










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. v.67 n.1-2 Mendoza ene./jun. 2008
Revalidation of Argentagrion and redefinition of Homeoura, with the description of H. obrieni n. sp. (Odonata: Coenagrionidae)
Revalidación de Argentagrion y redefinición de Homeoura, con la descripción de H. obrieni n. sp. (Odonata: Coenagrionidae)
Von Ellenrieder, Natalia
Instituto de Bio y Geo Ciencias (IBiGeo), Museo de Ciencias Naturales, Universidad Nacional de Salta, Mendoza 2, 4400 Salta, Argentina; e-mail: natalia.ellenrieder@gmail.com
ABSTRACT. Argentagrion Fraser, currently considered a synonym of Homeoura Kennedy, is revalidated; both genera are rediagnosed and their species illustrated, keyed and mapped. Ischnura sobrina Schmidt is transferred to Homeoura, and a new species, H. obrieni is described, resulting in five species being included in Homeoura (H. chelifera (Selys), H. lindneri (Ris), H. nepos (Selys), H. obrieni sp. nov. and H. sobrina (Schmidt) comb. nov.) and two in Argentagrion (A. ambiguum (Ris) and A. silviae Bulla).
KEY WORDS. Odonata; Coenagrionidae; Argentagrion; Homeoura ; South America.
RESUMEN. Se revalida Argentagrion Fraser, considerado actualmente como un sinónimo de Homeoura Kennedy; ambos géneros son rediagnosticados y se proporcionan ilustraciones, claves y mapas para sus especies. Se considera que Ischnura sobrina Schmidt pertenece a Homeoura y una nueva especie, H. obrieni, es descripta, resultando un total de cinco especies incluidas en Homeoura (H. chelifera (Selys), H. lindneri (Ris), H. nepos (Selys), H. obrieni sp. nov. y H. sobrina (Schmidt) comb. nov.) y dos en Argentagrion (A. ambiguum (Ris) y A. silviae Bulla).
PALABRAS CLAVE. Odonata; Coenagrionidae; Argentagrion; Homeoura; América del Sur.
INTRODUCTION
Generic concepts within Neotropical Coenagrionidae are still not fully resolved, with poorly defined and/or artificial genera (May, 2002; De Marmels & Garrison, 2005; De Marmels 2007; von Ellenrieder & Lozano, 2008). In this paper I revise the genus Homeoura Kennedy, 1920. According to recent usage (Bridges, 1994; Muzón & von Ellenrieder, 1998; Costa et al. 2000; Lencioni, 2006), the genus currently includes five species: H. ambigua (Ris), H. chelifera (Selys), H. lindneri (Ris), H. nepos (Selys) and H. silviae (Bulla). The history of how this genus came to be is complex and tortuous with various species being transferred from one genus to another.
Its species were originally described in the genera Acanthagrion Selys, Agrion Fabricius, Argentagrion Fraser and Ischnura Charpentier, and their generic position shifted over the years, involving the passage of some of them also through Enallagma Charpentier.
Selys (1876) described Acanthagrion? cheliferum under his 'Acanthagrion interruptum' group, with vein A1 originating anteriorly to CuA in FW and male S10 slightly elevated forming two points. He (Selys, 1876) also described Agrion? nepos based on a single male, but stated that if the then unknown female was found to have a vulvar spine, it should then be considered as an aberrant species of Ischnura. Calvert (1909) described the female, and transferred this species to Ischnura due to the presence of a vulvar spine.
Ris (1904) expressed doubts in describing Acanthagrion ambiguum as to its correct generic placement. Fraser (1948) created the genus Argentagrion for reception of this species and diagnosed it from Acanthagrion by its small size, delicate build and greatly reduced wing venation.
Kennedy (1920) created the genus Cyanallagma to include species of Selys' 'interruptum' group and "perhaps cheliferum", and Homeoura to include Agrion? nepos. Kennedy's diagnoses were extremely brief and inconclusive; Cyanallagma was characterized as «an Acanthagrion but with male cerci not decumbent and usually forked», and Homeoura as «an Ischnura but more Enallagmine, with proximal and distal sides of pterostigmata rounded, costal slightly indented at nodus and second segment of male genital ligula with large lateral patches of spines.»
Probably unaware of Kennedy's (1920) generic descriptions, Ris (1928) described Acanthagrion lindneri stating that it was similar to A. cheliferum and A. ambiguum, and Schmidt (1942) described Ischnura sobrina, noting its similarity to I. nepos.
Santos (1956) shifted Acanthagrion cheliferum to Enallagma, based on the petiolation of the wings ending proximal to anal crossing as in the latter, and the male cerci with a ventral process at base as in some species of the primarily Nearctic genus Enallagma. However, he offered no comments on the placement of the other species described under Acanthagrion which shared these characters. Rácenis (1958) disagreed with Santos (1956), noting that wing petiolation also ends proximal to anal crossing in all other species of Selys' second section of Acanthagrion, and even in some species of Acanthagrion sensu stricto. He (Rácenis, 1958) concluded that the species of Selys' second section should be placed in two different genera: Cyanallagma, with venation complete, 11-14 postnodals, male paraprocts at least as long as half of the cerci and larger size (abdomen 21.5-28; HW 16- 19.5), and Argentagrion, with venation reduced, 6-10 postnodals, male paraprocts shorter than half of cerci, and smaller size (abdomen 18-23; HW 12.5-15). Besides A. ambiguum he included A. cheliferum and A. lindneri in Argentagrion but did not mention H. nepos. The similarity of A. cheliferum with Homeoura, especially regarding genital ligula morphology, was first noted by Donnelly & Alayo (1966).
Bulla (1971b) described Argentagrion silviae, and since the generic redefinition by Rácenis (1958) for Argentagrion did not allow the inclusion of his new species, he transferred A. cheliferum and A. lindneri back to Acanthagrion «…until their placement could be reanalyzed», and restricted Argentagrion to include only A. ambiguum and A. silviae as follows: more slender species, HW of 10-12.5 mm, postocular spots triangular, very elongated transversally and partially or completely fused to pale occipital bar, posterior margin of prothorax not projected, pterostigma as long as half a cell, apical margin of male S10 with a pointed or blunt medial apophysis, ventral process of male cerci apical, represented by the ventrally bent tips of unforked cerci, and male paraprocts as long as or longer than half of cerci. Bulla (1971a) also noted the different shape of larval caudal lamellae of A. cheliferum and A. ambiguum. He (Bulla,1973) later described the similarities in male caudal appendages, female thorax and larval lamellae between Cyanallagma bonariense - C. interruptum and Acanthagrion cheliferum - A. lindneri, and suggested the possibility of placing the latter pair of species in Cyanallagma.
De Marmels (1989) redefined Cyanallagma, and diagnosed it from Argentagrion ambiguum, Acanthagrion cheliferum, Homeoura nepos and H. lindneri, and later (De Marmels, 1997) also placed Acanthagrion cheliferum in Homeoura.
Davies & Tobin (1984) listed A. ambiguum, A. silviae, A. cheliferum, A. nepos, A. lindneri and A. silviae under Argentagrion, and incorrectly treated Homeoura as its junior synonym without providing any explanation. This was followed by Steinmann (1997) who, however, incorrectly attributed the synonymy of Homeoura with Argentagrion to Fraser (1948). Following the principle of priority, Garrison (1991) had previously inverted the synonymy.
