










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. v.67 n.1-2 Mendoza ene./jun. 2008
Aphididae (Hemiptera) on ornamental plants in Córdoba (Argentina)
Aphididae (Hemiptera) en plantas ornamentales de Córdoba (Argentina)
Szpeiner, Alfonsina
Instituto de Investigaciones Fisiológicas y Ecológicas Vinculadas a la Agricultura (IFEVA)-CONICET, Facultad de Agronomía, Universidad de Buenos Aires. Avenida San Martín 4453, Capital Federal, C1417DSE, Argentina; e-mail: szpeiner@agro.uba.ar
ABSTRACT. In 30 surveys across Cordoba city and surroundings (Argentina) I recorded, from November 2001 to May 2002, a total of 109 ornamental plants infested with aphid colonies. Surveys were conducted in the local botanical garden as well as homestead gardens and nurseries. Seventeen species of aphids were identified, 4 of which are new species in the area, from 47 ornamental plants. In a total of 59 plant/aphid associations, 38 new associations were found. I present the first record of aphids colonizing Cactaceae in Argentina. Most aphid colonies were medium and a high proportion (more than 40%) showed alate individuals. The most frequent aphid species, Aphis gossypii Glover and Aphis craccivora Koch were especially associated with the most frequent damages: curled leaves or buds and necrosis, respectively. More than 30% of aphid colonies were attended by ants. Camponotus Mayr and Prenolepis Mayr were the ant genus attending the highest number of aphid species.
KEYWORDS. Aphids; Ornamental plants; Richness; Damages; Urban areas.
RESUMEN. En 30 censos realizados en la ciudad de Córdoba y alrededores (Argentina) se registraron, entre noviembre de 2001 y mayo de 2002, un total de 109 plantas ornamentales infestadas de colonias de áfidos. Los censos fueron realizados en jardines, viveros y el jardín botánico de la ciudad. Se identificaron 47 especies de plantas ornamentales infestadas con 17 especies de áfidos (4 nuevas en el área). Se establecen 59 relaciones áfido/planta, de las cuales 38 son nuevas. Se obtuvo el primer registro de áfidos que colonizan Cactaceae en Argentina. La mayoría de las colonias de áfidos fueron medianas y una alta proporción (más del 40 %) mostró individuos alados. Aphis gossypii Glover y Aphis craccivora Koch, fueron las especies más frecuentes y generaban los daños más conspicuos, como el enrollamiento y la necrosis foliar. Más del 30% de las colonias estuvieron atendidas por hormigas, siendo Camponotus Mayr y Prenolepis Mayr los géneros de hormigas que frecuentaban el mayor número de especies de áfidos.
PALABRAS CLAVE. Áfidos; Plantas ornamentales; Riqueza; Daños; Áreas urbanas.
INTRODUCTION
Urban landscapes are becoming widespread, with increasing number of overpopulated cities (United Nations, 2001; Wilby & Perry, 2006). The environment created around human settlements is very dynamic, complex and in some cases highly polluted, with a continuous introduction of exotic plant and animal species (Vitousek et al., 1997; Nieto Nafría et al., 1994). For example, in Argentina over 75% of aphids are exotic (Ortego et al., 2006).
Aphids (Hemiptera, Aphididae) are one of the major pests in ornamental plants (Arruda et al., 1996; Clements et al., 2000). However, a few biological studies on aphids have been made in South America (Dixon, 1998). For example, Peronti & Sousa da Silva (2002) surveyed the aphid richness in ornamental plants from Sao Pablo, Brazil. The arthropod communities in different areas of Argentina have been described, but records of arthropod communities in city gardens are almost non-existing.
Aphids are small, soft-bodied and plant sucking insects. Aphids' life cycles are very intricate and can imply both sexual and asexual reproduction as alate and apterous forms (Nieto Nafría et al., 1994). However, the loss of the sexual phase (or anholocycly) is a common phenomenon (Blackman & Eastop, 2000). A typical aphid colony consists of apterous viviparous females (apterous virginoparae) and, as the colony ages, alate viviparous females (alate virginoparae) appear.
