










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkBiocell
versão impressa ISSN 0327-9545
Biocell vol.36 no.2 Mendoza ago. 2012
ORIGINAL ARTICLES
Seasonal testicular changes in Dendropsophus minutus Peters, 1872 (Anura, Hylidae)
Adelina Ferreira1,* and Mahmoud Mehanna2
1 Universidade Federal de Mato Grosso (UFMT), Instituto de Biociências, Cuiabá, MT, Brasil.
2 Universidade Estadual Paulista Julio de Mesquita Filho (UNESP), Laboratório de Biologia de Peixes, Pós-graduação em Zoologia, Botucatu, SP, Brasil.
*Address correspondence to:
Adelina Ferreira.
Universidade Federal de Mato Grosso (UFMT), Instituto de Biociências Cuiabá, MT, BRASIL. E-mail: adelina@ufmt.br
ABSTRACT: The reproductive cycle in anurans may be either continuous or discontinuous. These differences may be connected to seasonal climate changes and/or to anthropic activity. Forty adult male individuals of the Dendropsophus minutus species were collected during one year, in the municipality of Chapada dos Guimarães (Mato Grosso, Brazil). The testicles were studied under light and transmission electron microscopy. No variations were observed when the diameter of the seminiferous tubules and the thickness of the interstitial tissue were studied. However, changes in spermatogenesis were conspicuous and indicated that the reproductive cycle of D. minutus in Chapada dos Guimarães is discontinuous and seems related to variations in air temperature and rainfall.
Key words: Spermatogenesis; Reproduction; Anura; Ultrastructure; Chapada dos Guimarães.
Introduction
Breeding activity of most anurans is associated with the rainy season both in the temperate zone (Wiest, 1972; Salvador and Carrascal, 1990) and the tropics (Heyer, 1973; Barbault and Rodrigues, 1978; Hoogmoed and Gorzula, 1979; Toft and Duellman, 1979; Aichinger, 1987; Gascon, 1991; Duellman, 1995). Yet there are no descriptions of anuran reproductive activity in the Brazilian Cerrado (savanna) bioma.
Dendropsophus minutus (Hylidae) is a small size anuran measuring 20-25 mm (crown-rump length). Its geographical distribution ranges from the lowlands up to 2000 meters in altitude, from low areas to the east of the Andes in Colombia, passing through Venezuela and Trinidad and, moving south, through Ecuador, Peru and Brazil as far as Bolivia, Uruguay and Argentina (Ribeiro et al., 2005).
Chapada dos Guimarães comprises a wide plateau with an altitude varying between 600 and 850 meters. In Mato Grosso, where the most elevated areas are formed by plateaus, the altitude associated with ventilation constitutes the geographical factor that most influences the variations in temperature and rainfall (Maitelli, 2005). Chapada dos Guimarães is located in the center-south of the state of Mato Grosso and is a watershed between the Platina and Amazon basins (Maitelli, 2005). During the year there are two notably different seasons, with regard to pluviometric precipitations: the rainy season (spring and summer) and the dry season (autumn and winter). This seasonal variation may be characterized by six rainy months and six hot months, with oscillations between extreme heat and dry cold (Fig. 3).
The anuran testicles show a seminiferous epithelium with cysts formed from the association of germ cells with Sertoli cells, and all cells present in the same cyst are at the same stage of differentiation (Ferreira et al. 2008; 2009).

Figure 3. Average monthly variations in temperature and rainfall during the studied period.
Material and Methods
Study area
Animals were collected in the permanent Monjolinho stream on the Buriti farm (15°36'S; 56°03'W) in the municipality of Chapada dos Guimarães, state of Mato Grosso, Brazil. This area belongs to the biogeographical domain of the Cerrado (Maitelli, 2005). The average monthly averages of temperature and rainfall during 2006 (Fig. 3), were supplied by INMET (Instituto Nacional de Metereologia - 9th District of the municipality of Cuiabá, Mato Grosso State).
