










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBiocell
Print version ISSN 0327-9545
Biocell vol.37 no.2 Mendoza Aug. 2013
ORIGINAL ARTICLES
Creatine supplementation in trained rats causes changes in myenteric neurons and intestinal wall morphometry
Solange Marta Franzói De Moraes, Thais Andréia Brogio, Jacqueline Nelisis Zanoni, Mariana Cristina Vicente Umada Zapater, Sidney Barnabé Peres, Luzmarina Hernandes*
Universidade Estadual de Maringá, Avenida Colombo 5790, Bloco H79, sala 108 Maringá, Paraná, Brazil, 87020 900
*Address correspondence to:
Luzmarina Hernandes. E-mail: lhernandes@uem.br / luzhernandes@gmail.com
Received: August 31, 2012.
Revised version received: August 14, 2013.
Accepted: August 18, 2013.
ABSTRACT: Creatine is widely used by athletes as an ergogenic resource. The aim of this study was to evaluate the influence of creatine supplementation on the duodenum of rats submitted to physical training. The number and myenteric neuronal cell bodies as well mucosal and muscular tunic morphometry were evaluated. Control animals received a standard chow for 8 weeks, and the treated ones received the standard chow for 4 weeks and were later fed with the same chow but added with 2% creatine. Animals were divided in groups: sedentary, sedentary supplemented with creatine, trained and trained supplemented with creatine. The training consisted in treadmill running for 8 weeks. Duodenal samples were either processed for whole mount preparations or for paraffin embedding and hematoxylin-eosin staining for histological and morphometric studies of the mucosa, the muscular tunic and myenteric neurons. It was observed that neither creatine nor physical training alone promoted alterations in muscular tunic thickness, villus height or crypts depth, however, a reduction in these parameters was observed when both were associated. The number of myenteric neurons was unchanged, but the neuronal cell body area was reduced in trained animals but not when training and creatine was associated, suggesting a neuroprotector role of this substance.
Key words: Nutritional supplementation; Duodenum; Exercise
Introduction
Creatine (=methylguanidinoacetic acid) is an aminoacid used as a nutritional supplement with large application as ergogenic substrate (Kreider et al., 1998; Volek and Rawson, 2004) and a neuroprotector (Matthews et al., 1999; Wyss and Schulze, 2002; Bender et al., 2006). During the last years this substance has become one of the most popular nutritional supplements, aiming the improvement in physical performance during training sessions, mainly because its use reportedly increase lean muscular mass, strength, and muscular power. In face of that, creatine intake has become popular among professional and amateur athletes (Mesa et al., 2002; Branch, 2003; Volek and Rawson, 2004).
The comprehension of creatine synthesis, transport and degradation is the basis for its use as a nutritional supplement and therapeutic compound in treatment of Parkinson's and Huntington's disease, disorders that affect the mitochondria (Klivenyi et al., 1999). It has also been used for the treatment of cardiac muscular atrophy rehabilitation due to disuse, as a neuroprotector in brain hypoxia (Baker-Fulco et al., 2006) and even for the treatment of diabetes (Blum, 2002).
The daily creatine demand is supplied by endogenous synthesis or diet, predominantly by meat intake (Peralta and Amancio, 2002). The endogenous synthesis is down-regulated by diet and when the creatine intake is increased (Mesa et al., 2002).
Creatine is exported from liver and gastrointestinal tract to be stored in tissues such as cardiac and skeletal muscles, which contains significant creatine kinase (CK) concentrations, enzyme responsible for the conversion of creatine to phosphocreatine (PCr), a high energy compound (Garcia, 2000).
The gastrointestistinal tract does not store considerable amounts of creatine, but is the first place to capture and export this substance (Peral et al., 2005). Creatine is coupled to the contractile myosin ring, indicating that a high-energy PCr system is required for an appropriate motor function (Gordon and Keller, 1992). The CK abundance in mitochondria and enterocytes suggests that creatine is part of a system, which keeps temporally and/or spatially the dynamic energetic need of the epithelial cells (Keller and Gordon, 1991).
The creatine intestinal transporters are mainly associated to the apical membrane of villus enterocytes, but is absent in crypts and goblet cells (Peral et al., 2005). The transporters are Na+/Cl- dependent and accumulate creatine against its concentration gradient (Wyss and Kaddurah-Daouk, 2000; Peral et al., 2005) in a stoichiometric ratio of 2 Na:1 Cl: 1 creatine (Peral et al., 2005). Some evidence suggests that changes in this transporter system are genetically programmed and controlled by creatine uptake (Peral et al., 2005).