In temperate regions, aphids are the most important honeydew producers, many of which have developed a mutualism with ants (Hölldobler & Wilson, 1990). Aphids benefit from this mutualism with ants by reduced predation and parasitism, and by reduced risk of fungal infection (Völkl et al., 1999). This aphid-ant mutualism may result in differences in aphid colony size (Addicott, 1978; Delfino & Buffa, 2000).
Aphids can be found virtually in all temperate cropping systems and have the potential to reduce plant yield substantially (Sadras et al., 1999). Those insects often reduce plant vigor, promote curled and distorted leaves, harden the buds and cause chlorosis as well as promoting malformations and heterogeneous coloration in organs like leaves, flowers and fruits (Sanchez et al., 2000). These damages produce important losses in the commercial value of ornamental plants which, in contrast to other crops, are usually related to aesthetics.
The aim of this work is to identify and characterize aphid colonies present in the most frequently cultivated ornamental plant species in gardens of Córdoba city (Argentina) and surroundings.
MATERIAL AND METHODS
Area description. Córdoba city (31°24'S, 64°11'W) is placed in the center of Argentina in a sub humid region. Rainfalls are mainly concentrated in summer and mean annual temperature is 17.4 °C. Surveys reported here were carried out in this city and nearby surroundings (i.e. Villa Giardino).
Sampling procedure. To collect aphid colonies, 30 surveys were conducted from November 2001 to May 2002. Surveys consisted of random tours along the whole Córdoba's botanical garden as well as several nurseries and homestead gardens in the study area. Aphids were maintained in test tubes filled with 65% alcohol. Only ornamental plants with aphids' colonies were recorded. The following variables were considered:
(a) Size of the colony: (1) small colonies: up to ten aphids per colony, (2) medium colonies: 10-50 aphids per colony or (3) large colonies: more than 50 aphids per colony.
(b) Aphid forms: (1) apterous or (2) alate virginoparae aphids.
(c) Infested plant organs: (1) buds, (2) leaves, (3) stems, (4) flowers and/or (5) fruits.
(d) Presence of damages: (1) stains, (2) necrosis, (3) curled leaves or buds, (4) chlorosis, (5) fading, (6) reduced plant vigor and/or (7) honeydew deposit.
(e) Functional plant forms: (1) herbs, (2) shrubs, (3) trees or (4) cactus/succulent plants.
(f) Ant attendance: (1) Yes, (2) No.
Species identification. Except for a few host plants that were herborized and identified at the laboratory, most determinations were carried out during the samplings. Both, aphid species and ant genus were identified under magnifying glass. When it was required, aphids were mounted on slides according to the technique described by Remaudière (1992), and species identification was done under optical microscope up to species level, using the following identification keys: Blackman & Eastop (2000); Delfino (1981); Heie (1980, 1982, 1986, 1992, 1994, 1995) and Hille Ris Lambers (1939, 1947,1953). The collected aphids and ants were placed in the Insect Collection of the Entomology Laboratory at National University of Cordoba (UNC).
Furthermore, aphids were classified according to their apterous virginoparae known host plant specificity based on Kennedy et al. (1962): (1) monophagous: colonize a plant genus; (2) oligophagous: colonize up to five plant genus and belong to no more than 20 plant families; (3) polyphagous: colonize more than 5 plant genus and belong to more than 20 families. Finally, aphid species were grouped based on Mondor et al. (2006) as: (1) strictly monoecious (i.e. no host alternation); (2) monoecious/heteroecious populations known; (3) strictly heteroecious (i.e. obligate host alternation).
RESULTS
Richness of aphids and their ornamental host plants
I recorded 109 ornamental plants with aphid in 30 surveys along the botanical garden, several nurseries and homestead gardens in Córdoba and surroundings from November 2001 to May 2002. In 47 different ornamental plants belonging to 13 plant families, I identified 17 species of aphids (Table I). From these aphid species, I found four new species in the area: Cryptomyzus korschelti Börner, Tinocallis kahawaluokalani (Kirkaldy), Tuberculatus querceus (Kaltenbach) and Uroleucon ambrosiae (Thomas). The finding of Cryptomyzus korshelti Börn was previously reported (Szpeiner & Delfino, 2002).
