










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. v.65 n.3-4 Mendoza ago./dic. 2006
Efectos de la variación temporal y los métodos de captura en la eficiencia de un muestreo de coleópteros en la Reserva Natural Loma del Medio, El Bolsón, Río Negro
Effects of temporal variation and trapping methods on the efficiency of Coleoptera sampling in Loma del Medio Nature Reserve, El Bolsón, Río Negro
Sackmann, Paula
Laboratorio Ecotono CRUB-Universidad Nacional del Comahue/CONICET. Pasaje Gutiérrez 1125. Bariloche. Río Negro. Argentina; e-mail sackmann@crub.uncoma.edu.ar
ABSTRACT . The present study evaluates for a community of Coleoptera of sub-antartic forest the efficiency of a systematic sampling. In particular, the convenience of distributing sampling effort in time (at different moments within a season and between years) and the use of two different kinds of traps are evaluated. Ten sampling sites were selected in an Austrocedrus chilensis (Cupresaceae) and Nothofagus dombeyii (Fagaceae) forest within the Reserva Forestal Loma del Medio, El Bolsón, Río Negro. At each site, nine pitfall traps were placed covering a 100 m2 surface, along with one Malaise trap. Four annual samplings (January, February, March and April) were carried out during three years (2002-2004). The abundance of species was higher in January-February than in March-April, while richness was similar and community composition differed markedly between periods for the three years. On the other hand, richness was similar when one, two or three years of sampling were considered and in general species composition was similar in different years for comparable periods. However, the accumulation of rare species became stable only after three years of sampling. Malaise traps were more efficient (number of species/individuals captured) than pitfall traps, but the sampling methods were highly complementary, capturing only 25% of species in common. In order to characterize the studied community it is recommended to that sampling efforts are distributed through a sampling season and that different trapping methods are used. Even though one year of sampling under the aforementioned conditions will be enough to have a general description of the Coleoptera community, at least three years will be necessary to reach a high level of sampling integrity. A list of species captured (N = 175), of which 14 are new records for Argentina, is included as an appendix.
KEY WORDS . Coleoptera. Malaise traps. Patagonia. Pitfall traps. Sampling efficiency. Systematic sampling.
RESUMEN . En este trabajo se analiza la eficiencia de un muestreo sistemático para una comunidad de coleópteros de bosque subantártico. En particular se evalúa: 1- la conveniencia de repartir el esfuerzo de muestreo a lo largo del tiempo (dentro de una misma temporada de actividad de insectos y entre años), y 2- la eficiencia del uso de dos tipos de trampas diferentes. Se seleccionaron 10 sitios en un bosque de Austrocedrus chilensis (Cupresaceae) y Nothofagus dombeyii (Fagaceae) en la Reserva Forestal Loma del Medio, El Bolsón, Río Negro. En cada sitio se colocaron nueve trampas de caída que abarcaban una superficie de 100 m2 y una trampa Malaise. Se realizaron cuatro muestreos anuales (enero, febrero, marzo y abril) durante tres años (2002-2004), y de una semana de duración cada uno. La abundancia de especies fue mayor en enero-febrero que en marzo-abril, la riqueza fue similar y la composición de la comunidad fue marcadamente distinta entre dichos períodos para los tres años. Por otro lado, la riqueza de especies fue similar al considerar uno, dos o tres años de muestreo, y en general la composición de la comunidad no varió entre años para períodos comparables. Sin embargo, la acumulación de especies raras sólo se estabilizó luego de tres años de muestreo. Aunque las trampas Malaise fueron más eficientes (número de especies observadas / individuos capturados) que las trampas de caída, los métodos de captura fueron altamente complementarios (25% de especies en común). Para caracterizar esta comunidad en particular, se recomienda distribuir el esfuerzo de muestreo a lo largo de una misma temporada de actividad y aplicar diferentes métodos de muestreos. Con un año de muestreo, bajo las condiciones propuestas anteriormente, se podrá caracterizar la comunidad de forma general, pero hacen falta al menos tres años de muestreo para alcanzar un alto grado de integridad. Se incluye un apéndice con la lista de especies capturadas (N = 175), de las cuales 14 son nuevos registros para Argentina.
PALABRAS CLAVE . Coleoptera. Trampas Malaise. Patagonia. Trampas de caída. Eficiencia de muestreo. Muestreo sistemático.
INTRODUCCIÓN
El conocimiento de la biodiversidad ha sido en los últimos tiempos un desafío de alcance mundial debido al impacto de las actividades humanas sobre los sistemas naturales, y por la dificultad que implica su conocimiento. En particular, resulta un desafío el conocimiento de grupos hiperdiversos como los artrópodos, que han sido menos estudiados que las aves y mamíferos, posiblemente debido a que son considerados menos carismáticos y más difíciles de censar e identificar. Sin embargo, los artrópodos están estrechamente involucrados en los procesos ecosistémicos, ya que son la porción mayoritaria de la diversidad macroscópica total y responden rápidamente a los cambios ambientales (Wilson, 1987; Erwin, 1991; Gaston, 1991; Groombridge, 1992; Kim, 1993; Jones et al ., 1994; Folgarait, 1998; Mc Geoch, 1998).