Lencioni (2006) characterized the genus Homeoura, with Argentagrion as its junior synonym, by the different size of quadrangles in FW and HW, and also small size, overall black coloration with light blue and yellow pale areas,presence of pale postocular spots, and females with a vulvar spine on S8. However, FW and HW quadrangles differ in size (costal side of FW quadrangle is shorter than half of HW quadrangle costal side) not only in Argentagrion and Homeoura, but also in Oxyallagma Kennedy, Tigriagrion Calvert and several Enallagma and Ischnura species, and the particular combination of small size and color pattern is also present in other neotropical Coenagrionidae (i.e. some species of Ischnura).
Based on morphological evidence, I contend that the species included in Homeoura belong to two separate genera (A. ambiguum and A. silviae in Argentagrion and H. cheliferum, H. nepos and H. lindneri in Homeoura); I also place Ischnura sobrina in Homeoura, describe a new species of Homeoura and provide keys, diagnoses, illustrations and distribution maps for all involved species.
MATERIAL AND METHODS
Adults of all described species of Argentagrion and Homeoura, with the sole exception of H. sobrina, the types of which are apparently lost (see remarks under this species account), and all original descriptions and revisions were analyzed. Additionally, several species of other coenagrionid genera were examined to establish generic boundaries. Acronyms used for collections are as follows:
DRP D.R. Paulson personal collection, Tacoma, Washington, USA;
FML Fundación Miguel Lillo, Tucumán, Argentina;
FNS Forschungsinstitut und Naturmuseum Senckenberg, Frankfurtam- Main, Germany;
IRSNB Institut Royal des Sciences Naturelles de Belgique, Bruxelles, Belgium;
MCZ Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA;
MLP Departamento Científico de Entomología, Museo de La Plata, Argentina;
MNRJ Departamento de Entomología, Museu Nacional, Río de Janeiro, Brazil;
RWG R.W. Garrison personal collection, Sacramento, California, USA;
SWD S.W. Dunkle personal collection, Tucson, Arizona, USA;
UMMZ University of Michigan, Museum of Zoology, Ann Arbor, Michigan, USA.
Diagnostic characters were illustrated with the aid of a camera lucida. Measurements are in mm, and total length and abdominal length do not include appendages. Wing terminology follows Riek & Kukalová-Peck (1984) and genital ligula terminology follows Kennedy (1916). Maps represent distribution records from collections and reliable literature records, and were created electronically from the Digital Chart of the World (1:1,000,000) using ArcView 9.1. Elevation data and longitude/latitude coordinates were culled
from the Global Gazetteer website (<http://www.fallingrain.com/world/> ) . Abbreviations for structures used throughout the text are as follows: FW: forewing; HW: hindwing; pt: pterostigma; Px: postnodal crossveins; S1-10: abdominal segments 1 to 10; SD: standard deviation.
RESULTS
Argentagrion Fraser, 1948
Argentagrion Fraser, 1948: 48-50 (diagnosis); Rácenis, 1958: 190-192 (generic redefinition); St. Quentin, 1960: 52, 58 (generic key, diagnosis); Donnelly & Alayo, 1966: 108-109 (comparison to Enacantha); Bulla, 1971b: 49-54 (generic redefinition); Bulla, 1973: 95 (discussion of generic placement); Davies & Tobin, 1984: 65; Bridges, 1994: III.6 (as junior synonym of Homeoura); Steinmann, 1997: 245; Tsuda, 2000: 34 (as junior synonym of Homeoura). Type species: Acanthagrion ambiguum Ris, 1904 by original designation.
Characterization. Small (HW 10.4- 15mm, abdomen 17.5-21.5mm) and largely black (brown in heterochromatic females) damselflies with pale blue to pale yellow areas. Head with rounded frons, longest at level of eyes; pale postocular spots present, transversely elongated and about twice as wide as long or wider, usually concave anteriorly (Figs. 1-2), except in some heteromorphic females whose entire posterior area of head dorsum is pale; pale occipital bar is present and usually confluent to postocular spots (except in some males of A. ambiguum in which they are adjacent but not fused). Posterior lobe of prothorax either bilobate (Figs. 21, 23, in both male and female of A. ambiguum) or with a short triangular apophysis (Fig. 22, in male of A. silviae), lacking a differentiated medial lobe; pterothorax with complete antehumeral pale stripe, and no black stripe over metapleural suture (Fig. 60); tibial spurs as long as or shorter than intervening spaces; supplementary teeth of pretarsal claws well developed. CuP reaching CuPAA (Fig. 13); petiolation ending at or proximal to CuP (variable intraspecifically); pt rhomboidal (Figs. 13, 15, 18). Male genital ligula with one latero-distal lobe on each side, lacking inner fold, with an inner process (Figs. 41, 43, ip) in the center of a transverse fold distal to flexure, and with no projections proximal to flexure; setae on sides of ligula second segment usually present. Posterior margin of male S10 entire (Figs. 65-70), usually with a medial apophysis. Distal half of male cercus bent ventrally into a triangular laminar process (Figs. 65, 67-68, 70); paraprocts short with dorsal branch rounded (in A. ambiguum, Fig. 67) or ending in a sclerotized tooth (in A. silviae, Fig. 70, to). Female vulvar spine of S8 well developed; outer valves of ovipositor with one row of spines, and extending to posterior margin of S10.

Figs. 1-12. Head, dorsal view. 1, Argentagrion ambiguum, male (Argentina, Buenos Aires, Punta Lara); 2, A. silviae, male (Holotype: Paraguay, Caaguazú); 3, Homeoura chelifera, male (Argentina, Salta, Campo Alegre); 4, H. lindneri, male (Argentina, Corrientes, Mercedes); 5-6, H. lindneri, female (Argentina, Corrientes, Iberá); 7, H. nepos, male (Bolivia, Beni, Reyes); 8, H. obrieni, male (Paratype: Colombia, Magdalena, Aracataca); 9, H. obrieni, female (Paratype: Colombia, Magdalena, Fundación); 10, H. nepos, male (Holotype: Amazon); 11-12, H. sobrina, male and female respectively (Syntypes: Peru, Pasco, Oxapampa). Fig. 10 by RWG; Figs. 11-12 redrawn from Schmidt (1942).

Figs. 13-20. Wings. 13, 15, Argentagrion ambiguum, male (Argentina, Buenos Aires, Punta Lara); 14, 20, Homeoura obrieni, male (Paratype: Colombia, Magdalena, Aracataca); 16, H. chelifera, male (Argentina, Salta, Campo Alegre); 17, H. lindneri, male (Argentina, Corrientes, Mercedes); 18, A. silviae, male (Holotype: Paraguay, Caaguazú); 19, H. nepos, male (Bolivia, Beni, Reyes).

Figs. 21-38. Thorax, dorsal view. 21, Argentagrion ambiguum, male pronotum (Argentina, Buenos Aires, Punta Lara); 22, A. silviae, male pronotum (Holotype: Paraguay, Caaguazú); 23-24, A. ambiguum, female pronotum and mesostigmal plates respectively (Argentina, Buenos Aires, Punta Lara); 25-28, Homeoura nepos, 25-26, female pronotum and mesostigmal plates respectively (Bolivia, Santa Cruz, Urubicha), 27. male pronotum (Bolivia, Beni, Reyes); 28, posterior lobe of male pronotum (Holotype: Amazon); 29-32, H. chelifera (Argentina, Salta, Campo Alegre), 29, male pronotum, 30-32, 30, female. posterior lobe of pronotum, 31, pronotum, 32, mesotigmal plates; 33-36, H. lindneri, 33, male pronotum (Argentina, Corrientes, Mercedes), 34-36, female (Argentina, Corrientes, Iberá), 34, posterior lobe of pronotum, 35, pronotum, 36, mesotigmal plates; 37-40, H. obrieni, 37, male pronotum (Paratype: Colombia, Magdalena, Aracataca), 38, posterior lobe of male pronotum (Paratype: Colombia, Magdalena, Aracataca), 39-40, female pronotum and mesostigmal plates respectively (Paratype: Colombia, Magdalena, Fundación). Fig. 28 by RWG.