Light microscopy
Adult male individuals were collected monthly from July 2004 to June 2005 (N=50). They were measured and fixed in 10 per cent formalin, preserved in 70 per cent alcohol and deposited in the Zoological Collection of Vertebrates of the Mato Grosso Federal University. One testicle was obtained from each animal, and was fixed in 5% paraformaldehyde, dehydrated in increasing graduations of ethanol, infiltrated and embedded in a methacrylate glycol resin; 3µm sections were obtained and stained with 1% toluidine blue.
Transmission electron microscopy
Testes were fixed overnight at 4ºC in a 0.1M sodium cacodylate buffer solution (pH = 7.2) containing 2.5% glutaraldehyde. After, they were fixed following the above protocol, they were post-fixed for 2 h in 0.1M sodium cacodylate buffer solution (pH = 7.2) containing 1% osmium tetroxide. Next, they were dehydrated in acetone and embedded in LR White resin. Ultra-thin sections were stained with uranyl acetate and lead citrate and observed with a transmission electron microscope (Zeiss, Leo 906).
Results
The testis of Dendropsophus minutus are round and whitish, without noticeable variations in their macroscopical appearance throughout the year.
The structural and ultrastructural analysis of germ cells showed that the primary spermatogonia is an isolated cell that does not form a cyst and is larger in size than the other cells and is located close to the seminiferous tubule wall. Its cytoplasm possesses a great affinity for staining, enabling good visualization of cell limits under light microscopy (Fig. 1a). Secondary spermatogonia are smaller cells organized in cysts of a dark appearance. The primary spermatocyte has a large nucleus with loose chromatin and a thread-like appearance (Figs. 1 b and d). Secondary spermatocytes are smaller than the primary ones, with a roundish and strongly colored nucleus, the chromatin of which is much more compact (Figs. 1 c and e). Spermatids are found in two stages of morphological differentiation: roundish or early and elongated or late. The early spermatids have a spherical nucleus, with relatively compact chromatin, while the elongated ones have a longitudinally extended nucleus (Figs. 1 f and g). The ultrastructure shows the formation of a pre-acrosomal vesicle in one of the nuclear poles of the round early spermatids (Fig. 1f). Following cell differentiation, the nucleus of the late spermatid shows an increasingly more compact chromatin, elongating itself and transforming itself into spermatozoid, which appears strongly colored, and thinner than the spermatid (Figs. 1 h-j). Sperm have a head composed of a nucleus whose chromatin is highly compacted, with few electron-lucent spaces, and an acrosome with an acrosomal vesicle (Fig. 1k). The middle piece is implanted in the nucleus through the penetration of a pair of perpendicular centrioles surrounded by groups of electron-dense granules (Figs. 1 i and j). This piece is constituted by a mitochondrial collar (4 to 8 mitochondria) and a typical axoneme (9+2) with associated dense fibers (Fig. 1l). Between the axoneme and the mitochondria there is a ring of dense fibers (Fig. 1l). The flagellum is simple, consisting of a typical axoneme and the plasma membrane (Fig. 1m).
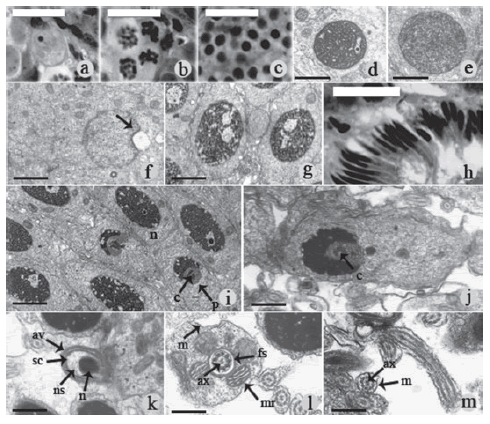
Figure 1: Spermatogenesis stages of Dendropsophus minutus. a-c. Light microscopy. Bar is 20µm. a. Primary spermatogonia. b. Primary spermatocyte. c. Secondary spermatocyte. d-g. Transmission electron microscopy. d. Primary spermatocyte. Bar is 580 nm. e. Secondary spermatocyte. Bar is 580 nm. f. Early spermatid, the arrow indicates the pro-acrosomal vesicle. Bar is 750 nm. g. Early spermatid in phase of compacting chromatin. Bar is 580 nm. h. Light Microscopy, late spermatids with nuclear elongate and compacting chromatin. Bar is 20 µm. i-m. Transmission electron microscopy. i. Late spermatid, in phase compacting chromatin and still evident nucleolus (n), formation of nuclear fossa is the implantation of the midpiece with centriole (c) and showing initial dense bodies (p). Bar is 1 µm. j. Late spermatid with more accented cromatin compactation and implantation nuclear fossa of the midpiece, with central centriole (c). Bar is 580 nm. k. Acrosome apical, nucleus (n), subacrosomal cone (sc), nuclear space (ns) and acrosomal vesicle (av). Bar is 430 nm. l. Middle piece, axoneme (ax), fibrous sheath (fs), mitochondrial ring (mr). Bar is 310 nm. m. End piece, axoneme (ax) and plasma membrane (m). Bar is 310 nm.