A pesar de su importancia, es muy poco lo que se conoce en general acerca de la diversidad de artrópodos (Hammond, 1992; Wilson, 1992). La Argentina no escapa a esta situación, ya que el conocimiento de la fauna de artrópodos es muy fragmentario y está restringido a ciertas regiones geográficas (Morrone & Coscarón, 1998). Por ejemplo, los esfuerzos en el conocimiento de la entomofauna asociada a bosques de fagáceas en particular, y a regiones patagónicas en general, han sido escasos (Morrone & Coscarón, 1998; Mario Elgueta, MNHN Santiago de Chile, comunicación personal). Además, esta región es una de las doscientas ecoregiones del mundo que el programa Global 200 considera que merecen máximo valor de protección, debido a su gran riqueza de especies y a su alto número de endemismos, a nivel de géneros y familias entre otros aspectos (Olson & Dinerstein, 2002).
Tradicionalmente los inventarios de artrópodos han sido realizados por taxónomos, empleando técnicas no cuantitativas y métodos de muestreo no estructurados (Longino & Colwell, 1997). Sin embargo, el creciente interés por el conocimiento de la biodiversidad es el responsable de la mayor atención que recibe en la actualidad la aplicación de métodos de muestreo estructurados y replicables. Estos métodos permiten realizar listas de especies completas, en forma eficiente (más especies con menor costo) y caracterizar ensambles de artrópodos de forma que reflejen los procesos ecológicos subyacentes (Oliver & Beattie, 1996; Longino & Colwell, 1997).
En los muestreos estructurados, las especies son registradas en un contexto de muestras replicadas que son estratificadas, con respecto a un conjunto de variables como tiempo, hábitat, método de captura, etc. Esto permite determinar la integridad del muestreo a medida que progresa, evaluar la metodología empleada y eventualmente ajustarla para aumentar su eficiencia (mayor número de especies por individuos capturados). Una consecuencia importante de la realización de este tipo de muestreos es que los resultados de los mismos, pueden ser comparados con los de otras áreas dentro de la misma u otra región, para describir patrones de variación de la diversidad (Fisher, 1999).
En este trabajo se evaluó la eficiencia de un muestreo sistemático como herramienta para el análisis de las variaciones en la abundancia, riqueza y composición de especies de coleópteros.
Los coleópteros son un buen grupo focal para este estudio por ser abundantes y diversos, tanto taxonómicamente como por sus hábitos de vida. En particular, se evaluó la eficiencia de un muestreo sistemático y estratificado de coleópteros con respecto a: 1- momento de realización del muestreo dentro de una misma temporada de actividad (comienzo y fin del verano), 2- la duración total del muestreo (uno, dos y tres años), 3- el tipo de trampas utilizado (caída vs. Malaise). En base a este muestreo se generaron recomendaciones concretas para caracterizar una comunidad de coleópteros en forma íntegra.
MATERIAL Y MÉTODOS
Área de estudio y sitios de muestreo
El área de estudio se ubicó en la la Reserva Forestal Loma del Medio. Esta reserva se encuentra en la región sudoeste de la provincia de Río Negro, en las cercanías de la localidad de El Bolsón, situada entre los 41° 40' y 42° 10' de latitud sur y los 71° 42' y 71° 20' de longitud oeste. El bosque de la reserva pertenece a la Región Austral , Dominio Subantártico, Provincia Subantártica, según la clasificación biogeográfica de Cabrera & Willink (1973), y está dominado por las especies arbóreas ciprés de la cordillera ( Austrocedrus chilensis ) y coihue ( Nothofagus dombeyii ). El radal ( Lomatia hirsuta ) y el maitén ( Maytenus boaria ) son especies con menor representación, y se las encuentra en las partes menos densas del bosque. Las especies dominantes del sotobosque son: el maqui ( Aristotelia maqui ), el chin-chin ( Azara microphylla ), la rosa mosqueta ( Rosa eganteria ), la laura ( Schmis patagonicus ), el calafate ( Berberis spp.) y helechos ( Blechnum sp. y Hymenophyllum sp.) (Dimitri, 1974).
Dentro de la reserva se seleccionaron 10 sitios, los cuales se ubicaron a una distancia de 400 a 2000 m entre sí, y que abarcaron un área de aproximadamente 80 ha . Esta separación se encuentra dentro del rango reportado en estudios similares, se puede suponer que los sitios representan muestras independientes (véase por ejemplo: Koivula et al ., 1999; Eubanks, 2001; Cook, 2003).
Para determinar la abundancia relativa de coleópteros, se utilizaron en todos los sitios dos técnicas diferentes: trampas de caída («pitfall») y trampas Malaise. Aunque existe una diversidad de trampas para capturar coleópteros (por ejemplo, trampas de luz y trampas de interceptación para especies que se dejan caer al ser interceptadas en vez de subir; Borror 1992), las técnicas utilizadas en este trabajo y en particular las trampas de caída, son las más usadas en trabajos de enfoque ecológico.
