










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. v.63 n.3-4 Mendoza ago./dic. 2004
Caracterización de las comunidades de acridios (Orthoptera: Acridoidea) del partido de Benito Juárez, sudeste de la provincia de Buenos Aires, Argentina
Grasshopper (Orthoptera: Acridoidea) community structure in Benito Juárez county, in outhern Buenos Aires province, Argentina
Wysiecki María Laura de*, Sandra Torrusio** y María Marta Cigliano***
* Centro de Estudios Parasitológicos y de Vectores (CEPAVE), Calle 2 nro. 584. 1900 La Plata, Argentina; e-mail: lw@museo.fcnym.unlp.edu.ar
** Centro de Procesamiento de Imágenes Satelitarias N. Copérnico. Facultad de Ciencias Exactas y Tecnología, UCALP, Diag. 73 nro. 2137. 1900 La Plata, Argentina; e-mail: torrusio@mail.retina.ar
*** División de Entomología, Museo de Ciencias Naturales, Paseo del Bosque. 1900 La Plata, Argentina; e-mail: cigliano@museo.fcnym.unlp.edu.ar
RESUMEN. Se analizó la riqueza, densidad, constancia y distribución de especies de acridios, en distintas comunidades vegetales (1997-2002) en el partido de Benito Juárez, sudeste de la provincia de Buenos Aires. Las comunidades vegetales se categorizaron en pastizales nativos, comunidades halófilas, pasturas sin disturbio, pasturas con moderado disturbio y pasturas con alto disturbio. En total se registraron 23 especies de acridios. Los melanoplinos dominaron en casi todas las comunidades vegetales, excepto en las halófilas que se caracterizaron por la presencia de un acridino, Covasacris pallidinota (Bruner). La riqueza de especies promedio por sitio por año, varió entre 2,10 ± 0,60 especies en las pasturas con moderado disturbio y 6,20 ± 0,58 especies en las comunidades halófilas. La riqueza de especies acumulada fue de 17 en los pastizales nativos, 14 en las comunidades halófilas, 19 en las pasturas sin disturbio, 18 en las pasturas con moderado disturbio y 14 en las pasturas con alto disturbio. La densidad promedio de acridios fue máxima en el 2001 y 2002. Dichroplus pratensis Bruner y Dichroplus elongatus Giglio-Tos dominaron en los pastizales nativos y en las pasturas con alto disturbio, C. pallidinota en las comunidades halófilas, D. elongatus y Scotussa lemniscata (Stål) en las pasturas sin disturbio y en las pasturas con moderado disturbio. La estructura taxonómica de las comunidades de acridios fue constante en los pastizales nativos, las comunidades halófilas y las pasturas. Hubo pocas especies de amplia distribución e intermedias y las raras fueron las más numerosas. De las 23 especies registradas en todos los sitios, D. elongatus fue la única de amplia distribución. Históricamente, para la zona de estudio, Dichroplus maculipennis (Blanch) fue señalada como la especie dominante. Sin embargo, nuestros resultados sugieren que en la actualidad la especie dominante es D. elongatus. Si bien resulta imposible conocer las causas que motivaron este cambio en la dominancia ya que no se cuenta con datos históricos, no pueden descartarse entre otros factores, eventuales cambios climáticos, campañas acridianas y contra otras plagas y el cambio en los patrones del uso de la tierra ocurridos en las últimas décadas en el área de estudio.
PLABRAS CLAVE. Tucuras. Comunidades. Densidad. Variación temporal. Dichroplus elongatus.