TABLE I. Aphids and host ornamental plants collected in Cordoba (Argentina).

Aphis gossypii Glover and Aphis craccivora Koch were the most frequent aphid species found. Whereas A. gossypii colonized 21 ornamental host plants, A. craccivora only colonized 11. Most aphid species identified in this study were exotic. Uroleucon ambrosiae (Thomas) was the only species with an exclusively American distribution (Nieto Nafría et al., 1994). In other words, 94% (1/17) of aphids colonizing ornamentals in Córdoba were cosmopolitan or exotic species.
With regard to ornamental plants, Rosa L. was the most frequent genus of plants colonized by aphids (data not shown). Rosaceae were the ornamental host plant family with the greatest number of plant species registered, and Cactaceae, with five ornamental host plant species, was a new aphids' host plant family in Argentina. Furthermore, I found 38 new associations in Argentina, from 59 associations between a particular plant species and a particular aphid species recorded (Table I). Among these plant/ aphid associations, the most frequently observed was the association between Rosa sp. and Wahlgreniella nervata (Gillette). This association was found in a 4.6% of the total samples recorded.
Aphid colonies characteristics
Most aphid colonies were medium size (10-50 aphids per colony) of polyphagous aphid species, and a high proportion (more than 40%) of colonies had alate aphid forms (Fig. 1). Aphis craccivora showed the highest number of large colonies, whereas Macrosiphonella sanborni (Gillette) and Rodobium porosum (Sanderson) never showed colonies with alate individuals during the study (data not shown). In terms of host aphid specificity, in the 17 aphid species, we found five monophagous, six oligophagous and six polyphagous species colonizing ornamentals in the area (Table I). Finally, most aphid species were strictly monoecious (59%), whereas C. korshelti was the only species strictly heteroecious (Table I).
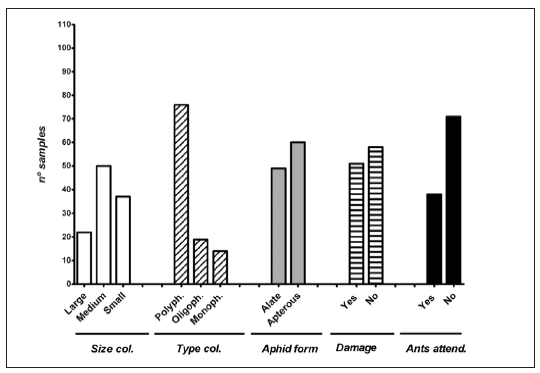
Fig. 1. Number of samples (n =109) for some variables that characterize Aphididae (Hemiptera) colonization in ornamental plants. Size col. = Size of the colony; Polyph. = Polyphagous; Oligoph. = Oligophagous; Ants Attend. =Ant attendance.
Ant attendance
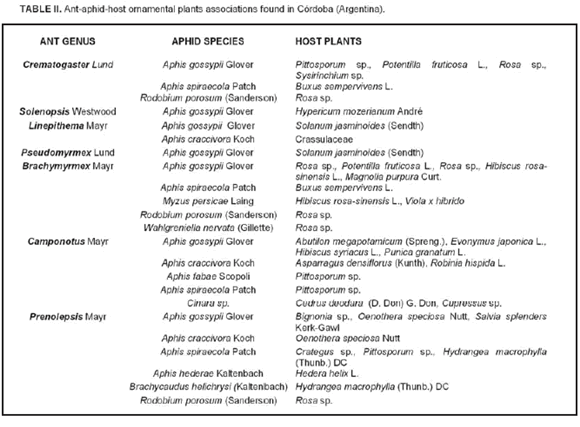
Ants attended more than 30% of aphid colonies (Fig. 1). Seven ant genus, nine aphid species and twenty-seven ornamental host plant species produced forty-one ant-aphid associations in the study area (Table II). Aphid species attended by ants were Aphis craccivora, A. fabae Scolopoli, A. gossypii, A. hederae Kaltenbach, A. spiraecola Patch, Brachycaudus helichrisy (Kaltenbach), Cinara sp., Myzus persicae (Sulzer), and Wahlgreniella nervata (Gillette). Aphis gossypii was attended by all ant genus recorded in the study area. Camponotus Mayr and Prenolepis Mayr were the ant attendant genus that attended the highest number of aphid species. Most aphid colonies attended by ants were medium or large size (data not shown).