Figs. 41-57. Male genital ligula. 41-42, Argentagrion ambiguum, lateral and ectal views respectively (Argentina, Buenos Aires, Punta Lara); 43-44, A. silviae, lateral and ectal views respectively (Holotype: Paraguay, Caaguazú); 45, Enallagma signatum, lateral view (USA, Virginia, Shenandoah); 46-49, Homeoura lindneri (Argentina, Corrientes, Mercedes), 46-47, lateral view, 48, ectal view, 49, inner view of first and second segments, frontal view of third segment; 50-52, H. nepos, 50, lateral view (Bolivia, Santa Cruz, Urubicha), 51-52, lateral and ectal views respectively (Holotype: Amazon); 53- 55, H. obrieni (Paratypes: Colombia, Magdalena, Aracataca), 53-54, lateral view, 55, ectal view; 56- 57, H. chelifera, lateral and ectal views respectively (Argentina, Salta, Campo Alegre). al: apical lobe; bl: bladed projections; f: flexure; if: inner fold; ip: inner process; s1-s3: genital ligula segments 1-3. Figs. 51-52 by RWG.
Diagnosis. Argentagrion can be diagnosed from all other coenagrionid genera based on its unique combination of characters: pale postocular spots transversely elongated in males and andromorphic females, about as wide as twice their length or wider (Figs. 1-2); posterior prothoracic lobe either bilobate or with a small medial triangular apophysis (Figs. 21-23); posterodorsal margin of male S10 entire with a small medio-dorsal triangular apophysis (Figs. 65- 70); distal half of male cercus bent ventrally into a triangular process (Figs. 65, 67-68, 70; as in Andinagrion, Leucobasis Rácenis, 1959 and Zoniagrion Kennedy, 1917); male genital ligula lacking inner fold and sclerotized inner bladed projections proximal to flexure, and with an inner bladed process distal to flexure (Figs. 41, 43, ip).
Species included. A. ambiguum (Ris, 1904) and A. silviae Bulla, 1971.
Argentagrion ambiguum (Ris, 1904) (Figs. 1, 13, 15, 21, 23-24, 41-42, 58, 65- 67, 86, 88)

Figs. 58-64. Color pattern. 58, Argentagrion ambiguum, 59, A. silviae, 58-59, male abdomen, dorsal view; 60, A. silviae thorax, lateral view (Holotype: Paraguay, Caaguazú); 61-62, Homeoura obrieni, 61, male thorax and S1-2, lateral view (Paratype: Colombia, Magdalena, Aracataca), 62, female S8-10, lateral view (Paratype: Colombia, Magdalena, Fundación); 63-64, Homeoura nepos male, lateral view (Holotype: Amazon), 63, thorax and S1-2, 64, part of S7-10. vs: vulvar spine. Figs. 63-64 by RWG; Figs. 58-60 from Bulla (1971b).

Figs. 65-70. male S10. 65-67, Argentagrion ambiguum (Argentina, Buenos Aires, Punta Lara), 65, medio-dorsal view (left cercus and hairs omitted), 66, dorsal view, 67, lateral view (hairs omitted); 68- 70, A. silviae (Holotype: Paraguay, Caaguazú), 68, posterior view, 69, dorsal view, 70, lateral view. ap: apophysis; ce: cercus; pa: paraprocts; sc: scalariformlike cuticle; to: tooth.
Acanthagrion ambiguum Ris, 1904: 13-14, fig. 8 (description of male and female, illustration of male S10); Calvert, 1909: 176 (comments on color and venation); Ris, 1913: 14 [ reprint 68] (mention from Argentina); Ris, 1918: 120 (mention from Catamarca, Argentina); Ris, 1928: 42 (comparison with A. lindneri); Fraser, 1947: 430 (mention from Argentina); Calvert, 1948: 24-25 (variability of female color and venation); St. Quentin, 1960: 58 (mention from Paraguay, Argentina and Brazil).
Argentagrion ambiguum; Fraser, 1948: 48-50, figs. 1-5 (designation as type species of Argentagrion, illustration of wings, pterothorax, female S8-10, male genital ligula and S10); Rácenis, 1958: 190-193 (discussion of generic placement); Donnelly & Alayo, 1966: 107-108, fig. 3 (discussion of history and comparison to Enacantha, illustration of genital ligula); Gloger, 1967: 45-46, 50-51, 57 (distribution in Argentina, discussion of history and generic placement); Bulla, 1970: 95-97, pl. 2 figs. 1-5, pl. 3 fig. 2 (description of last larval instar); Bulla, 1971a: 135-142, figs. 139-155 (redescription of adult and larva); Bulla, 1971b: 50-54, figs. 7-10 (discussion of relationship with Acanthagrion cheliferum and A. lindneri); Bulla, 1973: 95 (discussion of generic placement); Davies & Tobin, 1984: 65 (listed); De Marmels, 1989: 248 (comparison with Cyanallagma); Rodrigues Capítulo et al., 1991: 61 (listed); Steinmann, 1997: 245- 246 (synonymic list); May, 2002: 407 (character matrix).
Homeoura ambigua; Garrison, 1991: 12 (synonymic list); Rodrigues Capítulo, 1992: 54 (mention and distribution in Argentina); Bridges, 1994: VII.10 (list); Muzón & von Ellenrieder, 1998: 23 (mention and distribution in Argentina); Tsuda, 2000: 34 (synonymic list); Costa et al., 2000: 12 (mention from São Paulo); von Ellenrieder, 2000: 20-21, 23-25 (population dynamics); O'Grady & May, 2003: 2833 (character matrix); Lencioni, 2006: 36, 127, figs. B10B, 70A-I (distribution, illustration of wings, male genital ligula, abdomen and S10, female pronotum and S8-10); von Ellenrieder & Garrison, 2007: 52-53, figs. 112, 114 (characterization, illustrations of male S10 and female mesostigmal plates).
Diagnosis. Black male labrum; dorsum of head between antennae and black pale postocular spots (Fig. 1); pale in heteromorphic female labrum and entire posterior area of head. Posterior lobe of prothorax widely bilobate (Figs. 21, 23). Lateral margins of female mesostigmal plates (Fig. 24) bluntly pointed. Pt (Figs. 13, 15) rhomboidal. Dorsum of male S3-8 mostly black (Fig. 58). Male cercus about twice as long as wide in dorsal view (Fig. 66); dorsal branch of male paraproct rounded (Figs. 67). Larval prementum with three mental and five palpal setae; base of caudal lamella dark with non-discernible tracheae, primary and secondary tracheae on distal portion equally pigmented (Fig. 86).
Biology. Adults perch on twigs and leaves of emergent vegetation close to water surface and on bushes and grass blades near shores of ponds, pools, dams and slow streams rich in aquatic vegetation within grasslands and marginal forests, and fly in a slow steady beat when sunny. Bulla (1970) described the larva, which lives among filamentous algae and roots of aquatic plants; von Ellenrieder (2000) studied seasonal variations in populations of Buenos Aires.