En cada sitio se colocaron nueve trampas de caída ubicadas en una grilla de 5 x 5 m ( 100 m2 ) y una trampa Malaise. Como trampas de caída se usaron potes de plástico de 9 cm . de diámetro y 12 cm . De profundidad, enterrados al ras del suelo. Cada trampa se llenó en un tercio de su capacidad con una mezcla de agua, detergente y etilenglicol para evitar que los insectos capturados escaparan de la trampa, se descompusieran o desecaran. Al momento de la colecta, el contenido de las nueve trampas de un mismo sitio se juntó en una sola muestra.
Las trampas Malaise son carpas de malla plástica con los lados abiertos y con un frasco colector en su parte superior, que captura y acumula insectos voladores y eventualmente caminadores (Townes, 1972; Borror et al ., 1992). Al igual que en las trampas de caída, el número de individuos capturados de una especie determinada depende de su abundancia y de su grado de movilidad (Spence & Niemelä, 1994). Si el objetivo es comparar la fauna de distintos sitios, el uso de trampas pasivas resulta particularmente útil como método comparativo (Scudder, 2000).
De todas las muestras provenientes de ambos tipos de trampa, se separó la totalidad de los ejemplares de Coleoptera, que se determinaron como especies o morfoespecies, mediante la observación de los caracteres morfológicos típicamente usados para la determinación de cada grupo. La mayor parte del material se envió a especialistas para confirmar la identificación de las especies, o adjudicar identidad a las morfoespecies (ver Agradecimientos). Las especies y morfoespecies correspondientes a este estudio se pueden consultar en el Apéndice 1.
Se realizaron cuatro muestreos de una semana de duración cada uno en los meses de enero, febrero, marzo y abril, con trampas Malaise y trampas de caída; durante los años 2002, 2003 y 2004.
Análisis de los datos
La variación de la abundancia de coleópteros se analizó mediante un ANOVA de medidas repetidas. El factor de medidas repetidas fueron los 12 meses de muestreo. Se aplicó una transformación logarítmica a la variable dependiente (x = log (x +1)), para cumplir con los supuestos requeridos por el ANOVA. Luego de la transformación, los residuos del modelo se ajustaron a una distribución normal. Sin embargo, los resultados se presentan sin transformar y agrupados en dos períodos por año: enerofebrero y marzo-abril (comienzo y fin de verano respectivamente), para facilitar su interpretación.
Debido a que las estimaciones de riqueza de especies pueden ser influidas por diferencias en el tamaño de las muestras (por ejemplo el número de individuos capturados), se usaron curvas de rarefacción de Coleman, basadas en muestras para calcular y comparar la riqueza esperada de especies en los diversos grupos (por ejemplo para las muestras de enero-febrero vs. las de marzo-abril). La rarefacción elimina la variación en la riqueza de especies debida a diferencias en la abundancia de las muestras, mediante el remuestreo repetido y aleatorio de un conjunto de N individuos (Gotelli & Colwell, 2001).
En particular, las curvas de rarefacción de Coleman se calculan redistribuyendo al azar la totalidad de los individuos capturados en las muestras, manteniendo sus tamaños originales; mientras que en las curvas clásicas de rarefacción se muestrean al azar la totalidad de los individuos con un tamaño de muestra fijo (Colwell & Coddington, 1994). Las curvas de rarefacción fueron reescaladas al número de individuos capturados, y comparadas al máximo nivel común de individuos capturados (por ejemplo 2370 individuos para comparar las curvas correspondientes a uno, dos y tres años de muestreo). Los análisis se realizaron con el programa EstimateS versión 6.1 (Colwell, 2000), sumando los datos de enero y febrero (período de alta abundancia = comienzo del verano) y los de marzo y abril (período de baja abundancia = fines del verano). Esta decisión se basó tanto en los análisis de variación de la abundancia (ver más arriba) como en los análisis exploratorios de los datos que mostraron la existencia de una alta homogeneidad, en las muestras de los meses de enero-febrero y marzo-abril, en riqueza (según curvas de rarefacción de Coleman) y composición de especies (según ANOSIM, ver más abajo).
Para evaluar la integridad del muestreo (grado de representatividad de las muestras obtenidas en relación a la riqueza esperada), se analizó el patrón de acumulación de especies raras en relación al esfuerzo de muestreo (número de individuos capturados). Longino & Colwell (1997) sugieren que los inventarios de grupos muy diversos, raramente pueden ser completos y que sólo pueden alcanzar un nivel bajo (definido arbitrariamente) de aparición de nuevas especies en relación al esfuerzo de muestreo. Se analizó el patrón de acumulación de dos tipos de especies raras: a) singletons : especies para las cuales se capturó un solo individuo en total, y b) uniques : especies que aparecieron en una sola muestra del total. Para cada curva se observó la pendiente a altos esfuerzos de muestreo, para determinar si la misma era positiva, negativa o cercana a cero. Mientras que una pendiente positiva indica, que el esfuerzo de muestreo no fue suficiente para obtener muestras representativas de la totalidad del ensamble, las pendientes negativas o cero indican lo contrario. Para estas curvas se usó la escala logarítmica para el eje de las abscisas, ya que esta escala revela patrones que son difíciles de ver en una escala aritmética (Longino et al. , 2002).