Occurrence of cramps (Bizzarini and De Angelis, 2004), nausea, vomits and diarrhea (Brudnak, 2004) are often related to the usual creatine consumption by athletes (Wyss and Kaddurah-Daouk, 2000; Brudnak, 2004). Regardless of it, there are no clear evidences associating creatine supplementation to deterioration of the gastrointestinal function. For this reason, the aim of this study was to verify if creatine supplementation in trained and sedentary rats affects intestinal wall morphometry as well the number and morphometry of neurons in the myenteric plexus.
Material and Methods
Animals
Male Wistar rats weighing approximately 300 g, from the Animal Resources Center of the State University of Maringá, were used in the experiments. They were housed in group cages (4 animals per cage) with free access to water and food. The animals were maintained in light/dark cycle of 12h (lights on at 6 am, off at 6 pm) and temperature of 20º ± 2ºC. Control groups rats (sedentary and trained) were fed with a balanced chow (Nuvilab CR1, Nuvital SA, Colombo, Brazil) for 8 weeks, while animals in the treated groups (sedentary and trained rats supplemented with creatine) were fed with the same balanced chow for 4 weeks, but the chow was supplemented with 2% of creatine monohydrate (Probiótica®, São Paulo, Brazil) for 4 additional (Mc-Millen et al., 2001).
The Ethics Committee on Animal Experimentation of the State University of Maringá approved all procedures involving the use of animals.
Experimental procedure
Animals were randomly divided into 4 groups, 4 animals each: sedentary control (S), trained control (T), sedentary supplemented with creatine (SCre) and trained supplemented with creatine (TCre). Physical training lasted 8 weeks, during which the animals were forced to run in a treadmillfor 60 min, 5 times a week, according to the protocol established by Dufloth et al. (1997) and adapted by Negrão et al. (1992). Animals were sacrificed between 7:30 and 10:30h. No physical training sessions were perfomed on the day of sacrifi ce. Both body weight and food intake were recorded twice a week and water consumption three times a week throughout the experiments.
Exercise protocol
The physical exercise consisted of a running training protocol in a programmable treadmill (Inbramed,KT3000, Porto Alegre, Brasil), adapted to run 8 rats simultaneously, from 6:30 to 7:30 am, at an intensity of 60 to 75% of the maximum O2 uptake (VO2 max). Trained animals were submitted to an adaptation week constituted by sessions of 10 minutes/day, in which the treadmill speed ranged from 0.3 Km/h to 0.8 Km/h, in stages that lasted 2 minutes each. After this adaptation period, either the treadmill speed and session endurance were gradually increased until the animals were able to run 1 hour/day, in a average speed of 1.0 km/h during 5 days/week, for a total period of 8 weeks. Each session consisted of various stages, where speed and duration varied, with no changes on treadmill inclination.
Sacrifices and tissue sampling
Animals were anesthetized with sodium pentobarbital (Hypnol® 3%, 4 mg/100g/ body weight, intraperitoneally) and the duodenum was excised out and divided in two segments for either whole mount preparations or for paraffin embedding and hematoxylin-eosin staining.
Whole mount preparations
After washing the intestinal lumen with physiological solution, a cotton thread was set at one end and the lumen was filled with Giemsa fix solution until obtaining an intestinal wall distension similar to that normally provoked by the intestinal content. The other end was then tighten and the whole segment was placed in the fixing solution for 24h. The segments were then opened through their mesenteric side to obtain whole mount preparations of the muscular tunic, by removing the mucosa and submucosal layers under a stereomicroscope. The muscular tunic was stained by Giemsa (Barbosa, 1978), following dehydration, diaphanization, and slides assembling for microscopic analyses. For studying the neuronal profile area, images of myenteric neurons were captured with a QCapture Pro Olympus camera (Tokyo, Japan) coupled to Olympus BX41 microscope (Tokyo, Japan) with 40x objective. The profile (µm2) of 150 cell bodies/animals were measured using Image-Pro-Plus 4 (Media Cibernetics, USA). A total of 600 neurons per group were studied. The number of myenteric neurons was determined in the intermediate region (60º - 120º; 240º - 300º) of the intestinal circumference, considering 0º as the mesenteric insertion (Fig. 1). Neurons were counted in 40 randomized microscopic fields using the Olympus BX41 microscope with 40x objective. The microscopic field area was 0.24 mm2.