Aphid damages in ornamental plants
In 109 samples, more than 40% of ornamental host plant families showed damages (Fig. 1). Leaves and buds were the most commonly infested plant organs followed by flowers (data not shown). Aphis gossypii and A. craccivora, the most frequent aphid species, were especially associated with the most frequent damages: curled leaves or buds and necrosis, respectively (Fig. 2). M. sanborni and M. persicae were only associated with necrosis, whereas Cinara sp. was the only one that produced outstanding honeydew deposits.

Fig. 2. Aphid damages in ornamental plants. Columns show relative percentages of host plant species with different damages found in the study. Curled L/B= Curled leaves or buds, Honeydew= honeydew deposits, reduce vigor= reduce plant vigor.
DISCUSSION
This study confirmed that 13 out of 72 aphid species recorded in Córdoba (Argentina) by Nieto Nafría et al. (1994) colonize ornamental plants. Moreover, these surveys added four aphid species new in the area, one of them, Cryptomyzus korshelti Börn, was previously reported (Szpeiner & Delfino, 2002). The importance of the presence of C. korshelti in Argentina was related with this species range distribution, until now exclusively European (Heie, 1994).
For Tinocallis (Sarucallis) kahawaluokalani (Kirkaldy) and Tuberculatus querceus (Kaltenbach), their presence in Córdoba expanded their distribution along Argentina. La Rossa et al. (1997) reported the first finding of Tinocallis (Sarucallis) kahawaluokalani (Kirkaldy) in Argentina but limited to Buenos Aires. On the other hand, Nieto Nafría et al. (1994) mentioned the first occurrence in South America of Tuberculatus querceus (Kaltenbach) in Santa Fe (Argentina). Essig (1953) mentioned for the first time in Argentina (Salta) the fourth new species, Uroleucon ambrosiae (Thomas), colonizing Artimisia sp. However, Nieto Nafría et al. (1994) affirmed this finding could be accidental because Uroleucon ambrosiae (Thomas) only colonize composite plants. My report of U. ambrosiae (Thomas) in Córdoba colonizing two composite plant species (Euryops sp. and Gerbera jamesonii Bolus) confirmed Essig's (1953) finding but also it expanded this species distribution to Córdoba and surroundings. For more details about this genus in South America refer to (De Carvalho et al., 1998; Nieto Nafría et al., 1994)
Aphis gossypii and A. craccivora were the most frequent aphids colonizing ornamental plants. These species are cosmopolitan and very important agricultural pests, with very wide host ranges (Blackman & Eastop, 2000). Nonetheless, ornamentals are generally exotic, and host of exotic aphid species were most probably introduced together from holartic populations (Ortego et al., 2004).
The plant family with a greater number of species colonized by aphids was Rosaceae, which agrees with Dixon (1998) who mentions Rosaceae as the second most popular aphids' plant hosts group in the world, after Asteraceae. However, the reduced relevance of Asteraceae in this study may be related to restricted range of ornamentals that occurred in the surveyed area.
An interesting finding was that for the first time in Argentina I recorded Cactaceae as host of two exotic Aphididae species, Aphis gossypii and A. craccivora. However, these plants were growing outside their natural ecological conditions and exposed to the gardens´ fertile soils and higher water availability, which may have altered the plant's physiology, inducing the colonization. Food quality is known as an important feature in aphids' host selection (Dixon 1970, 1998).