ABSTRACT. The structure of grasshopper communities was studied at different sites in Benito Juárez county, Buenos Aires province, Argentina, over a six-year period (1997-2002). The sites were classified into five categories of disturbance: native grasslands, halophilous communities, pastures, moderately and highly disturbed pastures. A total of 23 grasshopper species was collected. Melanoplinae was the most abundant subfamily in all sites, except for halophilous communities that were characterized by the presence of one Acridinae species, Covasacris pallidinota (Bruner). Average species richness per site and per year, ranged from 2.10 ± 0.60 species in moderately disturbed pastures to 6.20 ± 0.58 species in halophilous communities. Cumulative species richness was: 17 in native grasslands, 14 in halophilous communities, 19 in undisturbed pastures, 18 in moderately disturbed pastures and 14 in highly disturbed pastures. Mean density was significantly higher in 2001 and 2002 than in the remaining years. The most abundant species were Dichroplus pratensis Bruner and Dichroplus elongatus Giglio-Tos in native grasslands and highly disturbed pastures; C. pallidinota in halophilous communities and D. elongatus - Scotussa lemniscata (Stål) in undisturbed and moderately disturbed pastures. Along the study, the taxonomic structure of the grasshopper assemblages was significantly constant in native grasslands, halophilous communities and undisturbed pastures. The pattern of species distribution showed few species widely or intermediately distributed, whereas numerous species were rare. Among the 23 species collected, D. elongatus was the most broadly distributed of all. Historically, Dichroplus maculipennis (Blanch.) has been known as the most conspicuous species in the studied area. However, our results suggest that nowadays D. elongatus is the most common species. Among other factors, climate changes, insecticidal treatments against grasshoppers or other pests, and changes in the land use might be involved in such change in dominance.
KEY WORDS. Grasshoppers. Community structure. Grasshopper density. Temporal variation. Dichroplus elongatus.
INTRODUCCIÓN
Los acridios son los herbívoros dominantes en la mayoría de los sistemas de pastizal. Como consumidores primarios son importantes en el ciclado de nutrientes y de energía y, en años de explosiones poblacionales, compiten con el ganado y la fauna silvestre por el forraje (Fielding & Brusven, 1995). Si bien existe un gran número de trabajos acerca de la biología y la importancia ecológica y económica de estos insectos (Uvarov 1966, 1977; Capinera, 1987; Chapman & Joern,1990; Gangwere et al., 1997; Lockwood et al., 2000, entre otros), aún quedan diferentes aspectos de la estructura y funcionamiento de sus comunidades por estudiar.
En la República Argentina la importancia económica de los acridios ha sido reconocida desde principios del siglo XX y en los últimos años se ha observado en distintas zonas un aumento significativo de sus poblaciones, las que causaron importantes daños en pasturas naturales e implantadas y en diversos cultivos (maíz, soja, etc.) (Cigliano et al., 1995; Cigliano & Lange, 1998; Cigliano et al., 2002). Para combatirlos, en la mayoría de los casos, se utilizaron métodos convencionales basados en la aplicación masiva de insecticidas de síntesis, produciendo un impacto negativo a nivel social, económico y ambiental. Varios factores motivan este tipo de accionar, principalmente la falta de una política ecológica para el control integrado de las especies más perjudiciales de estos insectos, y de monitoreos precisos durante varios años, que permitan evaluar la magnitud del problema y determinar las áreas más afectadas y, finalmente, el desconocimiento de los umbrales de daño y la falta de disponibilidad de los productos biológicos alternativos a los insecticidas.
En 1996 se iniciaron en el partido de Benito Juárez una serie de estudios sobre diferentes aspectos de la ecología de los acridios. Resultados parciales permitieron conocer la interrelación de las especies de acridios con las distintas comunidades vegetales (Torrusio et al., 2002), y analizar la composición y la variación temporal de las densidades de sus comunidades (Cigliano et al., 2002).
El objetivo del presente trabajo es analizar la riqueza y la constancia de especies de acridios, la densidad y la distribución de las mismas, en las distintas comunidades vegetales, en el partido de Benito Juárez, sudeste de la provincia de Buenos Aires, en el período 1997-2002.
MATERIAL Y MÉTODOS
Área de Estudio. El área de estudio está localizada en el partido de Benito Juárez (530.772 ha), al sudeste de la provincia de Buenos Aires (60°30´O, 59°15´O/ 37°15´S 38°00´S). La temperatura media es de 21°C en verano y 7°C en invierno. La precipitación anual promedio varía entre 700 y 800 mm de oeste a este. La vegetación nativa dominante en la región son pastizales naturales con abundantes especies de los géneros Stipa y Piptochaetium (Cabrera, 1968). El área es llana, principalmente usada para agricultura (los cultivos de invierno y verano cubren aproximadamente el 28% del área) y para ganadería (pasturas y pastizales naturales cubren el 60% del área), con la excepción de algunas porciones de sierras (10%) donde aún puede ser encontrada la vegetación prístina.