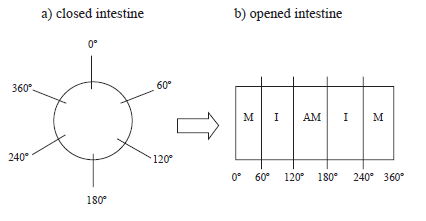
FIGURE 1. The diagram represents (a) closed intestine: 0o comprises the mesenteric insertion; (b) opened intestine: M corresponds to the mesenteric region, I, intermediary region and AM, anti-mesenteric region.
Histological preparations
After removing and washing of intestinal lumen, the segments were opened through the mesenteric border and placed on a cork plate with the epithelial surface up. The samples were then fixed in Bouin fluid for 24 h and submitted to dehydration and paraffin embedding. Finally, semi-serial sections (4 µm thick) were stained with hematoxylin and eosin.
Intestinal wall morphometry
Morphometric analyses were performed with a rule coupled to the 40x objective using the Olympus BX41 microscope. Crypts depth and villus height were determined in longitudinal cuts. The measurements were done on 40 villi from 40 crypts per animal. Muscular tunic thickness was determined in 8 sections per animal at 5 different places (a total of 40 measurements/ animal). Results were expressed in micrometers.
Statistical analysis
The statistical analysis was performed using the GraphPad Prism® 3.0 program. Data are presented as mean ± standard error. A two-way ANOVA was performed for all morphometric parameters and the Tukey test was employed as post-test to multiple comparisons between the groups. For animal weight analysis, the Student t test was used for pair-wise comparisons. In all cases, significance level was set at P < 0.05.
Results
Weight, water and chow consumption
Animals from all groups gained weight during the whole experimental period. Animals from the TCr group presented the lower gain in weight at the end of the experiment (Fig. 2a). The weight loss was estimated in 13% in relation to the S group (p<0.05), 6% in relation to T group and 3% in relation to SCr group. This was associated with a significant reduction in food intake by animals in the TCr group as compared with the S group (Fig. 2b). No differences in water consumption were observed between any of the groups studied.

FIGURE 2. Initial and final (a) body weight (g) and (b) chow consumption (g) in group of sedentary animals (S), sedentary animals supplemented with creatine (SCr), trained animals (T) animals trained and supplemented with creatine (TCr). The asterisk indicates statistically significant differences when compared to the S group. Values were expressed as mean ± SE (n=8), using ANOVA and Tukey's post test.
Myenteric neurons number and neuronal cell body area
The number of myenteric neurons remained unchanged in all studied groups as shown in Figure 3a significant reduction (p<0.01) on neuronal profile area was observed, however, in animals from group T (193.0 ± 3.3 µm2) when compared to either TCr animals (248.4 ± 3.9 µm2) or S animals (246.0 ± 4.6 µm2) (Fig. 3b).

FIGURE 3. (a) Myenteric neurons number /mm2 in the duodenal intermediate region and (b) neuronal profile area (µm2). Abbreviations as in Figure 2. Values were expressed as mean ± SE (n=4), using ANOVA and Tukey's post test. The asterisk indicates statistically significant differences when compared to S and TCr groups.
Muscular tunic, crypts and villus morphometry
Figures 4a, 4b and 4c show the measurements of the muscular tunic (µm), crypts depth (µm) and villus height (µm) respectively, of different studied groups. A significant reduction in all three parameters was observed when physical training and creatine supplementation were associated (TCr group), as compared with both the trained and supplemented groups.

FIGURE 4. Effect of the creatine supplementation on duodenal (a) muscular tunic thickness (µm) (b) cryptal depth (µm) and (c) villus height (µm). Abbreviations as in Figure 2. Values were expressed as mean ± SE (n=4);*p <0.05 when compared to SCr and T groups, using ANOVA and Tukey's post test.
Discussion
Our study showed for the first time that moderate physical training associated with creatine supplementation promoted a reduction on duodenal muscular and mucosal tunics. It was also shown that physical training promoted a significant reduction in the body of myenteric neurons of this part of the intestine.