Para analizar las diferencias en la composición de especies, se usó el procedimiento ANOSIM (Análisis de Similitudes) del programa PRIMER versión 5 (Clarke & Gorley, 2001). Se generó una matriz de similitud entre sitios para los diferentes períodos de muestreo (enero-febrero y marzo-abril en los tres años de muestreo = seis grupos), se usó el índice de similitud de Bray-Curtis para los datos estandarizados y transformados con raíz cuarta (esta transformación reduce el peso de las especies muy abundantes).
ANOSIM es un procedimiento de permutación no paramétrico que produce un estadístico R, que es una medida absoluta de distancia entre los grupos. Valores positivos y grandes (hasta 1) de R, indican baja similitud entre los grupos; mientras que valores bajos (hasta 0) indican alta similitud entre grupos. El nivel de significación se calcula mediante permutaciones de los sitios entre los grupos. Las relaciones de similitud entre los sitios, se muestran mediante escalamiento multidimensional amétrico (NMSD) usando las mismas matrices de similitud que para ANOSIM. Con el procedimiento SIMPER de PRIMER se detectaron las especies discriminantes de grupos (aquellas que no sólo contribuyen a la disimilaridad entre grupos, sino que lo hacen consistentemente entre todas las muestras de ambos grupos). El ANOSIM, el NMDS y el SIMPER fueron realizados con PRIMER.
RESULTADOS
A lo largo de los tres años de muestreo se capturaron 175 especies y morfoespecies (7.865 individuos totales), de las cuales el 43.4% fue identificado hasta el nivel de especie. El resto del material se separó en morfoespecies, un 25.1% dentro de géneros conocidos, y un 31.4% dentro de familias conocidas (ver Apéndice 1). La familia más abundante fue Carabidae (45% del total de individuos capturados), y luego Staphylinidae que representó el 13% de las capturas. Luego, siguieron las familias Cryptophagidae, Lampyridae, Leiodidae y Scarabaeidae con 6% a 3% de representatividad. Las familias restantes fueron mucho menos abundantes, aproximadamente entre 1 y 100 individuos capturados en total.
Análisis de la eficiencia del muestreo en cuanto al momento de realización dentro de una misma temporada
La abundancia de coleópteros varió significativamente a lo largo del periodo de tiempo analizado (F (11, 99) = 6.0, p < 0.001) pero, para los tres años la misma fue igual en enero y febrero así como en marzo y abril (comparaciones a posteriori con Prueba de Tukey, p < 0.05). Sin embargo, la abundancia fue siempre mayor a principios del verano (en enero-febrero) que a fines del mismo (marzo-abril, comparaciones a posteriori con Prueba de Tukey, p < 0.05); Fig. 1).
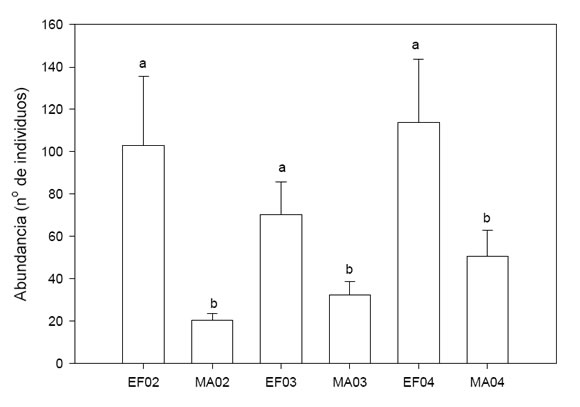
Fig. 1 . Variación de la abundancia estival de coleópteros en la Reserva Loma del Medio a lo largo de tres años. EF corresponde a los meses de enero y febrero, MA, corresponde a los meses de marzo y abril, mientras que 02, 03 y 04 corresponden a los años 2002, 2003 y 2004 respectivamente. Las medias (ES) se calcularon a partir de la abundancia obtenida en los diez sitios, las diferentes letras indican diferencias significativas (Prueba de Tukey con a = 0.05).
A pesar de las variaciones en la abundancia, la riqueza esperada de especies fue muy similar entre esos períodos, durante los tres años analizados (Fig. 2A). En el año 2002 la riqueza de especies en enero-febrero fue de 47, y 51 en marzo-abril (diferencia de 8%), para 410 individuos capturados. En el año 2003 la diferencia en la riqueza de especies, entre los períodos considerados fue sólo de 3% (para 702 individuos, la riqueza de especies fue igual a 61 en enero-febrero, y 59 en marzo-abril). Finalmente en 2004 la riqueza fue 14% más baja en marzo-abril, que en enero-febrero (para 1012 individuos capturados la riqueza fue 49 y 57 respectivamente; Fig. 2A).