Sitios de muestreo. Se seleccionaron 20 sitios en 1997, se incrementaron a 27 en 1998 y permanecieron constantes hasta 2002. Los sitios representan la variedad de comunidades vegetales nativas y pasturas cultivadas existentes en la zona, con diferentes grados de disturbio y fueron clasificados en las siguientes cinco categorías:
Pastizales nativos. Cinco sitios dominados por gramíneas nativas (Stipa caudata Trin., Stipa neesiana Trin. et Rupr., Stipa papposa Nees, Piptochaetium stipoides Trin. et Rupr, Piptochaetium medium (Speg.) Torres y Paspalum quadrifarium Lam.)
Comunidades halófilas. Cinco sitios caracterizados por pastos cortos y suelo desnudo, con suelos halomórficos, donde el pasto dominante es Distichlis spicata (L.) Greene..
Pasturas sin disturbio. Seis sitios dominados por especies forrajeras anuales y perennes implantadas (Avena sp., Melilotus officinalis (L.) Pallas, Medicago sativa L., Lolium multiflorum Lam., Thynopyrum ponticum (Podp.) Bardworth et Dewey, entre otras).
Los sitios restantes fueron áreas perturbadas por pastoreo, y se dividieron en dos categorías, de acuerdo a la proporción de especies introducidas implantadas y especies de pastos y hierbas anuales y perennes: pasturas con moderado disturbio (seis sitios) con un 24,16% de hierbas introducidas y un 19,16% de especies forrajeras perennes; y pasturas con alto disturbio (cinco sitios), con un 44% de hierbas introducidas y un 13% de especies forrajeras perennes. Más detalles de la clasificación se brindan en Torrusio et al., 2002.
Muestreo de Acridios. En cada uno de los sitios se realizó el muestreo de acridios, combinando dos métodos de recolección diferentes, de amplia aceptación en este tipo de estudios (Duranton et al., 1982).
La densidad total fue calculada contando el número de insectos que saltaban desde una serie de 30 aros (0,1 m2), colocados cada cinco metros, a lo largo de tres transectas ubicadas al azar, siguiendo el método desarrollado por Onsager & Henry (1977). Al mismo tiempo, en la zona circundante a las transectas, se realizaron 200 golpes de red, de acuerdo a Evans (1984, 1988) y los acridios colectados se llevaron al laboratorio para el recuento e identificación de especies. La abundancia relativa se expresó como la proporción (%) de individuos de una misma especie en relación al número total colectados en cada sitio.
La riqueza de especies de acridios en los diferentes sitios y en cada momento de muestreo se expresó como el número de especies promedio por sitio registrado cada año en cada una de las comunidades vegetales. También se estimó la riqueza de especies de acridios acumulada, que expresa el total de especies registrado en el período de estudio (1997-2002) (Morneau et al., 1999).
La constancia en la estructura taxonómica de las diferentes comunidades de acridios, a lo largo de los seis años del estudio, se estimó a través del cálculo del Coeficiente de Concordancia de Kendall (W), basado en el ranking de importancia de las especies más abundantes en cada comunidad vegetal (Siegel, 1991).
La distribución de las especies de acridios en cada una de las comunidades vegetales y en toda el área de estudio, se estimó a partir de la frecuencia de ocurrencia de cada especie en los diferentes años y momentos de muestreo. Las especies se clasificaron en especies raras (R), si estaban presentes en £ 25% de los sitios-años, especies intermedias (I), si estaban presentes en > 25 y < 75% de los sitios-años y especies de amplia distribución (A), si estaban presentes en ³ 75% de los sitios-año, de acuerdo al agrupamiento sugerido por Kemp (1992).
Los acridios adultos fueron muestreados, en cada sitio, tres veces durante el verano (fin de diciembre, mediados de enero y principios de febrero), a fin de maximizar la detección de especies con diferente fenología en los años 1998 y 2000. El resto de los años se muestreó una sola vez (mediados de enero).
Los datos de densidad, abundancia relativa y de riqueza de especies por sitio y por año fueron promediados para el análisis en aquellos años que se muestreó tres veces durante el verano.