Distribution (Fig. 88). Bolivia (Santa Cruz department), Brazil (São Paulo, Santa Catarina and Rio Grande do Sul states), Paraguay (Amambay, Concepción, San Pedro, Central and Guaira departments), Argentina (Salta, Tucumán, Catamarca, Chaco, Formosa, Corrientes, Misiones, Santa Fe, Entre Ríos and Buenos Aires provinces) and Uruguay (Florida department), from 2 to 1040 m a.s.l.
Type material. 3 m 3 F syntypes in FNS [ not examined].
Material examined (39![]() 36
36![]() ). BOLIVIA. Santa Cruz: Robore, Chiquitos, xi 1960, 1
). BOLIVIA. Santa Cruz: Robore, Chiquitos, xi 1960, 1 ![]() (UMMZ). BRAZIL. Santa Catarina: Nova Teutonia, 10 v 1939, Fritz Plaumann col., 1
(UMMZ). BRAZIL. Santa Catarina: Nova Teutonia, 10 v 1939, Fritz Plaumann col., 1 ![]() (RWG). PARAGUAY. Concepción: Concepción, 24 viii 1956, C. J. D. Brown col., 5
(RWG). PARAGUAY. Concepción: Concepción, 24 viii 1956, C. J. D. Brown col., 5 ![]() 8
8 ![]() (UMMZ), same but 2
(UMMZ), same but 2 ![]() 1
1 ![]() (RWG), Asunción, 1 viii 1956, C. J. D. Brown col., 1
(RWG), Asunción, 1 viii 1956, C. J. D. Brown col., 1 ![]() (RWG). Guaira: Villarica, 7 iv/ 21 v 1911, 9
(RWG). Guaira: Villarica, 7 iv/ 21 v 1911, 9 ![]() 9
9 ![]() (UMMZ), Mbuvevo, 9 iv/ 21 v 1911, 5
(UMMZ), Mbuvevo, 9 iv/ 21 v 1911, 5![]() 2
2![]() (UMMZ). ARGENTINA. Misiones: Puerto Rico, 04-08 iv 1971, C. M. & O. S. Flint, Jr. col., 1
(UMMZ). ARGENTINA. Misiones: Puerto Rico, 04-08 iv 1971, C. M. & O. S. Flint, Jr. col., 1 ![]() (RWG). Corrientes: Pariopá marsh and stream, on prov. road 94, aprox. 12 km N of Santo Tomé, 21 ix 2005, N. v. Ellenrieder col., 1
(RWG). Corrientes: Pariopá marsh and stream, on prov. road 94, aprox. 12 km N of Santo Tomé, 21 ix 2005, N. v. Ellenrieder col., 1 ![]() 1
1 ![]() (RWG), Mburucuyá National Park, central station, 12-14 xii 1999, C. Molineri col., 1
(RWG), Mburucuyá National Park, central station, 12-14 xii 1999, C. Molineri col., 1 ![]() (FML). Formosa: ditch 10 km N of Formosa on route 11, 5 xi 2007, R.W. Garrison & N. v. Ellenrieder col., 5
(FML). Formosa: ditch 10 km N of Formosa on route 11, 5 xi 2007, R.W. Garrison & N. v. Ellenrieder col., 5 ![]() 4
4 ![]() (RWG), same but ditch 12 km E from route 11 on route 2 to Mojón de Fierro, 1
(RWG), same but ditch 12 km E from route 11 on route 2 to Mojón de Fierro, 1 ![]() 1
1 ![]() (RWG), same but roadside pond and grassy ditches by route 81, 40 km W of Formosa, 6 xi 2007, 1
(RWG), same but roadside pond and grassy ditches by route 81, 40 km W of Formosa, 6 xi 2007, 1 ![]() 1
1![]() (RWG), same but ponds 12 km S of Gran Guardia on road 16, 1
(RWG), same but ponds 12 km S of Gran Guardia on road 16, 1 ![]() (RWG), slough 2 km S of Bañado La Estrella, 43 km N of Las Lomitas on road 28, 2
(RWG), slough 2 km S of Bañado La Estrella, 43 km N of Las Lomitas on road 28, 2 ![]() (RWG). Salta: Campamento Jakulica, 40 km W to Aguas Blancas, 20 xi 1971, Porter col., 1
(RWG). Salta: Campamento Jakulica, 40 km W to Aguas Blancas, 20 xi 1971, Porter col., 1 ![]() (FML), km 327 on National road 81, between Hickmann and Dragones, sloughs with pleuston, 30 xi 2007, N. v. Ellenrieder col., 1
(FML), km 327 on National road 81, between Hickmann and Dragones, sloughs with pleuston, 30 xi 2007, N. v. Ellenrieder col., 1 ![]() 1
1 ![]() (RWG). Santa Fe: Esperanza, 10 x 1896, 1
(RWG). Santa Fe: Esperanza, 10 x 1896, 1 ![]() (UMMZ). Entre Ríos: P.N. El Palmar, 20 xi 1987, J. Muzón col., 1
(UMMZ). Entre Ríos: P.N. El Palmar, 20 xi 1987, J. Muzón col., 1 ![]() (MLP). Buenos Aires: Arroyo Zapata, Rt. 11, 25 km E La Plata, 24 xi 1969, C. M. & O. S. Flint, Jr. col., 1
(MLP). Buenos Aires: Arroyo Zapata, Rt. 11, 25 km E La Plata, 24 xi 1969, C. M. & O. S. Flint, Jr. col., 1 ![]() (RWG), Punta Lara, road to Villa Elisa, 06 iv 1990, J. Muzón col., 2
(RWG), Punta Lara, road to Villa Elisa, 06 iv 1990, J. Muzón col., 2 ![]() 1
1 ![]() (RWG), same but 24 viii 1997, N. v. Ellenrieder col., 1
(RWG), same but 24 viii 1997, N. v. Ellenrieder col., 1 ![]() (RWG), Punta Blonce, 07 xi 1989, J. Muzón & Aguirre col., 1
(RWG), Punta Blonce, 07 xi 1989, J. Muzón & Aguirre col., 1 ![]() (RWG), Punta Indio, Arroyo Villoldo (North), 24 xi 1998, J. Muzón & N. v. Ellenrieder col., 1
(RWG), Punta Indio, Arroyo Villoldo (North), 24 xi 1998, J. Muzón & N. v. Ellenrieder col., 1 ![]() (RWG).
(RWG).
Argentagrion silviae Bulla, 1971 (Figs. 2, 18, 22, 43-44, 59-60, 68-70, 88)
Argentagrion silviae Bulla, 1971b: 51-54, figs. 1-6 (description of male, illustration of head, thorax, abdomen, genital ligula and S10); Davies & Tobin, 1984: 66 (listed); Steinmann, 1997: 246 (synonymic list).
Homeoura silviae; Garrison, 1991: 12 (synonymic list); Bridges, 1994: VII.217 (list); Tsuda, 2000: 34 (synonymic list); Costa et al., 2000: 12 (mention from São Paulo); Lencioni, 2006: 36, 131 (distribution and illustration of head, thorax, male genital ligula, abdomen and S10); Muzón et al., 2007: 303 (type catalog MLP).