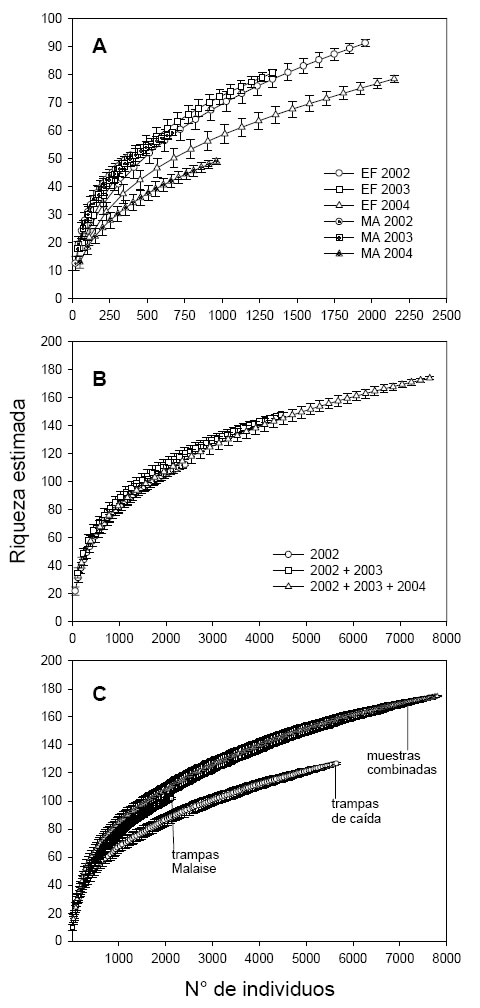
Fig. 2 . Variación de la riqueza esperada de especies en función del número de individuos capturados según curvas de rarefacción de Coleman. (A) a lo largo de tres años (véase Fig. 1 para explicación de las leyendas); (B) en función de los años de muestreo considerados (2002 corresponde a la totalidad de muestras obtenidas ese año, 2002+2003 corresponde a la totalidad de muestras de ambos años, y 2002+2003+2004 corresponde al total de muestras obtenidas a lo largo de tres años ); (C) en función del tipo de método de captura empleado («combinado» corresponde a todas las muestras de trampas de caída y Malaise sumadas). Las barras indican el desvío estándar de la media producida luego de 50 aleatorizaciones.
La composición de especies en enerofebrero difirió significativamente de la de marzo-abril para los tres años analizados (Fig. 3, Tabla I). En 2002 las especies responsables de la discriminación entre los grupos fueron Mimodromites nigrotestaceus (Solier) (Carabidae), Loncovilius discoideus (Fairmaire & Germain) (Staphylinidae), y Trirammatus unistriatus (Dejean) (Carabidae). Las dos primeras especies fueron más abundantes en marzo-abril que en enerofebrero, mientas que ocurrió lo opuesto con la tercera. En 2003 M . nigrotestaceus también fue una de las principales especies discriminantes, más abundante a fines que a comienzos del verano. Otras especies discriminantes fueron Systolosoma breve Solier (Trachypachidae) y Frickius variolosus Germain (Geotrupidae) ambas más abundantes a fines que a principios del verano, y Chiliotis gen. sp. 1 (Cryptophagidae) que mostró una variación de la abundancia opuesta. Finalmente en 2004 M . nigrotestaceus y T. unistriatus aparecieron nuevamente como especies discriminantes de grupos, y con la misma variación de abundancia que en años anteriores. La mayor abundancia de Eunemadus chilensis Portevin (Leiodidae) a principios del verano también determinó la separación entre los grupos.
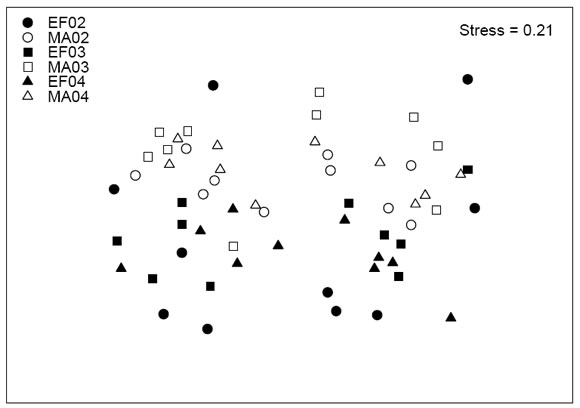
Fig. 3 . Escalamiento multidimensional amétrico (NMDS) basado en la composición estival de especies (véase también Fig. 1)

Tabla I . Resultados del análisis de similitudes (ANOSIM). El valor de R refleja el grado de separación de los grupos comparados en base a su composición de especies: si R tiene un valor cercano a 1 los grupos son diferentes, mientras que si R tiene un valor cercano a 0 los grupos son similares. Los p menores a 0.05 se muestran en negritas. EF = datos para enero y febrero sumados, MA = datos para marzo y abril sumados, 02, 03 y 04 corresponden a los años 2002, 2003 y 2004 respectivamente.
Análisis de la eficiencia del muestreo en cuanto a su duración
La riqueza esperada de especies para un número dado de individuos capturados, fue la misma que cuando se calculó para un solo año de muestreo, para dos y para tres (Fig. 2B). El número de especies fue de 113 cuando se capturaron 2.470 individuos en 2002, de 119 cuando se sumaron las muestras de 2002 y 2003, y de 114 al sumarse las muestras de los tres años. Sin embargo, sólo cuando los tres años de muestreo son considerados en conjunto, el número de especies raras (tanto singletons como uniques ) se mantiene constante con el aumento del esfuerzo de muestreo (por ejemplo, el número de individuos capturados; Fig. 4). Las curvas de acumulación de especies raras en altos esfuerzos de muestreo, correspondientes a uno y dos años de muestreo, revelan pendientes positivas similares (Fig. 4). Esto indica que al sumar las muestras del segundo año, no se incrementa la integridad del muestreo con respecto al primer año.