Análisis de datos. Los valores de densidad y de riqueza de especies de los diferentes años, para cada una de las comunidades vegetales y entre las diferentes comunidades vegetales, se compararon mediante la aplicación de dos tests no paramétricos: análisis unidireccional de Kruskal Wallis y análisis para dos muestras de Wilcoxon (SYSTAT 5.02, 1993).
RESULTADOS
Riqueza de especies. En los seis años del estudio se registraron 23 especies de acridios pertenecientes a las Acrididae (13 Melanoplinae, 4 Gomphocerinae, 4 Acridinae, 1 Copiocerinae y 1 Leptysminae) (Tabla I). Los melanoplinos fueron los más abundantes en casi todas las comunidades vegetales, a excepción de las comunidades halófilas donde los acridinos representaron, en promedio, el 46% de los individuos (Tabla II).
Tabla 1: Especies presentes, riqueza de especies acumulada y porcentaje de aparición de cada especie de acridio, para cada comunidad vegetal y para la totalidad de las comunidades (1997-2002), en el partido de Benito Juárez, provincia de Buenos Aires, Argentina.
(R = especies raras; I = especies intermedias; A = especies de amplia distribución).

Tabla 2: Abundancia relativa (expresada en %) de las subfamilias de Acrididae registradas en las diferentes comunidades vegetales (1997-2002), en el partido de Benito Juárez, provincia de Buenos Aires, Argentina.

La riqueza de especies de acridios promedio por sitio, en los diferentes años de muestreo, no mostró diferencia significativa (Kruskal Wallis P > 0,05) entre las distintas comunidades vegetales. La misma varió entre un mínimo de 2,10 ± 0,60 especies en las pasturas con moderado disturbio y un máximo de 6,20 ± 0,58 especies en las comunidades halófilas (Tabla III). El análisis de cada comunidad vegetal a lo largo de los años mostró que existió diferencia significativa en la riqueza de especies en las comunidades halófilas (1998 vs. 2002 y 1999 vs. 2002, Wilcoxon P= 0,039 para ambos), en las pasturas con moderado disturbio (1997 y 1998 vs. 2000, Wilcoxon P= 0,039 y 0,026 respectivamente; 1998 vs. 2002, Wilcoxon P= 0,041 y 1999 vs. 2000 y 2002, Wilcoxon P= 0,039 y 0,038 respectivamente) y en las pasturas altamente perturbadas (1998 vs. 2001 y 2002, Wilcoxon P= 0,043 y 0,042, respectivamente). La riqueza de especies de acridios acumulada en cada sitio varió entre 14 y 19 especies (Tabla I).
Tabla 3: Riqueza promedio de especies (± error estándar) para cada año y riqueza de especies promedio (± error estándar) de los seis años, en las diferentes comunidades vegetales, en el partido de Benito Juárez, provincia de Buenos Aires, Argentina.

Densidad. La densidad promedio de acridios mostró una tendencia creciente a través de los años en la mayoría de las comunidades vegetales, excepto en los pastizales nativos donde no se registraron diferencias significativas (Kruskal Wallis, P > 0,05) (Fig. 1). Las densidades fueron significativamente diferentes en las comunidades halófilas (Kruskal Wallis, P = 0,001), en las pasturas sin disturbio, con moderado y alto disturbio (Kruskal Wallis, P = 0,0001). Los años que fueron significativamente diferentes para cada comunidad vegetal (Wilcoxon, P < 0,05 en todos los casos), se observan en la Figura 1. Comparando todas las comunidades vegetales dentro de un mismo año, el 2001 fue el único que presentó diferencias significativas en las densidades (Kruskal Wallis P= 0,039). El pastizal nativo mostró una densidad significativamente menor que las comunidades halófilas, pasturas sin disturbio y altamente disturbadas (Wilcoxon, P= 0,043 en los tres casos).


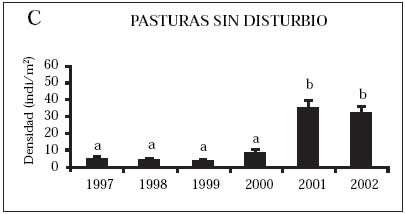


Fig. 1: Densidad promedio de acridios y error estándar en las diferentes comunidades vegetales, partido de Benito Juárez, provincia de Buenos Aires, Argentina (1997-2002).