Self-regulation allows aphid colonies to maintain aphid individual number according to available food resources (Shaposhnikov, 1989). This self-regulation could be the ecological mechanism behind the maintenance of most colonies found in ornamentals with a medium size. This idea is also supported by the frequent presence of alate individuals found. The production of alate individuals, which is a response to overcrowding or shortage of food resources, is the main mechanism in self-regulation of aphid colonies (Dixon, 1998)
In the study area, as expected according to soluble nitrogen level, aphids infested mainly buds, leaves and flowers (Dixon, 1998). However, most aphids found were oligophagous or polyphagous, which disagrees with a previous study in Sao Carlos, Brazil (Peronti & Sousa da Silva, 2002), where monophagy prevailed as the most common pattern of host specificity. It is not clear from our study if this difference was caused by the ecological conditions affecting overall aphid diversity in each geographical region, or if it is just a sampling effect.
Most aphid species on ornamentals in Córdoba were strictly monoecious. This result agrees with some authors (i.e. Dixon, 1998; Mondor et al., 2006) that assert monoecious aphid species must be better able to survive in new environments. Ornamental plants may be considering «new environments» as a result of, as I discussed above, their exotic origin.
The high percentage of aphid colonies attended by aphids (more than 30%) showed the relevance of these interactions on ornamentals in the study area. According to Delfino & Buffa (2000), ant attendance favors aphids' host plant colonization, strengthening the colonies by allowing a higher number of individuals per colony. Most aphid colonies on ornamentals were medium or large. A. gossypii, the aphid species that colonized most ornamental host plant species, was attended by a high number of different ant genera. It is well documented that aphid species are associated with several ant genera that may vary according to host plants distributions irrespectively of aphid host plant specificity (Delfino & Buffa, 1996). In agreement with Delfino & Buffa (1996), Camponotus was the ant genus associated with the highest number of aphid species even when Prenolepis attended the same number of species.
It is known that, in ornamentals, aesthetic damage has low tolerance (Krips et al., 1998). The results presented here showed that most ornamental plants had some direct damage when they are infested by aphids, warning about the relevance of Aphididae as pest in ornamental plants in the study area. In addition, curled leaves or buds and necrosis were the most common damage, which suggests virus diseases associated with the aphid infection (Miles, 1989). Honeydew deposits, which produce a significant reduction in light use efficiency and maximum rate of assimilation in leaves promoting leave aging (Wellings, 1989) were also observed. However, in the study area, this type of damage was less frequent than direct damage and closely related to Cinara sp. and, therefore, associated with Coniferae.
Final remarks
This work provides evidence that a high percentage of aphid fauna cited in Argentina colonize and cause damages in ornamental plants in urban habitats. Further studies will help us to confirm aphid species distribution, to assess ornamental host plant range as well as to find ecological patterns that allow us to characterize more completely aphid infestation on ornamentals. This information on aphids and other arthropod species is important in the context of continuous urban expansion.
ACKNOWLEDGEMENTS
I gratefully acknowledge Biól. Liliana Buffa for the identification of ant specimens. Furthermore, I really appreciate Dr. Miguel Angel Delfino's guidance in aphid specimen's identification. I also thank the Cordoba's botanical garden authorities, who allowed me to collect aphid samples from its collection. Finally, I would like to thank Dr. María Alejandra Martínez-Ghersa for her helpful comments on the manuscript.
LITERATURE CITED
1. ADDICOTT, J. H. 1978. Competition for mutualists: aphids and ants. Can. J. Zool. 56: 2093-2096.
2. ARRUDA, S. T., M. P. OLIVETTE & C. E. CASTRO. 1996. Diagnostico da floricultura do Estado de Sao Paulo. Rev. Bras. Hort. Ornam. 2: 1-18.
3. BLACKMAN, R. L. & V. F. EASTOP. 2000. Aphids on the World's Crops: An Identification and Information Guide. John Wiley, Chichester.
4. CLEMENTS, K. M., C. E. SORENSON, B. M. WIEGMANN & M. ROE. 2000. Insecticide resistence in the Myzus persicae complex (Homoptera-Aphididae) with emphasis on tobacco pest management. Rev. Toxicol. 3: 1-23.