Fig. 4 . Curvas de acumulación de especies raras en función de los años de muestreo considerados (véase también Fig. 2).
La variación en la composición de especies fue baja (Fig. 3), y el ordenamiento de los sitios se mantuvo constante entre años, especialmente para el período marzo-abril (Tabla I). Sólo una comparación (2003 vs. 2004) mostró diferencias significativas en la composición del ensamble para el período enero-febrero (Tabla I).
Análisis de la eficiencia del uso de dos tipos de trampas
Para bajos esfuerzos de muestreo (menos de 1200 individuos capturados), las trampas de caída y Malaise capturaron el mismo número de especies de coleópteros, y que además es el mismo que para ambos métodos combinados. A partir de los 1200 individuos capturados, las trampas Malaise fueron más eficientes que las trampas de caída en términos de obtención de número de especies. Además, para todos los niveles de esfuerzo de muestreo, las trampas Malaise capturaron tantas especies como los dos métodos combinados (Fig. 2C ). A partir de los 1200 individuos capturados, la tasa de acumulación de especies en las trampas de caída, fue menor que para las trampas Malaise, y también menor que para ambos métodos combinados. Esto significa que para obtener una especie nueva, es necesario capturar más individuos en las trampas de caída que en las Malaise.
Las trampas Malaise capturaron el 58% del total de especies observadas en los tres años, mientras que las trampas de caída capturaron el 69% de especies del total (N = 175). Sólo 48 especies fueron capturadas por ambos métodos, 55 especies fueron exclusivas de muestras obtenidas con trampas Malaise, y 74 con trampas de caída. A pesar de que la combinación de métodos fue tan eficiente como las trampas Malaise, en términos de número de especies/individuos, para el conjunto de todos los coleópteros los dos métodos de captura son altamente complementarios (Fig. 5).
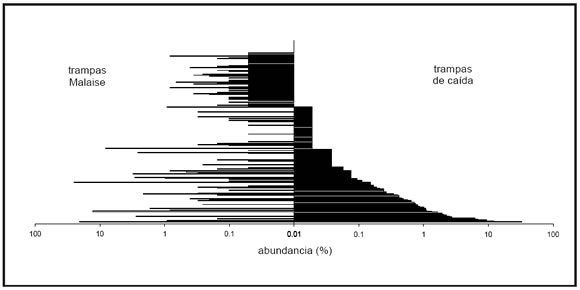
Fig. 5 . Comparación de los dos tipos de trampa utilizados para capturar coleópteros. El lado derecho de la figura muestra la abundancia relativa de las especies, ordenadas de mayor (abajo) a menor (arriba) abundancia, capturadas en trampas de caída. Del lado izquierdo se muestra la abundancia relativa de las mismas especies capturadas en trampas Malaise.
DISCUSIÓN
La utilización de muestreos cuantitativos y estructurados permite evaluar la eficiencia y la distribución del esfuerzo de muestreo en el tiempo y el espacio. Para la comunidad de coleópteros estudiada, se puede concluir que repartir el esfuerzo de muestreo en distintos momentos de una misma temporada de actividad de los insectos, es una buena estrategia ya que hay recambio de especies a lo largo del tiempo, y que el número de especies per se es relativamente constante. Por ejemplo, para los tres años analizados Mimodromites nigrotestaceus fue mucho más abundante en las muestras de finales del verano que en las del comienzo. Esto mismo ocurrió con otras especies que sólo se capturaron en uno u otro de los periodos analizados, por ejemplo, Metius malachiticus (Dejean) (Carabidae) sólo se capturó en el periodo enero-febrero en los tres años. Si el análisis se ampliara a otras épocas del año, como la primavera, las conclusiones serían probablemente las mismas: a mayor cobertura de una temporada de actividad, mayor diversidad de especies.
Es posible obtener una buena caracterización general del ensamble con un solo año de muestreo, si se toman al menos dos períodos diferentes dentro de la temporada de actividad de insectos. La riqueza de especies fue similar cuando se compararon las muestras obtenidas el primer año con las muestras obtenidas en el primero más el segundo, y en el primero más el segundo y el tercero. Además la composición de especies fue muy consistente entre años y para épocas de muestreo equivalentes, esto sugiere que la comunidad analizada es estable a lo largo del tiempo. Sin embargo, pueden haber años en que la riqueza sea más baja de lo habitual, como ocurrió en 2004 para esta comunidad de coleópteros. Si bien la misma puede ser bien caracterizada en su estructura general con un solo año de muestreo, son necesarios al menos tres años para lograr que la aparición de especies raras (es decir singletons y uniques ) se mantenga constante, aunque aumente el esfuerzo de muestreo. Por ejemplo en este estudio, las pendientes de las curvas de acumulación de especies raras fueron positivas con el máximo esfuerzo de muestreo, al considerar las muestras de uno y dos años de muestreo, se observó cierta estabilización de las curvas al tercer año. Por lo tanto, según cuál sea el objetivo de un eventual estudio, un muestreo anual podrá ser suficiente o no.