A) Pastizales nativos, B) Comunidades halófilas, C) Pasturas sin disturbio, D) Pasturas con moderado disturbio, E) Pasturas con alto disturbio.
Años indicados con letras diferentes presentaron diferencias significativas entre sí (Wilcoxon, P < 0.05)
Abundancia relativa. Con respecto a la composición de especies, pocas fueron las que concentraron la mayor proporción de la comunidad. En promedio, las más abundantes en los pastizales nativos y en las pasturas con alto disturbio fueron Dichroplus pratensis Bruner y Dichroplus elongatus Giglio-Tos, en las comunidades halófilas fue Covasacris pallidinota (Bruner), en las pasturas sin disturbio D. elongatus y en las pasturas con moderado disturbio D. elongatus y Scotussa lemniscata (Stål.) (Fig. 2).

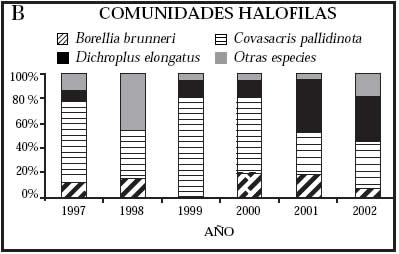



Fig. 2: Abundancia relativa (expresada en %) de las especies de acridios más abundantes en las diferentes comunidades vegetales, partido de Benito Juárez, provincia de Buenos Aires, Argentina (1997-2002).
A) Pastizales nativos, B) Comunidades halófilas, C) Pasturas sin disturbio, D) Pasturas con moderado disturbio, E) Pasturas con alto disturbio.
Constancia en la estructura taxonómica. La estructura taxonómica de las comunidades de acridios a lo largo de los años del estudio, fue constante en los pastizales nativos, las comunidades halófilas y las pasturas sin disturbio. La constancia alcanzó valores estadísticamaente significativos en los pastizales nativos (W= 0,51 P< 0,05) al considerar las cuatro especies más abundantes (D. pratensis, D. elongatus, S. lemniscata y Sinipta dalmani Stål.),en las comunidades halófilas (W = 0,62 P < 0,05) al considerar las tres especies más abundantes (Borellia brunneri (Rehn), D. elongatus y C. pallidinota) y en las pasturas sin disturbio (W = 0,56 P < 0,05) al considerar las cuatro especies más abundantes (D. pratensis, D. elongatus, S. lemniscata y C. pallidinota). En las restantes comunidades, los valores de constancia obtenidos no fueron significativos (pasturas con moderado disturbio: W= 0,33 P> 0,05 y pasturas con alto disturbio: W = 0,26 P > 0,05).
Distribución de especies. A lo largo de los seis años del estudio, hubo pocas especies de amplia distribución e intermedias y las especies raras fueron las más numerosas, tanto al considerar la totalidad de los sitios muestreados, como al considerar cada una de las comunidades vegetales. Dichroplus elongatus fue la única especie de amplia distribución al considerar todos los sitios (Tabla I).
DISCUSIÓN
La riqueza de especies acumulada varió en las diferentes comunidades vegetales, entre un mínimo de 14 especies en las comunidades halófilas y en las pasturas con alto disturbio, y un máximo de 19 en las pasturas sin disturbio. La riqueza promedio de especies por sitio fue similar en las distintas comunidades vegetales y los valores registrados fueron más bajos que los señalados por Sánchez & de Wysiecki (1993) y Cigliano et al. (2000), para diferentes sitios de pastizal y pasturas de la región pampeana (7 a 12 especies por sitio y 4 a 16 especies por sitio, respectivamente).
Con respecto a las subfamilias presentes, los melanoplinos fueron los más abundantes en casi todas las comunidades vegetales, excepto en las comunidades halófilas, que se caracterizaron por la presencia del acridino C. pallidinota.
Los registros sobre las especies de acridios que dominaron cada una de las comunidades vegetales a lo largo del estudio, corroboran los resultados obtenidos por Torrusio et al. (2002), durante la temporada 2000 en Benito Juárez, en donde D. pratensis estaría asociada con los pastizales nativos, C. pallidinota con las comunidades halófilas, D. elongatus con las pasturas sin disturbio y con las pasturas con alto disturbio y D. elongatus - S. lemniscata con las pasturas con moderado disturbio.