5. DE CARVALHO, R. C. Z.., R. BLACKMAN & J. M. SPENCE.. 1998. The genus Uroleucon Mordvilko (Insecta, Aphididae) in South America, with a key. Zool. J. Linn. Soc. 123:117-141.
6. DELFINO, M. A. Inéd. Aphididae (Homoptera) de la provincia de Córdoba. Tesis, Universidad Nacional de Córdoba, Córdoba, 1982, 195 pp.
7. DELFINO, M. A. 1981. Una nueva especie del género Wahlgreniella H. L. R. 1949 (Hom. Aphididae). Rev. Soc. Entomol. Argent. 41: 111-113.
8. DELFINO, M. A. & L. M. BUFFA. 1996. Asociaciones hormigas-planta en la Argentina. Rev. Per. Ent. 39: 81-84.
9. DELFINO, M. A. & L. M. BUFFA. 2000. Algunas interacciones planta-hormiga en Córdoba (Argentina). Zool. Baetica 11: 3-15.
10. DELFINO, M. A. & A. BINAZZI. 2002. Áfidos de Coníferas en la Argentina (Hemiptera: Aphididae). Rev. Soc. Entomol. Argent. 61(3-4): 27-36.
11. DIXON, A. F. G. 1970. Quality and availability of food for sycamore aphid population. In: Waton, A. (ed.), Animal Populations in Relation to their Food Resources, Blackwell, Oxford, pp. 271-287.
12. DIXON, A. F. G. 1998. Aphid Ecology. Chapman & Hall, New York.
13. ESSIG, E. O. 1953. Some new and noteworthy Aphididae from western and southern South America. Proc. Calif. Acad. Sci. 28: 59-164.
14. HEIE, O. E. 1980. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica. Scandinavian Science Press, Leiden- Copenhagen.
15. HEIE, O. E. 1982. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica II. Scandinavian Science Press, Leiden- Copenhagen.
16. HEIE, O. E. 1986. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica III. Scandinavian Science Press, Leiden- Copenhagen.
17. HEIE, O. E. 1992. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica IV. Scandinavian Science Press, Leiden- Copenhagen.
18. HEIE, O. E. 1994. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica V. Scandinavian Science Press, Leiden- Copenhagen.
19. HEIE, O. E. 1995. The Aphidoidea (Hemiptera) of Fennoscandia and Denmark: Fauna Entomologica Scandinavica VI. Scandinavian Science Press, Leiden-Copenhagen.
20. HILLE RIS LAMBERS, D. 1939. Contributions to a monograph of the Aphididae of Europe II. Temminkia 4: 1-134.
21. HILLE RIS LAMBERS, D. 1947. Contributions to a monograph of the Aphididae of Europe III. Temminkia 7: 179-319.
22. HILLE RIS LAMBERS, D. 1953. Contributions to a monograph of the Aphididae of Europe V. Temminkia 11: 1-176.
23. HÖLLDOBLER, B. & E. WILSON. 1990. The ants. Springer, Berlin - New York.
24. KENNEDY J. S., M. F. DAY & V. F. EASTOP. 1962. A Conspectus of aphids as vector of plants viruses. Commonwealth Institute of Entomology, London.
25. KRIPS, O. E., A. WITUL, P. E. L. WILLEMS & M. DICKE. 1998. Intrinsic rate of population increase of the spider mite Tetranychus urticae on the ornamental crop Gerbera: intraspecific variation in host plant and herbivore. Entomol. Exper. Applic. 89: 159-168.
26. LA ROSSA, F. R., A. L. VASICEK & A. NORIEGA. 1997. Presencia de Sarucallis kahawaluokalani sobre «crespón» (Lagerstroemia indica) en la Argentina (Homoptera: Aphidoidea). Rev. Soc. Entomol. Argent. 56: 97-100.