Diagnosis. Labrum pale; dorsum of head between antennae and pale postocular spots with two transversely elongated spots between medial and posterior ocelli (Fig. 2). Posterior lobe of prothorax with a short medial triangular apophysis (Fig. 22). Pt (Fig. 18) rhomboidal. Dorsum of male S3-6 with black restricted to narrow mediolongitudinal stripe, limited to basal half on S7, and dorsum of S8 entirely pale (Fig. 59). Male cercus about as long as wide in dorsal view (Fig. 69); dorsal branch of male paraproct ending in a sclerotized tooth (Fig. 70). Female unknown.
Biology. Larva and habits unknown.
Distribution (Fig. 88). Paraguay (Caaguazú department) at 300 m a.s.l.
Type material. Holotype m in MLP, with following labels: [ Museo de La Plata/ Holotipo/ Argentagrion/ silviae Bulla] [ Caaguazu/ Paraguay/ 10/3/70/ Bulla col.] [ 3484] [ examined].
Remarks. The record from São Paulo state, Brazil, has not been verified because the specimen(s) upon which Costa et al. (2000) and Lencioni (2006) have based their records is/ are lost (Costa pers. comm.). The only known bona fide specimen in existence is therefore the holotype, deposited at the MLP. However, the locality belongs to the same biogeographic province (Amazon province of the Neotropical region) and the species could possibly be present there.
Homeoura Kennedy, 1920
Homeoura Kennedy, 1920: 88 (diagnosis); Davies & Tobin, 1984: 65 (listed as junior synonym of Argentagrion); Bridges, 1994: III.22 (synonymic list); Steinmann, 1997: 246 (listed as senior synonym of Argentagrion); Tsuda, 2000: 34 (synonymic list); Lencioni, 2006: 26, 36, fig. B10 (generic key and diagnosis). Type species: Ischnura neops [ sic for Agrion? nepos] by original designation.
Characterization. Small (HW 10.2- 17.5mm, abdomen 16.5-26mm) and largely black (brown in heterochromatic females) damselflies with pale blue to pale yellow areas. Head with rounded frons, longest at level of eyes; pale postocular spots present, approximately oval to rounded or triangular, narrower than twice their length, with anterior margin usually convex (Figs. 3-12), concave in some H. nepos; pale occipital bar usually present (Figs. 4-10, 12), absent in males of H. chelifera and H. sobrina (Figs. 3, 11), and usually separated from postocular spots (except in some males of H. lindneri and heteromorphic females where the entire posterior area of head dorsum is pale, i.e. Figs. 6, 9, 12). Posterior lobe of prothorax with medial portion differentiated into a trapezoidal or rectangular lobe in male (Figs. 27-29, 33, 37-38), usually into a triangular or rectangular lobe in female (Figs. 25, 30, 34-35, 39; in some females of H. chelifera and H. nepos barely bilobate, Fig. 31); pterothorax (Fig. 61) with black medio-dorsal and humeral stripes and usually a complete antehumeral pale stripe (interrupted distally in some H. nepos), usually lacking a complete black stripe over metapleural suture (present only in H. sobrina); in some heteromorphic females thorax entirely pale; tibial spurs as long as or shorter than intervening spaces; supplementary teeth of pretarsal claws well developed. CuP reaching CuPAA (Fig. 14), petiolation of wing ending at or proximal to CuP (variable intraspecifically), pt quadrangular (H. nepos and H. obrieni, Figs. 14, 19-20) or rhomboidal (H. chelifera and H. lindneri, Figs. 16-17). Male genital ligula with one latero-distal lobe on each side, lacking inner fold, with a small digitiform inner process in the center of a transverse fold distal to flexure, and with a pair of unique sclerotized projections shaped as triangular blades proximal to flexure (Figs. 46, 47, 50, 53-56, bl), setae on sides of second segment usually present. Posterior margin of male S10 with a 'u' shaped medial cleft with two apical tubercles (Figs. 71-72, 74-85, ct). Male cerci approximately horizontal with inner margin projected ventro-basally into a laminar process (Figs. 71-76, 78, 80, 82, 85); paraprocts short with dorsal branch ending in a sclerotized tip (Figs. 74-76, 78, 80, to). Female vulvar spine of S8 well developed; outer valves of ovipositor with one row of spines, and extending slightly beyond posterior margin of S10 (Fig. 62).
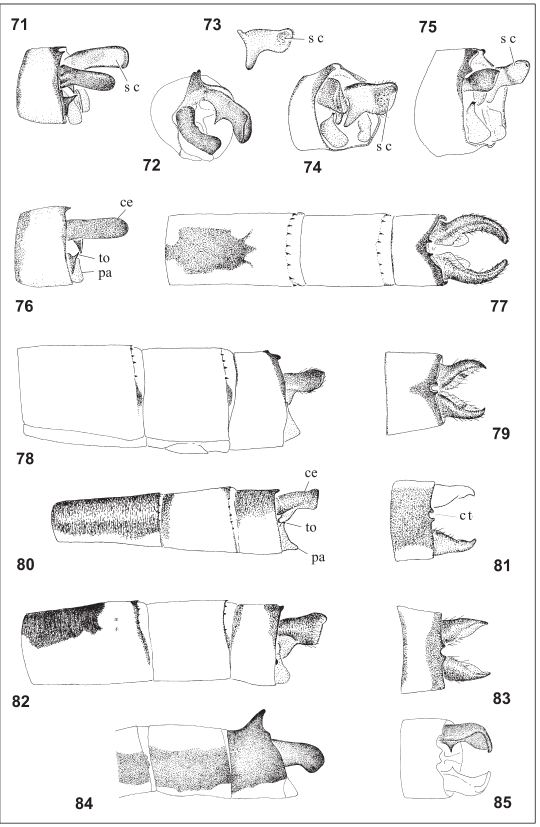
Figs. 71-85. Male abdomen. 71, Homeoura chelifera (Argentina, Salta, Campo Alegre), 72, H. sobrina (Syntype: Peru, Pasco, Oxapampa), modified from Schmidt (1942), 73, H. nepos, right cercus, inner view (Bolivia, Beni, Reyes), 74, H. obrieni (Paratype: Colombia, Magdalena, Aracataca), 75, H. lindneri (Argentina, Corrientes, Mercedes), 71, 72, 74-75, S10, medio-dorsal view; 76-77, H. chelifera (Argentina, Salta, Campo Alegre), 76, S10, lateral view, 77, S8-10, dorsal view; 78-79, H. lindneri (Argentina, Corrientes, Mercedes), 78, S8-10, lateral view, 79, S10, dorsal view; 80-81. H. nepos (Bolivia, Beni, Reyes), 80, S8-10, lateral view, 81, S10, dorsal view; 82-83, H. obrieni (Paratype: Colombia, Magdalena, Aracataca), 82, 84, 85, H. sobrina (Syntype: Peru, Pasco, Oxapampa), redrawn from Schmidt (1942), 84, part of S8-10, lateral view, 85, S10, dorsal view. ce: cercus; ct: S10 postero-dorsal cleft with tubercles; pa: paraprocts; sc: scalariformlike cuticle; to: tooth.

Figs. 86-87. Larval lateral caudal lamella. 86, Argentagrion ambiguum; 87, Homeoura chelifera. Fig. 86 from Bulla (1970); Fig. 87 from Bulla (1971a).

Fig. 88. Distribution of Argentagrion in South America.
