF (1994). The mammalian ovum. In: The physiology of reproduction (E Knobil, J Neil eds.), Vol 1. p. 79-122. Raven Press, New York. [ Links ]
Received: May 16, 2010.
Revised version received: March 16, 2011.
Acepted: March 31, 2011.