27. MILES, P. W. 1989. Specific responses and damage caused by Aphidoidea. In: Minks, A. K. & P. Harrewijn (eds.), Aphids, their Biology, Natural Enemies and Control, Elsevier, Amsterdam, pp. 255-268.
28. MONDOR E. B.., E. TREMBLAY & R. H. MESSIG. 2006. Morphological and ecological traits promoting aphid colonization of the Hawaiian Islands. Biol. Invasions DOI 10.1007/s10530-006-9010-z.
29. NIETO NAFRÍA, J. M., M. A. DELFINO & M. P. MIER DURANTE. 1994. La Afidofauna de la Argentina. Su conocimiento en 1992. Universidad de León, León, España.
30. ORTEGO J., M. E. DIFABIO & M. P. MIER DURANTE. 2004. Nuevos registros y actualización de la lista faunística de los pulgones (Hemiptera-Aphididae) de la Argentina. Rev. Soc. Entomol. Argent. 63: 13-30.
31. ORTEGO J., N. PERÉZ HIDALGO, J. M. NIETO NAFRÍA & M. P. MIER DURANTE. 2006. Six alien aphid species (Hemiptera: Aphididae) recorded for the first time from South America. Fla. Entomol. 89: 111-116.
32. PERONTI, A. L. B. G. & C. R. SOUSA DA SILVA. 2002. Aphids (Hemiptera: Aphidoidea) of ornamentals plants from Säo Carlos, Säo Paulo State, Brazil. Rev. Biol. Trop. 50 (1): 137-144.
33. REMAUDIÈRE, G. 1992. Une Méthode simplifiée de montage des aphides et autres petits insectes dans le baume du canada. Rev. Fr. Entomol. 14(4): 185-186.
34. SADRAS, V. O., A. FERERES & R. H. RATCLIFFE. 1999. Wheat Growth, Yield, and Quality as affect by insect herbivores. In: Satorre, E. H. & G. A. Slafer, (eds.), Wheat: Ecology and physiology of yield determination, The Haworth, New York, pp. 183-227.
35. SANCHEZ, M. C., M. CERMELIO, W. MACHADO, F. CENTENO & E. BROWN. 2000. Diversidad de áfidos (Homoptera: Aphididae) capturados con trampas amarillas en el cultivo de pimentón (Capsicum annuum L.) y su relación con los factores climáticos. Bol. Entomol. Venez. 15 (1): 61-83.
36. SHAPOSHNIKOV, G. C. 1989. Organization (structure) of populations and species, and speciation. In: Minks, A. K. & P. Harrewijn (eds.), Aphids, their Biology, Natural Enemies and Control, Elsevier, Amsterdam, pp. 415-430.
37. SZPEINER, A. & M. A. DELFINO. 2002. Cryptomyzus korschelti Börner (Hemiptera, Aphididae) nuevo áfido para la fauna argentina. In: XI Jornadas Fitosanitarias Argentinas, Río Cuarto, Córdoba, Argentina, pp. 127.
38. UNITED NATIONS. 2001. World urbanization prospects: the 2001 revision. United Nations, Department of Social and Economic Affairs.
39. VITOUSEK, P. M.., H. A. MOONEY, J. LUBCHENCO & J. M. MELILLO. 1997. Human domination of earth's ecosystems. Science 277: 494-499.
40. VÖLKL W., J. WOODRING, M. FISHER, M. W. LORENZ & K. H. HOFFMANN. 1999. Ant-aphid mutualisms: the impact of honeydew production and honeydew sugar composition on ant preferences. Oecologia 118: 483-491.
41. WELLINGS P. C., S. A. WARD, A. F. G. DIXON & R. RABBINGE. 1989. Crop Loss Assessment. In: Minks, A. K. & P. Harrewijn (eds.), Aphids, their Biology, Natural Enemies and Control, Elsevier, Amsterdam, pp. 49-64.
42. WILBY, R. L. & G. L., PERRY. 2006. Climate change, biodiversity and the urban environment: a critical review based on London, U..K. Prog. Phys. Geog. 30(1): 73-98. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 12-12-2006;
Aceptado: 6-11-2007