Diagnosis. Homeoura differs from Argentagrion by the medial lobe of posterior lobe of prothorax projected into a rectangular or trapezoidal process in males (projected medial lobe shared with Andinagrion Bulla, Cyanallagma, Hesperagrion Calvert, Mesamphiagrion Kennedy, Oxyallagma Kennedy and Protallagma Kennedy), the presence of a 'u' shaped cleft with two apical tubercles on postero-dorsal margin of male S10 (shared with previous genera and with Enallagma, Enacantha Donnelly & Alayo and some Ischnura species) and male cercus approximately horizontal with a ventro-basal branch, which is a projection of its ventromesal margin (as in Enallagma and Mesamphiagrion). The paired sclerotized projections shaped as triangular blades proximal to flexure on second segment of male genital ligula of Homeoura distinguish it from all other genera of New World Coenagrionidae.
Remarks. The «bladed projection» (De Marmels, 1989) observed in Enallagma signatum (Hagen, 1861) is single (Fig. 45), and therefore not equal to the projection of all Homeoura species, which is composed of two parallel blades (Figs. 46-47, 49-50, 53-54, 56, bl).
Species included. H. chelifera (Selys, 1876), H. lindneri (Ris, 1928), H. nepos (Selys, 1876), H. obrieni n. sp. and H. sobrina (Schmidt, 1942) n. comb.
Homeoura chelifera (Selys, 1876) (Figs. 3, 16, 29-32, 56-57, 71, 76-77, 87, 89)

Fig. 89. Distribution of Homeoura in South America.
Acanthagrion? cheliferum Selys, 1876: 319- 321 [ reprint 75-77] (description of male).
Acanthagrion cheliferum? rusticum Calvert, 1909: 175-176 (description as possible subspecies).
Acanthagrion cheliferum; Ris, 1904: 12-13, fig. 7 (description of female, illustration of male S10); Needham, 1904: 717 (incomplete larval description); Calvert, 1909: 175-176 (comments on color and venation); Ris, 1913: 14 [ reprint 68] (mention from Argentina); Ris, 1928: 41, 44 (comparison with A. lindneri); Fraser, 1947: 430 (mention from Argentina); Bulla, 1971a: 159-166, figs. 175-182, 246 (redescription of adult and larva); Bulla, 1971b: 49-51 (discussion of relationship with Argentagrion); Bulla, 1973: 95 (comparison with Cyanallagma bonariense and C. interruptum).
Enallagma cheliferum; Santos, 1956: 571- 576, figs. 1-12 (new combination proposed, discussion of similarity with Enallagma species, redescription of male and female, illustrations of head, thorax, abdomen, wings); De Marmels, 1990: 335 (mention for Venezuela).
Argentagrion cheliferum; Rácenis, 1958: 190- 192 (mention from Venezuela, discussion of generic placement); Donnelly & Alayo, 1966: 108-109, fig. 3 (discussion of history and comparison with Enacantha, illustration of genital ligula); Gloger, 1967: 45-46, 50, 55, 57 (distribution in Argentina, discussion of history and generic placement); Davies & Tobin, 1984: 65 (listed); Rodrigues Capítulo et al., 1991: 61, 66 (chromosomal data); Steinmann, 1997: 246 (synonymic list); May, 2002: 407 (character matrix).
Cianallagma chelifera; Cruz, 1986: 745-747, fig. 8 (comparison with C. demarmelsi, illustration female posterior prothoracic lobe and mesostigmal plates).
Homeoura chelifera; Garrison, 1991: 12 (synonymic list); Rodrigues Capítulo, 1992: 54 (mention and distribution in Argentina); Bridges, 1994: VII.49 (list); De Marmels, 1997: 136 (comparison with Cyanallagma); Muzón & von Ellenrieder, 1998: 23 (mention and distribution in Argentina); Tsuda, 2000: 34 (synonymic list); Costa et al., 2000: 12 (mention from São Paulo); von Ellenrieder, 2000: 20-21, 23-25 (population dynamics); Lencioni, 2006: 36, 128, figs. B10A, 71A-I (distribution, illustration of wings, head, thorax, male genital ligula, abdomen and S10, female pronotum, mesostigmal plates and S8-10); von Ellenrieder & Garrison, 2007: 52-53, figs. 110-111, 113 (characterization, picture, illustrations of male S10 and female mesostigmal plates).
Diagnosis. Pale postocular spots on dorsum of male head (Fig. 3) large (larger than 1/3 of postocular lobe's width) and approximately oval (shared with H. lindneri and H. nepos), and pale postocular bar absent (shared with H. sobrina); in heteromorphic female entire posterior area of head pale (shared with all Homeoura species). Medial projection of posterior lobe of prothorax rectangular (Figs. 29-30) (shared with H. nepos) or narrowly bilobate (Fig. 31) in some females (unique). Lateral margins of female mesostigmal plates (Fig. 32) truncated (unique). Pt (Fig. 16) rhomboidal (shared with H. lindneri). Second segment of genital ligula with a few thin hairs on each side or lacking hairs (Figs. 56-57), and bladed projections proximal to flexure in the shape of equilateral triangles, with acutely pointed apex and about as high as wide (Fig. 56; both shared with H. lindneri and H. obrieni). Dorsum of male S8 partially or entirely black, S9 entirely pale and S10 mostly pale, with black posterior margin (Figs. 76, 77; shared with H. obrieni). Dorsal and ventral margins of male cercus (Fig. 76) approximately parallel and of equal length in lateral view, so that apical width is approximately the same as basal width and apex is roundly truncated (shared with H. sobrina). Larval prementum with three mental and five palpal setae; caudal lamella with three diffuse dark spots along midline and secondary tracheae much darker than primary (Fig. 87).
Biology. Adults perch on twigs and leaves of emergent vegetation close to water surface and on bushes and grass blades near the shores of ponds, pools, dams and slow streams rich in aquatic vegetation, and fly in a slow steady beat when sunny (von Ellenrieder & Garrison, 2007). Needham (1904) provided a short description of the larva based on an incomplete putative specimen of H. chelifera from São Paulo, which was later complemented by Bulla (1971a); von Ellenrieder (2000) studied seasonal variations in populations from Buenos Aires.
Distribution (Fig. 89). Colombia (Antioquia department), Venezuela (Aragua, Guárico, Barinas and Apure states), Guyana (Upper Takutu-Upper Essequibo region), Brazil (Bahia, Minas Gerais, Espirito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina and Rio Grande do Sul states), Paraguay (Guaira and Paraguarí departments), Uruguay (de Rocha department) and Argentina (Jujuy, Salta, Tucumán, Corrientes, Santa Fe, Entre Ríos and Buenos Aires provinces), from 4 to 1530 m a.s.l.
Type material. Types of Acanthagrion? cheliferum: 1 ![]() lectotype by present designation; 7
lectotype by present designation; 7 ![]() paralectotypes in IRSNB [ examined]. In order to clarify application of the name, I designate the syntype male from Brazil, Minas Gerais, Entre Ríos, ix, Walthère de Selys col. as LECTOTYPE; labels: Entrerios; cheliferum; W. de Selys. Paralectotypes: Minas Gerais, 3
paralectotypes in IRSNB [ examined]. In order to clarify application of the name, I designate the syntype male from Brazil, Minas Gerais, Entre Ríos, ix, Walthère de Selys col. as LECTOTYPE; labels: Entrerios; cheliferum; W. de Selys. Paralectotypes: Minas Gerais, 3 ![]() (IRSNB), Santa Teresa, 1
(IRSNB), Santa Teresa, 1 ![]() (IRSNB), Rio de Janeiro, 2
(IRSNB), Rio de Janeiro, 2 ![]() (IRSNB), Rio Grande, 1
(IRSNB), Rio Grande, 1![]() (IRSNB). Types of Acanthagrion? cheliferum rusticum: 2
(IRSNB). Types of Acanthagrion? cheliferum rusticum: 2![]() , 1
, 1 ![]() syntypes in MCZ [ not examined].
syntypes in MCZ [ not examined].