Water consumption did not vary during the experimental period. On the other hand, there was a significant reduction in the final chow consumption in TCr group. Animals from this group have also presented the lowest weight gain in the end of the experiment. Creatine supplementation in trained animals was able to reduce the weight gain in 13%, the training itself had just reduced 6% and supplementation alone 3%, in comparison to sedentary group. Animals from all studied groups gained weight with time. However, animals from TCr group, the final mean weight was lower (p<0.05) when compared to T and SCr.
Studies have shown that chronic exercise has an important effect over alimentary behavior, stimulating the release of anorexic cytokines (Valsamakis et al., 2004). This effect was not evident in our study since food intake was not different between S and T groups, but the association of aerobic activity with 2% creatine reduced food intake in 16%. However the decrease in food intake was not different among the studied groups when values were corrected on a 100g of body weight basis.
Previous studies, using the same experimental model, showed that endurance exercise training leads to a decrease in fat deposits, without any effect of creatine supplementation (Proença et al., 2007). This is related to the fact that exercise per se is efficient to mobilize the fat reserves in submaximal intensities independently from creatine. However, in a preceding study, Wistar rats submitted to a treadmill running training during 10 weeks and supplemented with 2% creatine, presented higher lean mass and lower fat mass than creatine exercised or control groups. The authors inferred that supplementation in combination with exercise increased the proportion of lean mass more than exercise or creatine alone (Ferreira et al., 2005).
The gastrointestinal tract represents the first barrier for creatine to reach its target tissues. However, there is a lack of information regarding creatine effects on intestinal function and morphology. Creatine absorption may be determined by its physicochemical properties, as well as blood flow (Orsenigo et al., 2005; McCall and Persky, 2007). As creatine is structurally similar to basic aminoacids, it can enter to systemic circulation through amino acids transporters, peptides transporters or specialized transporters. The absence of creatine increase in feces, after supplementation, has indicated that it is totally absorbed by intestinal lumen (Conway and Clark, 1996).
In this study, we have shown that moderate physical training associated with creatine supplementation promotes reduction in muscular tunic thickness, villus height and crypts depth in the duodenum. It is possible that the higher energetic demand provoked by exercise and the increased creatine availability in the intestinal lumen promoted a saturation of the creatine transporters of enterocytes, which could increase energetic availability, leading to a downregulation of proliferation of cryptal cells, thus reducing cryptal depth reduction and villus height.
The interaction between creatine and physical training was also reflected in the smooth muscle since a reduction in muscular tunic thickness was observed. Neither creatine nor exercise itself provoked changes in intestinal wall morphometry.
The myenteric plexus neurons were evaluated considering the density and cell body area. The neurons number did not change; however, the trained group presented a significant reduction (21%) in cell body area. The decrease of the neuronal profile could have been an effect of the decreasing in the nutrient availability toward neurons. The concentration of proteins in tissues is easily affected by nutritional alterations such as changes in quality and quantity of proteins in diet. These changes can directly reflect protein synthesis and ribosomal profile of the rough endoplasmic reticulum, especially in liver and muscles (Goldspink et al., 1984; Lewis et al., 1984). Experimental studies showed the suppression of protein synthesis in both liver and intestine after exercise in rats (Hayase and Yokogoshi, 1992).
It has been estimated that the energy cost for the maintenance of intestinal epithelium and accessory structures represents approximately 20% of the total energy consumed by the animal (McBride and Kelly, 1990). During physical activity the blood flow in gastrointestinal tract can decrease up to 80% (Rowell et al., 1964) depending on the intensity of exercise. The redirection of blood flow to muscles and lungs has been associated to cramps and diarrhea (Peters et al., 2000).
The animals submitted to treadmill running and supplemented with creatine, showed neuronal areas similar to those found in sedentary ones. This result suggests a neurotrophic role for creatine, which is a substrate for creatine kinase (CK), thus increasing phosphocreatine levels, and phosphocreatine in conjunction with the creatine kinase isoenzyme system acts as a potent intracellular energy buffer (Hausmann et al., 2002). Oral creatine supplementation has been shown to elevate phosphocreatine content in the brain, leading to neuroprotective effects (Hausmann et al., 2002) through mechanisms involving maintenance of mitochondrial membrane potential, decreased reactive oxygen species, decreased mitochondrial calcium levels, and preservation of ATP levels (Sullivan et al., 2000). The enteric nervous system is similar to the central nervous system in many aspects (Sternini, 1998) and thus may need a large amount of ATP to maintain its metabolic activities.