Las trampas Malaise fueron más eficientes que las trampas de caída, en cuanto a que capturaron mayor número de especies para un mismo esfuerzo de muestreo; esto indica que con menor esfuerzo (número de individuos capturados) las trampas Malaise rinden más en número de especies. Sin embargo ambos métodos resultaron ser altamente complementarios, ya que la composición de especies de las muestras obtenidas por ambos métodos difirió. Esto resalta la utilidad de implementar distintos métodos de muestreo, si lo que se busca es caracterizar una comunidad de manera completa.
En este trabajo sólo se analizó la eficiencia de dos tipos de trampas, aunque existe diversidad de métodos de captura, tendientes a obtener especies con distintos hábitos. Probablemente, con la utilización de otros métodos de captura en la misma área de estudio, se podrían obtener más especies como por ejemplo especies que vivan en troncos de árboles.
Los resultados de estudios particulares no pueden ser generalizados inmediatamente a otros taxones, métodos de captura, tipos de hábitat u otras escalas espaciales. Por ejemplo, a diferencia de las conclusiones de este trabajo, Parr & Chown (2001) encontraron que usar simultáneamente dos tipos de trampas (caída y Winkler), para muestrear hormigas en la sabana africana es ineficiente, ya que las trampas de caída son más eficaces, productivas, y consistentes. Sin embargo, estos autores destacan que en el hábitat de bosque la eficiencia del método Winkler es muy superior, y puede reemplazar totalmente las trampas de caída. Concluyen que la eficiencia relativa de estos métodos varía con la complejidad y heterogeneidad del hábitat.
Por otro lado, la eficiencia de un determinado método puede variar, esto depende de las condiciones climáticas, ya sean regionales o locales. Longino & Colwell (1997), usaron trampas Berlese para capturar hormigas tropicales y concluyeron que no es eficiente distribuir los muestreos a lo largo del año debido a la estabilidad climática, y por ende la estabilidad de las comunidades. Contrariamente, Leponce et al . (2004) analizó la eficiencia de un protocolo estándar para muestrear hormigas y concluye que las variaciones de la temperatura ambiente pueden afectar considerablemente la eficiencia de dicho protocolo. También encontraron variaciones en la eficiencia de un muestreo de hormigas en un bosque subtropical seco, al tener en cuenta componentes espaciales como la extensión y el grano del muestreo. Finalmente, dependiendo del grupo taxonómico analizado, la eficiencia de un muestreo puede variar.
Trabajando en el noroeste de la Patagonia , Sackmann et al . (2006) concluyeron que la eficiencia de un muestreo no es la misma para coleópteros que para hormigas, en cuanto a la manera de repartir el esfuerzo del mismo, entre diversos hábitats de un gradiente ambiental que incluye áreas disturbadas y no disturbadas por incendios.
Debido a que la estimación de la diversidad de artrópodos es una tarea difícil, es necesario aplicar métodos de muestreo que sean rigurosos, prácticos y eficientes. La utilización de muestreos sistemáticos permite, mediante el uso de herramientas estadísticas apropiadas, la evaluación de la eficiencia de los mismos. Teniendo en cuenta el objetivo de un estudio, es posible decidir cuándo el esfuerzo de un muestreo fue suficiente o no, y decidir la distribución de dicho esfuerzo en el espacio y en el tiempo. Además la aplicación de muestreos sistemáticos permite obtener estimaciones de riqueza para una determinada área de estudio, comparables con las resultantes de estudios realizados en otras áreas, y así describir patrones más generales de variación de la diversidad (Fisher, 1999).
APÉNDICE 1 . Especies y morfoespecies capturadas en la Reserva Natural Loma del Medio, El Bolsón, Río Negro, en los meses de enero, febrero, marzo y abril, entre los años 2002 y 2004. Observaciones: NR = nuevo registro para Argentina.

AGRADECIMIENTOS
A Sergio Roig-Juñent, Gustavo Flores, Federico Ocampo, Vladimir Gusarov, Paula Posadas y Mario Elgueta por la determinación de especies. A Juan Corley por la lectura crítica del manuscrito y dos árbitros anónimos por los valiosos aportes realizados. Este trabajo fue financiado en parte por el PID 391/03 (BID 1728/OC-AR) y PIP-CONICET 2000. PS es becaria post-doctoral de CONICET.