Material examined (44 ![]() 13
13![]() ).
).
VENEZUELA. Barinas: NE to Mantecal, marshy pond at entrance to Hato El Frío, 27 xii 2000, D.R. Paulson & N. Smith col., 1![]() 1
1 ![]() (DRP). COLOMBIA. Antioquia: Represa La Fe, W of La Fe, 26 ii 1984, C.M. & O.S. Flint, Jr. col., 1
(DRP). COLOMBIA. Antioquia: Represa La Fe, W of La Fe, 26 ii 1984, C.M. & O.S. Flint, Jr. col., 1 ![]() 1
1 ![]() (RWG). BRAZIL. Minas Gerais: Entre Ríos, ix, Walthère de Selys col., 1
(RWG). BRAZIL. Minas Gerais: Entre Ríos, ix, Walthère de Selys col., 1 ![]() lectotype (IRSNB), Minas Gerais, 3
lectotype (IRSNB), Minas Gerais, 3![]() paralectotypes (IRSNB), Belo Horizonte, Lagoa Santa, 15 x 1978, D.A.L. Davies col., 1
paralectotypes (IRSNB), Belo Horizonte, Lagoa Santa, 15 x 1978, D.A.L. Davies col., 1 ![]() (RWG). Espirito Santo: Santa Teresa, 1
(RWG). Espirito Santo: Santa Teresa, 1 ![]() paralectotype (IRSNB). Rio de Janeiro: Rio de Janeiro, 2
paralectotype (IRSNB). Rio de Janeiro: Rio de Janeiro, 2 ![]() paralectotypes (IRSNB), same but 27 ix 1922, J.H. Williamson col., 3
paralectotypes (IRSNB), same but 27 ix 1922, J.H. Williamson col., 3![]() (RWG). Santa Catarina: Nova Teutonia, 3 iv /25 iii 1945, F. Plaumann col., 6
(RWG). Santa Catarina: Nova Teutonia, 3 iv /25 iii 1945, F. Plaumann col., 6 ![]() 1
1 ![]() (RWG). São Paulo: Fazenda Santana do Rio Abaixo, Jacareí, 08 xii 1998, F. A. A. Lencioni col., 2
(RWG). São Paulo: Fazenda Santana do Rio Abaixo, Jacareí, 08 xii 1998, F. A. A. Lencioni col., 2 ![]() 1
1 ![]() (RWG), same but 15 ix 1996, 1
(RWG), same but 15 ix 1996, 1 ![]() (RWG), Salesopolis, 14 iii 1999, F. A. A. Lencioni col., 1
(RWG), Salesopolis, 14 iii 1999, F. A. A. Lencioni col., 1 ![]() (RWG), Campos do Jordão, 02 xi 1999, F. A. A. Lencioni col., 1
(RWG), Campos do Jordão, 02 xi 1999, F. A. A. Lencioni col., 1![]() (RWG), Rio Claro, 10 ix 1996, F. A. A. Lencioni col., 1
(RWG), Rio Claro, 10 ix 1996, F. A. A. Lencioni col., 1![]() (RWG), São Paulo (RWG), 3
(RWG), São Paulo (RWG), 3 ![]() Cipó, 8 iv 1967/ 2 iii 1968, V. N. Alin, 2
Cipó, 8 iv 1967/ 2 iii 1968, V. N. Alin, 2![]() 1
1 ![]() (DRP). Rio Grande do Sul: Rio Grande do Sul, 1
(DRP). Rio Grande do Sul: Rio Grande do Sul, 1![]() paralectotype (IRSNB). PARAGUAY. Guaira: Mbuevo, 9 iv 1911, 1
paralectotype (IRSNB). PARAGUAY. Guaira: Mbuevo, 9 iv 1911, 1 ![]() (RWG). ARGENTINA. Jujuy: La Angostura, pond at house, 1
(RWG). ARGENTINA. Jujuy: La Angostura, pond at house, 1 ![]() (RWG). Salta: pond between La Angostura and Campo Alegre, 21 iv 2001, N. v. Ellenrieder col., 6
(RWG). Salta: pond between La Angostura and Campo Alegre, 21 iv 2001, N. v. Ellenrieder col., 6 ![]() 1
1 ![]() (RWG), Campo Alegre Dam, 12 i 1997, T.W. Donnelly col., 1
(RWG), Campo Alegre Dam, 12 i 1997, T.W. Donnelly col., 1 ![]() (RWG), same but J. Muzón & N. v. Ellenrieder col., 1
(RWG), same but J. Muzón & N. v. Ellenrieder col., 1 ![]() 1
1 ![]() (RWG). Tucumán: pond at Museo, 23 xi 1968, L.A. Bulla col., 2
(RWG). Tucumán: pond at Museo, 23 xi 1968, L.A. Bulla col., 2 ![]() (MLP). Corrientes: Pariopá marsh and stream, on prov. road 94, aprox. 12 km N of Santo Tomé, 21 ix 2005, N. v. Ellenrieder col., 2
(MLP). Corrientes: Pariopá marsh and stream, on prov. road 94, aprox. 12 km N of Santo Tomé, 21 ix 2005, N. v. Ellenrieder col., 2 ![]() (RWG), Mburucuyá National Park, central station, 12-14 xii 1999, C. Molineri col., 1
(RWG), Mburucuyá National Park, central station, 12-14 xii 1999, C. Molineri col., 1 ![]() (FML). Santa Fe: Santo Tomé, Río Salado, 30-31 iii 1971, C.M. & O.S. Flint, Jr., 1
(FML). Santa Fe: Santo Tomé, Río Salado, 30-31 iii 1971, C.M. & O.S. Flint, Jr., 1 ![]() (RWG). Buenos Aires: Punta Lara, grassland, 07 xi 1997, N. v. Ellenrieder col., 1
(RWG). Buenos Aires: Punta Lara, grassland, 07 xi 1997, N. v. Ellenrieder col., 1 ![]() (RWG), Punta Indio, stream on km 223 of road 11, 1,5 km N Arroyo La Matilde, 25 xi 1998, J. Muzón & N. v. Ellenrieder col., 1
(RWG), Punta Indio, stream on km 223 of road 11, 1,5 km N Arroyo La Matilde, 25 xi 1998, J. Muzón & N. v. Ellenrieder col., 1 ![]() (MLP), Lima, Club de Pesca near Central Nuclear Atucha I, 31 x 1995, N. v. Ellenrieder col., 2
(MLP), Lima, Club de Pesca near Central Nuclear Atucha I, 31 x 1995, N. v. Ellenrieder col., 2 ![]() (MLP).
(MLP).
Remarks. Needham (1904) mentioned one mental seta and seven palpal setae for the single «mutilated» larva at his disposal, which he identified by association with adults from the same locality. This does not agree with bona fide reared material of H. chelifera from Argentina, which shows three and five setae respectively (Bulla, 1971a). Needham's description (1904) probably does not apply to this species.