This work demonstrated that creatine consumption associated to moderate exercise provoked alterations in the architecture of the intestinal wall and myenteric neurons area of Wistar rats. However, such changes were not associated to any obvious alterations of intestinal function.
Acknowledgments
Our gratitude to Maria Euride Carlo Cancino, Maria dos Anjos Fortunato and Ana Paula Santi Rampazzo for technical support.
References
1. Baker-Fulco CJ, Fulko CS, Kellog MD, Glickman E, Young AJ (2006). Voluntary muscle function after creatine supplementation in acute hypobaric hypoxia. Medical Science Sports and Exercise 38: 1418-1424. [ Links ]
2. Barbosa AJA (1978). Técnica histológica para gânglios nervosos intramurais em preparados espessos. Revista Brasileira de Pesquisa Médica e Biológica 11: 95-97. [ Links ]
3. Bender A, Koch W, Elstner M, Schombacher Y, Bender J, Moeschl M, Gekeler F, Müller-Myhsok B, Gasser T, Tatsch K, Klopstock T (2006). Creatine supplementation in Parkinson disease: a placebo-controlled randomized pilot trial. Neurology 67: 1262-1264. [ Links ]
4. Bizzarini E, De Angelis L (2004). Is the oral creatine supplementation safe? Journal of Sports Medicine and Physical Fitness 44: 411-416. [ Links ]
5. Blum M (2002). Are school nurses using the recommendations of the diabetes control and complications trial in the care of students with diabetes? Journal of School Nursing 18: 138-143. [ Links ]
6. Branch JD (2003). Effect of creatine supplementation on body composition and performance: a meta-analysis. International Journal of Sports Nutrition Exercise and Metabolism 13: 198-226. [ Links ]
7. Brudnak AM (2004). Creatine: are the benefits worth the risk? Toxicology Letters 150: 123-130. [ Links ]
8. Conway MA, Clark JF (1996). In: Creatine and creatine phosphate: scientifical and clinical perspectives (MA Conway, Clark JF, eds.), Academic Press, San Diego. [ Links ]
9. Dufloth DL, Morris M, Michelini C (1997). Modulation of exercise tachycardia by vasopressin in the nucleus tractus solitarii. American Journal of Physiology 273: R1271-1282. [ Links ]
10. Ferreira LG, Bergamaschi CT, Lazareti-Castro M, Heiberg I (2005). Effects of creatine supplementation on body composition and renal function in rats. Medicine Sciences Sports and Exercise 37: 1525-1529. [ Links ]
11. Garcia J (2000). Creatina e exercício: função e aspectos da suplementação. Revista de Nutrição 41: 20-22. [ Links ]
12. Goldspink DF, Lewis SEM, Kelly FJ (1984). Protein synthesis during the developmental growth of the small and large intestine of the rat. Biochemistry 217: 527-553. [ Links ]
13. Gordon P, Keller TCS (1992). Functional coupling to brush border creatine kinase selective energetic advantage to contractile ring myosin in intestinal epithelial cells. Cell Motility and the Cytoskeleton 21: 38-44. [ Links ]
14. Hausmann ON, Fouad K, Wallimann T, Schwab ME (2002). Protective effects of oral creatine supplementation on spinal cord injury in rats. Spinal Cord 40: 449-456. [ Links ]
15. Hayase K, Yokogoshi H (1992). Effect of exercise on tissue protein synthesis in rats. Bioscience, Biotechnology and Biochemistry 56: 1637-1639. [ Links ]
16. Keller TCS, Gordon P (1991). Discrete subcellular localization of a cytoplasmic and a mitochondrial isozyme of creatine kinase in intestinal epithelial cells. Cell Motility and the Cytoskeleton 19: 169-179. [ Links ]
17. Klivenyi P, Ferrante JR, Matthews RT, Bogdanov MB, Klein AM, Andreassem OA, Mueller G, Wermer M, Kaddurah-Daouk, R, Beal MF (1999). Neuroprotective effects of creatine in transgenic animal model of amyotrophic lateral sclerosis. Natural Medicine 5: 347-350. [ Links ]