La densidad de acridios registrada en 2002 se mantendría en los niveles de explosión poblacional detectados en el 2001 (Cigliano et al., 2002). Las mayores densidades en los años 2001 y 2002 se registraron en las comunidades halófilas y en los tres tipos de pasturas. En las comunidades halófilas, C. pallidinota fue la especie dominante todos los años, excepto en aquellos de explosión poblacional en los cuales disminuyó su proporción, aumentando la de D. elongatus. En las pasturas sin disturbio y en las pasturas con moderado disturbio, D. elongatus y S. lemniscata, representaron la mayor proporción de los individuos, mientras que en las pasturas con alto disturbio fueron dominantes D. elongatus y D. pratensis. Comparando la proporción de D. elongatus entre ambos años, en el 2001 alcanzó los mayores valores, disminuyendo durante el año 2002.
Si bien existieron diferencias en las densidades registradas en los seis años del estudio, la estructura taxonómica de los pastizales nativos y las pasturas sin disturbio fue constante al considerar las cuatro especies más abundantes, mientras que en las comunidades halófilas lo fue al considerar las tres especies más abundantes. Patrones similares fueron señalados por Joern & Pruess (1986), en un estudio temporal de las comunidades de acridios realizado en dos pastizales naturales de América del Norte a lo largo de veinticinco años. Estos autores registraron que la constancia taxonómica estaba dada por las tres y las cinco especies de acridios más abundantes en los pastizales naturales de "North Plate" y "Arapaho Prairie", estado de Nebraska, respectivamente.
El presente estudio no permite conocer cuales son los factores (bióticos y abióticos) que podrían influir en la constancia o la inconstancia de las especies de las comunidades de acridios a través de los años, sin embargo, no se puede descartar que los cambios en la calidad y cantidad del recurso alimenticio entre sitios y entre años, estén entre los más importantes.
Con respecto a la distribución de especies en cada una de las comunidades vegetales, pocas fueron de amplia distribución (D. elongatus, D. pratensis, C. pallidinota). Al considerar la totalidad de los sitios, D. elongatus fue la única especie de amplia distribución. Esta especie ocupa un lugar destacado ya que además dominó las comunidades cuando las densidades alcanzaron niveles de explosión poblacional. Cigliano et al. (2000), en un estudio realizado en 12 sitios del oeste de la provincia de Buenos Aires y este de La Pampa, a lo largo de seis años, también determinaron que D. elongatus fue la única especie de amplia distribución, registrándose en el 97% de los sitios-años. La misma es considerada una de las doce especies más perjudiciales de la República Argentina (COPR, 1982) y se encuentra en casi la totalidad del territorio, excepto en Tierra del Fuego (Cigliano & Lange, 1999).
Para nuestra zona de estudio, Liebermann (1972) señalaba a Dichroplus maculipennis (Blanch.) como la especie característica e históricamente más perjudicial en el sudeste de la provincia de Buenos Aires. Los resultados del presente trabajo la muestran como una especie rara cuando consideramos todas las comunidades vegetales (6%), encontrándose mayormente en las comunidades halófilas (21%). Registros recientes de explosiones poblacionales en este área, parecerían demostrar que D. maculipennis fue desplazada por D. elongatus (Cigliano et al., 1995; Cigliano & Lange, 1998). Específicamente, en la explosión poblacional ocurrida en el verano de 2001, D. elongatus fue la especie que más aportó al aumento de la abundancia, representando el 66% de los 27 individuos registrados por m2 (Cigliano et al. 2002). Cigliano et al.(1995), señalan que, si bien actualmente resulta imposible conocer las causas que motivaron este cambio en la dominancia, no pueden descartarse, entre otros factores, eventuales cambios climáticos, campañas acridianas y contra otras plagas, características propias de la especie, y el cambio en los patrones del uso de la tierra ocurridos en las últimas décadas en el área de estudio. Al respecto, Soriano (1992) y Llorens (1995) indican que los cambios en las actividades agrícolas y ganaderas de la región pampeana determinaron el reemplazo de las pasturas naturales por cultivos y pasturas artificiales, mientras que Viglizo et al. (2002) señalan, entre los cambios importantes ocurridos durante la década del 90 en los sistemas de producción, la interrupción de la rotación entre los cultivos de cosecha y la ganadería, la incorporación de nuevas variedades de cultivos, la aplicación generalizada de plaguicidas de nueva generación y la siembra directa como forma de labranza. Para la zona de estudio, Torrusio (2003) señala que para el período 1995-1999, sucedieron cambios notables en el uso de la tierra, aumentando las dedicadas a la agricultura sobre las ganaderas o las de uso mixto (agrícolo-ganadera), las que disminuyeron en superficie.