Upon analyzing the structural variations in spermatogenesis throughout the year, we observed that there occurs a subtle, but noticeable difference in the proportion of germ cells. However, no variation was noted in the interstitial tissue. All germ cell stages are found in every month of the year (Fig. 2). However, both elongated spermatids and spermatozoa are more frequent than the other cell types from January to April (Figs. 2 a-d) and an appreciable reduction in frequency of these elongated cells was observed from May to September, when spermatogonia and spermatocytes were more frequently found (Figs. 2 e-i). In October to December a subtle increase in the proportion of elongated cells recommenced (Figs. 2 j-l).

Figure 2. a1. Histological images showing the general aspect of the seminiferous tubule of Dendropsophus minutus throughout the year. a. January. b. February. c. March. d. April. e. May. f. June. g. July. h. August. i. September. j. October. k. November. l. December. Bar is 20µm.
The presence of sperm in the lumen of the seminiferous tubules enabled us to identify the period of reproductive activity (November to February) which was coincident with the highest rainfall sperm (Fig. 3). In the following months there was a tendency for spermatogenesis to decrease and about half of the tubules show no sperm in the lumen in August, just before the increase in rainfall (Fig. 3). However, no obvious changes in the diameter of the seminiferous tubules or in the extent of the interstitial tissue were observed in the course of the year.
Discussion
The seminiferous tubules of anurans showing continuous cycles, present cysts at various stages of maturation, with sperm production throughout the year, as it is frequently found in tropical and subtropical species (Saidapur, 1983). However, germ cells mature uniformly in each tubule in anuran species exhibiting discontinuous cycles. Notwithstanding, males of some anuran species of the temperate zone enter a sexually quiescent period after the reproductive period, and spermatogenesis is interrupted for some months (Lofts et al., 1972).
All differentiation stages of spermatogenesis are present in Dendropsophus minutus in October, reaching maximum development in February. This is similar to observations in Hyla pulchella andina (Montero and Pisanó, 1992). In April, when the dry season starts, we observed that spermatogenesis decreases in comparison with the preceding months, and this tendency continues until reaching a peak in August, when secondary spermatogonia become the predominant cells. This pattern of spermatogenesis is very similar to the spermatogenic cycle of Telmatobius pisanoi (Montero and Pisanó, 1990), which halts the spermatogenic process in August, when secondary spermatogonia predominate in the seminiferous tubules. A reactivation of spermatogenesis is indicated by an increase in spermatocyte number in September.
Up to now, the spermatogenic process has received little attention in anurans. The initial phases have only been described in Xenopus laevis (Pipidae) (Kalt, 1976). Similar findings are reported here for D. minutus, mainly in relation to spermatogonia and spermatocyte morphology, and we have added a few more cytoplasmatic characteristics.
Among vertebrates, amphibians present an enormous diversity in their reproductive strategies (Duellman and Trueb, 1994), and this is reflected as large differentiation and variability of the caudal filament, including the protean appendages and membranous parts that this region (Taboga and Dolder, 1993).