18. Kreider RB, Ferreira M, Wilson M, Grindstaff P, Plisk S, Reinardy J, Cantler E, Almada AL (1998). Effects of creatine supplementation on body composition, strength, and sprint performance. Medicine and Science in Sports and Exercise 30: 73-82. [ Links ]
19. Lewis SEM, Kelly FJ, Goldspink DF (1984). Pre- and pos- natal growth and protection turnover in smooth muscle, heart and slow-and fast-twitch skeletal muscles of the rat. Biochemical Journal 217: 517-526. [ Links ]
20. Matthews RT, Ferrante RJ, Klivenyi P, Yang L, Klein AM, Mueller G, Kaddurah-Daouk R, Beal MF (1999). Creatine and cyclocreatine attenuate MPTP neurotoxicity. Experimental Neurology 157: 142-149. [ Links ]
21. McBride BW, Kelly JM (1990). Energy costs of absorption and metabolism in the ruminant gastrointestinal tract and liver: A review. Journal of Animal Science 68: 2997-3010. [ Links ]
22. McCall W, Persky AM (2007) Pharmacokinetics of creatine. In: Subcellular Biochemistry, vol. 46 (GS Salomons, M Wyss, eds.) p. 261-273. Springer. [ Links ]
23. McMillen J, Donovan CM, Messer JI, Willis WT (2001). Energetic driving forces are maintained in resting rat skeletal muscle after dietary creatine supplementation. Journal of Applied Physiology 90: 62-66. [ Links ]
24. Mesa JLM, Ruiz JR, Gonzales-Gross MM, Gutierrez SA, Garzon MJC (2002). Oral creatine supplementation and skeletal muscle metabolism in physical exercise. Sports Medicine 32: 903-44. [ Links ]
25. Negrão CE, Moreira ED, Santos MC, Farah VM, Krieger EM (1992) Vagal function impairment after exercise training. Journal of Applied Physiology 1749-1753. [ Links ]
26. Orsenigo MN, Faelli A, De Biasi S, Sironi C, Laforenza U, Paulmichl M, Tosco M (2005) Jejunal creatine absorption: what is the role of the basolateralmembrane? Journal of Membrane Biology 207: 183-195. [ Links ]
27. Peral MJ, Gálvez M, Soria ML, Ilundáin AA (2005). Developmental decrease in rat small intestinal creatine uptake. Mechanisms of Ageing and Development 126: 523-530. [ Links ]
28. Peralta J, Amancio OMSA (2002). A creatina como suplemento ergogênico para atletas. Revista de Nutrição 15: 83-93. [ Links ]
29. Peters HP, Wiersma WC, Akkermans LM, Bol E, Kraaijenhagen RJ, Mosterd WL, de Vries WR, Wielders JP (2000). Gastrointestinal mucosal integrity after prolonged exercise with fluid supplementation. Medicine and Science in Sports and Exercise 32: 134-142. [ Links ]
30. Proença ARG, Nunes LM, de Lima EB, de Moraes SMF (2007). Creatine supplementation and physical training: effects on morphofunctional parameters. Fiep-Federation Internacionale d'Education Physique. 77: 680-682. [ Links ]
31. Rowell LB, Taylor HL, Wang Y, Carlson WS (1964). Saturation of arterial blood with oxygen during maximal exercise. Journal of Applied Physiology 19: 284-286. [ Links ]
32. Sternini C (1998). Structural and chemical organization of the myenteric plexus. Annual Review of Physiology 50: 81-93. [ Links ]
33. Sullivan PG, Geiger JD, Mattson MP, Scheff SW (2000). Dietary supplement creatine protects against traumatic brain injury. Annals of Neurology 48: 723-729. [ Links ]
34. Valsamakis G, McTernan PG, Chetty R, Al Daghri N, Field A, Hanif W, Barnett AH, Kumar S (2004) Modest weight loss and reduction in waist circumference after medical treatment are associated with favorable changes in serum adipocytokines. Metabolism 53: 430-434. [ Links ]
35. Volek JS, Rawson ES (2004). Scientific basis and practical aspects of creatine supplementation for athletes. Nutrition 20: 609-614. [ Links ]
36. Wyss M, Kaddurah-Daouk R (2000). Creatine and creatinine metabolism. Physiological Reviews 80: 1107-1213. [ Links ]
37. Wyss M, Schulze A (2002). Health implication of creatine: Can oral creatine supplementation protect against neurological and atherosclerotic disease? Neuroscience 112: 243-260. [ Links ]