BIBLIOGRAFÍA CITADA
1. BORROR, D. J., C. A. TRIPLEHORN & N. F. JOHNSON. 1992. An Introduction to the Study of Insects. Saunders College, Orlando. [ Links ]
2. CABRERA, A. L. & A. WILLINK. 1973. Biogeografía de América Latina. Secretaría General de la Organización de los Estados Americanos, Washington. [ Links ]
3. CLARKE, K. R. & R. N. GORLEY. 2001. PRIMER v5: User Manual Tutorial, Primer-e, Plymouth Marine Laboratory, Plymouth, UK. [ Links ]
4. COLWELL, R. K. 2000. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User's Guide), Version 6. http://viceroy.eeb.uconn.edu/estimates [ Links ]
5. COLWELL, R. K. & J. A. CODDINGTON. 1994. Estimating terrestrial biodiversity through extrapolation. Philosophical Transactions of the Royal Society B: Biological Sciences 345: 101-118. [ Links ]
6. COOK, J. L. 2003. Conservation of biodiversity in an area impacted by the red fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Biodiversity and Conservation 12: 187-195. [ Links ]
7. DIMITRI, M. J. 1974. Pequeña flora ilustrada de los Parques Nacionales Andino-Patagónicos. Anales de Parques Nacionales . Tomo XIII: 1-122. [ Links ]
8. ERWIN, T. 1991. How many species are there? Revisited. Conservation Biology 5: 330-333. [ Links ]
9. EUBANKS, M. D. 2001. Estimates of direct and indirect effects of red imported fire ants on biological control in field crops. Biological Control 21: 35-43. [ Links ]
10. FISHER, B. L. 1999. Improving inventory efficiency: a case study of leaf-litter ant diversity in Madagascar. Ecological Applications 9: 714-731. [ Links ]
11. FOLGARAIT, P J. 1998. Ant biodiversity and its relationship to ecosystem functioning: a review. Biodiversity and Conservation 7:1221-1244. [ Links ]
12. GASTON, K. 1991. The magnitude of global insect species richness. Conservation Biology 5 238-296. [ Links ]
13. GOTELLI, N. J. & R. K. COLWELL. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters 4: 379-391. [ Links ]
14. GROOMBRIDGE, B. 1992. Global Biodiversity, Status of the Earth's Living Resources. Chapman & Hall. Londres. [ Links ]
15. HAMMOND, P. M. 1992. Species inventory. En: B. Groombridge (ed.), Global Biodiversity, Status of the Earth's Living Resources. Chapman & Hall. Londres, pp. 17-39. [ Links ]
16. JONES, C. G., J. H. LAWTON & M. SHACHAK. 1994. Organisms and ecosystem engineers. Oikos 69: 373-386. [ Links ]
17. KIM, K. C. 1993. Biodiversity, conservation and inventory; why insects matter. Biodiversity and Conservation 2: 191-214. [ Links ]
18. KOIVULA, M., P. PUNTTILA, Y. HAILA & J. K. NIEMELÄ. 1999. Leaf litter and small-scale distribution of carabid beetles (Coleoptera, Carabidae) in the boreal forest. Ecography 22: 424-435. [ Links ]
19. LEPONCE, M., L. THEUNIS, H. C. DELABIE, & Y. ROISIN. 2004. Scale dependence of diversity measures in a leaf-litter ant assemblage. Ecography 27: 253-267. [ Links ]
20. LONGINO, J. T. & R. K. COLWELL. 1997. Biodiversity assessment using structured inventory: capturing the ant fauna of a tropical rain forest. Ecological Applications 7: 1263-1277. [ Links ]
21. LONGINO, J. T., J. CODDINGTON & R. COLWELL. 2002. The ant fauna of a tropical rain forest: estimating species richness in three different ways. Ecology 83: 689-702. [ Links ]
22. MC GEOCH, M. A. 1998. The selection testing and application of terrestrial insects as bioindicators. Biological Reviews 73: 181-210. [ Links ]
23. MORRONE, J. J & S. COSCARÓN (Directores). 1998. Biodiversidad de Artrópodos Argentinos: una perspectiva biotaxonómica. Ediciones Sur, La Plata , Argentina. [ Links ]
24. OLIVIER, I. & A. J. BEATTIE. 1996. Designing a cost-effective invertebrate survey: a test of methods for rapid assessment of biodiversity. Ecological Applications 6: 594-607. [ Links ]
25. OLSON, D. M. & E. DINERSTEIN. 2002. The Global 200: priority ecoregions for global conservation. Annals of the Missouri Botanical Garden 89: 199-244. [ Links ]
26. PARR, C. & S. L. CHOWN. 2001. Inventory and bioindicator sampling: Testing pitfall and Winkler methods with ants in a South African savanna. Journal of Insect Conservation 5: 27-36. [ Links ]
27. SACKMANN, P., A. RUGGIERO, M. KUN & A. FARJI-BRENER. 2006. Efficiency of a rapid assessment of the diversity of ground beetles ad ants, in natural ad disturbed habitats of the Nahuel Huapi region (NW Patagonia, Argentina). Biodiversity and Conservation 15: 2061-2084. [ Links ]
28. SCUDDER, G. G. E. 2000. Pitfall trapping. Ecological Monitoring and Assessment Network Coordinating Office, Ecosystem Science Directorate of Environment Canada. http://eqb-dqe.cciw.ca/eman [ Links ]
29. SPENCE, J. R. & J. K. NIEMELÄ. 1994. Sampling carabid assemblages with pitfall traps: the madness and the method. Canadian Entomologist 126: 881-894. [ Links ]
30. TOWNES, H. 1972. A light-weight Malaise trap. Entomological News 83: 239-247. [ Links ]
31. WILSON, E. O. 1987. The little things that run the world (the importante and conservation of invertebrates). Conservation Biology 1:344-346. [ Links ]
32. WILSON, E. O. 1992. The diversity of life. Cambridge, Massachusetts, Belknap Press. [ Links ]
Recibido: 29-V-2006
Aceptado: 7-VIII-2006