The caudal filament presents a typical axoneme (9+2) in D. minutus. Similar axonemes without any caudal annex have been observed in R. pipiens and R. clamitans (Poirier and Spink, 1971) and some Microhylidae (Scheltinga et al., 2002), or with two axonemes in Megophrys montana (Asa and Phillips, 1988) and Lepdobatrachus laevis (Waggener and Carrol Jr., 1998). The caudal filament with axoneme, an undulating membrane and an absent axial rod occur in Pachymedusa dacnicolor (Rastogi et al., 1988) and Bufo gargarizans (Kwon and Lee, 1995). The tail of Hyla japonica consists of an axoneme and an axial rod, but does not show an undulating membrane (Kwon and Lee, 1995). In D. minutus no appendage to the axoneme was observed, except for the fibrous ring at the midpiece region.
Global diversity is changing rapidly as a complex response to various environmental changes caused by man (Vitousek et al., 1997). Globally, intact ecosystems are being degraded at an average rate of 1.2% each year, with the greatest threat to biological diversity and the loss of habitats (Primack and Rodrigues, 2001; Balmford et al., 2002). The Cerrado has for some time suffered high rates of biodiversity loss due to human occupation and agricultural expansion (Klink and Moreira, 2002). For this reason, urgent conservation strategies are important for this bioma.
The decline of amphibian populations, mainly in Australia, Central America and the Pacific north-east of the United States, has been the focus of research projects over the last 12 years (Lanoo, 2005) but amphibian populations in the tropics (as in Ecuador and Brazil) have been insufficiently monitored (Stuart et al., 2004). The causes of amphibian populations' decline, both within and outside protected areas, are still uncertain, but a consensus exists that four main factors may be involved: climate change, pollution, increase in ultraviolet B (UVB) radiation and infectious diseases (Lanoo, 2005). These factors render the group wanting in terms of swift action for conservation strategies.
Acknowledgements
The research was supported by the FAPEMAT (Fundação de Amparo a Pesquisa do Estado de Mato Grosso, process # 0769/2006).
References
1. Aichinger M (1987). Annual activity patterns of anurans in a seasonal Neotropical environment. Oecologia 71: 583-592. [ Links ]
2. Asa CS, Phillips DM (1988). Nuclear shaping in spermatids of the Thai leaf frog Megophrys montana. Anatomical Record 220: 287-290. [ Links ]
3. Balmford A, Bruner A, Cooper P, Costanza R, Farber S, Green RE, Jenkins M, Jefferiss P, Jessamy V, Madden J, Munro K, Myers N, Naeem S, Paavola J, Rayment M, Rosendo S, Roughgarden J, Trumper K, Turner RK (2002). Economic reasons for conserving wild nature. Science 297: 950-953. [ Links ]
4. Barbault R, Trefaut Rodrigues M (1978). Observations sur la reproduction et la dynamique des populations de quelques anoures tropicaux I. Ptychadena maccarthyensis et Ptychadena oxyrhynchus. Terre et Vie 32: 441-452. [ Links ]
5. Duellman WE, Trueb L (1994). Biology of Amphibia. New York, McGraw-Hill Book Company. [ Links ]
6. Duellman WE (1995). Temporal fluctuation in abundances of anurans amphibians in a seasonal Amazonian rainforest. Journal of Herpetology 29: 13-21. [ Links ]
7. Ferreira A, Mehanna M, Prado CAP (2008). Morphologic and morphometric analysis of testis of Pseudis limellum (Cope, 1862) (Anura, Hylidae) during the reproductive cycle in the Pantanal, Brazil. Biocell 32: 185-194. [ Links ]
8. Ferreira A, Santos ABR, Mehanna M (2009). Organização celular dos testículos em Hylidae e Leptodactylidae, no Pantanal (Estado de Mato Grosso do Sul, Brasil). Acta Scientiarum. Biological Sciences 31: 447-452. [ Links ]
9. Gascon C (1991). Population-and community-level analyses of species occurrences of central Amazonian rainforest tadpoles. Ecology 72: 1731-1746. [ Links ]
10. Heyer WR (1973). Ecological interactions of frog larvae at a seasonal tropical location in Thailand. Journal of Herpetology 7: 337-361. [ Links ]