Los resultados del presente trabajo, sumados a los de trabajos previos (Torrusio et al., 2002; Cigliano et al. 2002), permitirán implementar acciones que mejoren ambiental y económicamente el control de estos insectos en la región.
AGRADECIMIENTOS
A los evaluadores anónimos por los aportes realizados. Este estudio fue financiado, en parte, por un subsidio (PIP 4015/ 963/97) del Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina, y es parte de un programa de cooperación entre la Facultad de Ciencias Naturales y Museo, UNLP, la Dirección de Aplicación de Imágenes Satelitarias, DAIS, Ministerio de Obras y Servicios Públicos de la Provincia de Buenos Aires (1997-2000), la Comisión de Lucha contra las Plagas y la Municipalidad de Benito Juárez, provincia de Buenos Aires.
BIBLIOGRAFÍA CITADA
1. CABRERA, A. L. 1968. Vegetación de la Provincia de Buenos Aires. En: Cabrera A. L. (ed.) Flora de la Provincia de Buenos Aires I. Colección Científica del Instituto Nacional de Tecnología Agropecuaria, Buenos Aires, pp.101-122. [ Links ]
2. CAPINERA, J. L. 1987. Integrated Pest Management on Rangeland: A shortgrass Prairie perspective. Westview. Boulder, CO. [ Links ]
3. CHAPMAN, R. F. & A. JOERN. 1990. Biology of Grasshoppers. John Wiley & Sons, Inc. New York. [ Links ]
4. CIGLIANO, M. M. & C. E. LANGE. 1998. Orthoptera. En: Morrone J. J. & S. Coscarón (eds.). Biodiversidad de Artrópodos argentinos. Ediciones Sur, La Plata, Argentina, pp. 67-83. [ Links ]
5. CIGLIANO, M. M. & C. E. LANGE. 1999. Dichroplus elongatus. En: Centre for Agriculture and Biosciences International (CABI) (ed.), Global Crop Protection Compendium, Wallingford, Reino Unido. Pp? 9 pp. Publicación electrónica (www. cabicompendium. org/cpc). [ Links ]
6. CIGLIANO, M. M., M. L. de WYSIECKI & C. LANGE. 1995. Disminución de la abundancia de Dichroplus maculipennis en comunidades del sudoeste de la provincia de Buenos Aires. Rev. Soc. Entomol. Argent. 54 (1-4): 41-42. [ Links ]
7. CIGLIANO, M. M., M. L. de WYSIECKI & C. E. LANGE. 2000. Grasshopper (Orthoptera, Acrididae) species diversity in the Pampas, Argentina. Diversity and Distributions 6: 81-91. [ Links ]
8. CIGLIANO M. M., S. TORRUSIO & M. L. de WYSIECKI. 2002. Grasshopper (Orthoptera: Acrididoidea) community composition and temporal variation in the Pampas, Argentina. J. Orthopt. Res. 11 (2): 215-221. [ Links ]
9. COPR (Centre for Overseas Pest Research). 1982. The locust and grasshopper agricultural manual, London. [ Links ]
10. DURANTON, J. F., M. LAUNOIS, M. H. LAUNOIS-LUONG & M. LECOQ. 1982. Manuel de prospection acridienne en zone tropicale séche (2 volumes). Ministére des Relations exterieures – Cooperation et Développement / GERDAT, París. [ Links ]
11. EVANS, E. W. 1984. Fire as a natural disturbance to grasshopper assemblages of tallgrass prairie. Oikos 43: 9-16. [ Links ]
12. EVANS, E. W. 1988. Grasshopper (Insecta: Orthoptera: Acrididae) assemblages of tallgrass prairie: influences of fire frequency, topography, and vegetation. Can. J. Zool. 66: 1495-1501. [ Links ]
13. FIELDING, D. J. & M. A. BRUSVEN. 1995. Ecological correlates between rangeland grasshopper (Orthoptera: Acrididae) and plant communities of southern Idaho. Environ. Entomol. 24 (6): 1432-1441. [ Links ]