11. Hoogmoed MS, Gorzula SJ (1979). Checklist of the savanna inhabiting frogs of the El Manteco region with notes on their ecology and the description of a new species of treefrog (Hylidae, anura). Zoologische Mededelingen (Leiden) 54: 183-216. [ Links ]
12. Kalt MR (1976). Morphology and kinetics of spermatogenesis in Xenopus laevis. Journal of Experimental Zoology 195: 393-407. [ Links ]
13. Klink CA, Moreira AG (2002). Past and current human occupation, and land use. In: Oliveira PS, Marques RJ (Ed). The Cerrado of Brazil: Ecology and Natural History of a Neotropical Savanna. New York, Columbia University, p. 69-88. [ Links ]
14. Kwon AS, Lee YH (1995). Comparative spermatology of anurans with special references to phylogeny. In: Advances in Spermatozoal Phylogeny and Taxonomy (Special issue). Mémoires du Muséum National d'Histoire Naturelle 166: 321-332. [ Links ]
15. Lanoo M (2005). Amphibian Declines: The Conservation Status of United States Species. Berkeley, University of California, p. 03-06. [ Links ]
16. Lofts B, Wellen J, Benvaad TJ (1972). Seasonal changes in endocrine organs of the male common frog, Rana temporaria. General and Comparative Endocrinology 18: 344-363. [ Links ]
17. Maitelli GT (2005). Interações atmosfera-superfície. In: Moreno G, Higa TCS (Ed.). Geografia de Mato Grosso: território, sociedade e ambiente. pp. 239-249. Editora Entrelinhas, Cuiabá, MT. [ Links ]
18. Montero R, Pisanó A (1990). Ciclo espermatogênico de dos especies de Telmatobius del Noroeste Argentino. Amphibia-Reptilia 11: 97-110. [ Links ]
19. Montero R, Pisanó A (1992). El ciclo espermatogénico anual de Hyla pulchella andina: un análisis numérico. Acta Zoologica Lilloana 41: 173-180. [ Links ]
20. Poirier GR, Spink GE (1971). Ultrastructure of testicular spermatozoa in two species of Rana. Journal of Ultrastructure Research 36: 455-465. [ Links ]
21. Primack RB, Rodrigues E (2001). Biologia da Conservação. Londrina, Midiograf pp.55-67. [ Links ]
22. Rastogi RK, Bagnara JT, Iela L, Krasovich MA (1988). Reproduction in the mexican leaf frog, Pachymedusa dacnicolor. I V. Spermatogenesis: A light and ultrasonic study. Journal of Morphology 197: 249-369. [ Links ]
23. Ribeiro RS, Egito GTBT, Haddad CFB (2005). Chave de Identificação: Anfíbios Anuros da Vertente de Jundiaí da serra do Japí, Estado de São Paulo. Biota Neotropica 5: 235-247. [ Links ]
24. Saidapur SK (1983). Patterns of testicular activity in Indian amphibians. Indian Review of Life Sciences 3: 157-184. [ Links ]
25. Salvador A, Carrascal LM (1990). Reproductive phenology and temporal patterns of mate acess in Mediterranean anurans. Journal of Herpetology 24: 438-441. [ Links ]
26. Scheltinga DM, Jamieson BGM, Bickford D P, Garda AA, Báo SN, McDonald KR (2002). Morphology of the spermatozoa of the Microhylidae (Anura, Amphibia). Acta Zoologica 83: 263-275. [ Links ]
27. Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fischman DL, Waller RW (2004). Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786. [ Links ]
28. Taboga SR, Dolder H (1993). Ultrastructural analysis of the Hyla ranki spermatozoan tail (Amphibia, Anura, Hylidae). Cytobios 75: 85-92. [ Links ]
29. Toft CA, Duellman WE (1979). Anurans of the lower Rio Llullapichis, Amazonian Peru: a preliminary analyses of community structure. Herpetologica 35: 71-77. [ Links ]
30. Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997). Human domination of Earth's ecosystems. Science 277: 494-499. [ Links ]
31. Waggener WL, Carrol Jr EJ (1998). Spermatozoon structure and motility in the anuran Lepidobatrachus laevis. Development, Growth and Differentiation 40: 27-34. [ Links ]
32. Wiest JA, Jr (1972). Anuran succession at temporary ponds in a post oak-savanna region of Texas. M.Sc. Thesis, Texas A&M University. [ Links ]
Received: April 12, 2011.
Revised version received: July 14, 2012.
Accepted: July 17, 2012.