14. GANGWERE, S. K., M. C. MURALIRANGAN & M. MURALIRANGAN. 1997. The Bionomics of Grasshoppers, Katydids and Their Kin. CAB International, London. [ Links ]
15. JOERN, A. & K. P. PRUESS. 1986. Temporal constancy in grasshopper assemblies (Orthoptera: Acrididae). Ecol. Entomol. 11: 379-385. [ Links ]
16. KEMP, W. P. 1992. Temporal variation in rangeland grasshopper (Orthoptera: Acrididae) communities in the steppe region of Montana, USA. Can. Entomol. 124: 437- 450. [ Links ]
17. LIEBERMANN, J. 1972. The current state of the locust and grasshopper problem in Argentina. En: Proc. Int. Study Conf. on the Current and Future Problems of Acridology, London, pp.191-198. [ Links ]
18. LOCKWOOD, J.A., A. V. LATCHININSKY & M. G. SERGEEV. 2000. Grasshoppers and Grassland Health. Managing grasshopper Outbreaks without risking Environmental Disaster. Kluwer Acad. Publ. NATO Sciences series 2. Environmental Security. [ Links ]
19. LLORENS, E. M. 1995. The state and transition model applied to the herbaceous layer of Argentina´s Calden Forest. J. Range Manage. 48 (5): 442-447. [ Links ]
20. MORNEAU, F., R. DECARIE, R. PELLETIER, D. LAMBERT, J. DESGRANGES & J. P. SAVARD. 1999. Changes in breeding bird richness and abundance in Montreal parks over a period of 15 years. Landscape Urban Plan. 44: 111-121. [ Links ]
21. ONSAGER, J. A. & J. E. HENRY. 1977. A method for estimating the density of rangeland grasshoppers (Orthoptera, Acrididae) in experimental plots. Acrida 6: 231-237. [ Links ]
22. SÁNCHEZ, N. E & M. L. de WYSIECKI. 1993. Abundancia y diversidad de acridios en pasturas de la Provincia de La Pampa, Argentina. Rev. Invest. Agropec. 24 (1): 29-39. [ Links ]
23. SIEGEL, S. 1991. Estadística no paramétrica aplicada a las ciencias de la conducta. Editorial Trillas, México. [ Links ]
24. SORIANO, A. 1992. Río de La Plata Grasslands. En: Coupland R.T. (ed). Natural grasslands. Introduction and Western hemisphere. Ecosystems of the World,. Elsevier, Amsterdam, pp. 367-407. Systat 5.02, Inc. 1993. Evanston, IL, USA. [ Links ]
25. TORRUSIO, S. 2003. Teledetección y sistemas de información geográfica aplicados a la dinámica de la distribución espacial y temporal de densidades de acridios (Orthoptera: Acridoidea) en el sudeste de la provincia de Buenos Aires (partido de Benito Juárez). Tesis doctoral. Facultad de Ciencias Naturales y Museo. [ Links ]
26. TORRUSIO, S., M. M. CIGLIANO & M. L. de WYSIECKI. 2002. Grasshopper (Orthoptera: Acridoidea) and plant community relationships in the Argentine Pampas. J. Biogeogr. 29: 221-229. [ Links ]
27. UVAROV, B. 1966. Grasshoppers and Locusts: a handbook of general Acridology. Vol. I. Cambridge Univ. Press, Cambridge. [ Links ]
28. UVAROV, B. 1977. Grasshoppers and Locusts: a handbook of general Acridology. Vol. II. Cambridge Univ. Press, Cambridge. [ Links ]
29. VIGLIZO, E. F., A. J. PORDOMINGO, M. G. CASTRO & F. A. LERTORA. 2002. La sustentabilidad ambiental de la agricultura pampeana: ¿oportunidad o pesadilla?. Ciencia Hoy 12 (68): 38-51. [ Links ]
Recibido: 2-II-200
Aceptado: 13-VIII- 2004