Homeoura lindneri (Ris, 1928) (Figs. 4-6, 17, 33-36, 46-49, 75, 78-79, 89)
Acanthagrion lindneri Ris, 1928: 41-44, figs. 1-2 (description of male and female, illustration of male S10, comparison with A. ambiguum and A. cheliferum); Calvert, 1948: 23-24, figs. 18-22 (illustrations of male S10, posterior lobe of prothorax and genital ligula based on specimen from Pirassununga, São Paulo state, Brazil); Fraser, 1947: 431 (mention from Argentina); Bulla, 1971a: 167-171, figs. 183-191 (redescription of adult); Bulla, 1971b: 49-51 (discussion of relationship with Argentagrion); Bulla, 1973: 95 (discussion of generic affinities, comparison with Cyanallagma bonariense and C. interruptum).
Argentagrion lindneri; Donnelly & Alayo, 1966: 108-109 (discussion of history); Davies & Tobin, 1984: 66 (listed); Kiauta, 1988: 438 (incorrect statement of Argentagrion lindneri being a junior synonym of A. nepos); Rodrigues Capítulo et al., 1991: 61 (listed); Steinmann, 1997: 246 (synonymic list).
Cianallagma lindneri; Cruz, 1986: 745-747, fig. 8 (comparison with C. demarmelsi, illustration of female posterior prothoracic lobe and mesostigmal plates).
Homeoura lindneri; De Marmels, 1989: 248 (comparison with Cyanallagma); Garrison (1991: 12; synonymic list); Rodrigues Capítulo, 1992: 54 (mention and distribution in Argentina); Bridges (1994: VII.135; list); Muzón & von Ellenrieder, 1998: 23 (mention and distribution in Argentina); Tsuda, 2000: 34 (synonymic list); Costa et al., 2000: 12 (mention from São Paulo); Lencioni, 2006: 36, 129, figs. 72A-C (distribution, illustration of male S10 and pt).
Ischnura sobrina; O'Grady & May, 2003: 2826, 2833 (character matrix based on male from Peru, Explorama Lodge) misidentified material.
Diagnosis. Pale postocular spots on dorsum of male and andromorphic female head (Figs. 4-5) large (larger than 1/3 of postocular lobe's width) and approximately oval (shared with H. chelifera and H. nepos), and pale postocular bar present (shared with H. nepos and H. obrieni), sometimes both are fused (Fig. 5); in heteromorphic female (Fig. 6) entire posterior area of dorsum pale (shared with all Homeoura species). Medial projection of posterior lobe of prothorax trapezoidal to rectangular in male (Fig. 33) and triangular to rectangular (Figs. 34-35) in female (shared with H. obrieni). Lateral margins of female mesostigmal plates (Fig. 36) are bluntly pointed (shared with H. obrieni). Pt (Fig. 17) rhomboidal (shared with H. chelifera). Second segment of genital ligula with a few thin hairs on each side or lacking hairs (Figs. 46-49), and bladed projections proximal to flexure in the shape of equilateral triangles, with acutely pointed apex and about as high as wide (Figs. 46-47; both shared with H. chelifera and H. obrieni). Male S8-9 entirely pale and S10 mostly pale, with black posterior margin (Figs. 78-79; unique). Dorsal and ventral margins of male cercus (Fig. 78) in lateral view diverging apically, and dorsal margin longer than ventral, so that apical width is larger than basal width, and apex is obliquely truncated dorso-ventrally (shared with H. nepos and H. obrieni).
Biology. Adults collected at ponds and marshes (locality data from collections). Larva unknown.
Distribution (Fig. 89). Peru (Loreto department), Brazil (São Paulo state), Paraguay (San Pedro department) and Argentina (Corrientes, Formosa, Chaco, Santa Fe and Buenos Aires provinces), from 14 to 630 m a.s.l.
Type material. 1 ed ![]() 1
1 ![]() syntypes in FNS [ not examined].
syntypes in FNS [ not examined].
Material examined (12 ![]() 3
3 ![]() ). PERU. Loreto: Explorama Lodge, 25 mi NE Iquitos on Amazon River, 17/21 vii 1989, G. B. Edwards col., 1
). PERU. Loreto: Explorama Lodge, 25 mi NE Iquitos on Amazon River, 17/21 vii 1989, G. B. Edwards col., 1 ![]() (SWD), Explorama Lodge, 50 mi NE Iquitos on Amazon River at junction with Yanamono River, Manatí river and pasture pond at Lodge, 16/ 31 viii 1989, S.W. Dunkle col., 2
(SWD), Explorama Lodge, 50 mi NE Iquitos on Amazon River at junction with Yanamono River, Manatí river and pasture pond at Lodge, 16/ 31 viii 1989, S.W. Dunkle col., 2 ![]() 1
1 ![]() (SWD), same but 4 vii 1991, S.W. Dunkle col., 1
(SWD), same but 4 vii 1991, S.W. Dunkle col., 1 ![]() (SWD). PARAGUAY. San Pedro: Antequera, xii 1979, L. Strickman col., 1
(SWD). PARAGUAY. San Pedro: Antequera, xii 1979, L. Strickman col., 1 ![]() (DRP). ARGENTINA. Corrientes: Laguna Iberá, 3 xi 2003, J. Muzón & P. Pessacq, 3
(DRP). ARGENTINA. Corrientes: Laguna Iberá, 3 xi 2003, J. Muzón & P. Pessacq, 3 ![]() 2
2![]() (MLP), Mercedes, 03 x 2004, F. Lozano & A. Garré col., 1
(MLP), Mercedes, 03 x 2004, F. Lozano & A. Garré col., 1![]() (MLP), same but 2
(MLP), same but 2 ![]() (RWG). Santa Fe: Laguna Don Felipe, E of Santa Fe, 01 iv 1971, C.M. & O.S. Flint, Jr. col., 1
(RWG). Santa Fe: Laguna Don Felipe, E of Santa Fe, 01 iv 1971, C.M. & O.S. Flint, Jr. col., 1 ![]() (RWG).
(RWG).
Remarks. Kiauta (1988) in an abstract review declared «Argentagrion» lindneri a synonym of «A.» nepos with no further discussion («….further, lindneri is in Argentagrion, and is a junior synonym of nepos.»). My examination shows them to be distinct.
Homeoura nepos (Selys, 1876) (Figs. 7, 10, 19, 25-27, 50-52, 63-64, 73, 80-81, 89)
Agrion? nepos Selys, 1876: 1246-1247, 1249- 1250 [ reprint 136-137, 139-140] (description of male).
Coenagrion? nepos; Kirby, 1890: 148 (mention).
Ischnura? nepos; Calvert, 1909: 206-209, figs. 121-122 (description of female, illustration of male S10); Calvert, 1948: 9-10 (incomplete description of larva by supposition).
Homeoura nepos; Kennedy, 1920: 88 (designation as type species of Homeoura); De Marmels, 1989: 248 (comparison to Cyanallagma); De Marmels, 1990: 335 (mention from Venezuela); Garrison, 1991: 12 (synonymic list); Bridges (1994: VII.163; list); Tsuda, 2000: 34 (synonymic list); Costa et al., 2000: 12 (mention from São Paulo); May, 2002: 407 (character matrix); Lencioni, 2006: 36, 130, figs. 73A-H (distribution, illustration of male head, thorax, pronotum, pt, abdomen, genital ligula and S10).
Argentagrion nepos; Davies & Tobin, 1984: 66 (listed); Kiauta, 1988: 438 (incorrect statement of Argentagrion lindneri being a junior synonym of A. nepos); Machet, 1989: 11 (mention from French Guiana); Steinmann, 1997: 246 (synonymic list).
Diagnosis. Pale postocular spots on dorsum of male head (Figs. 7, 10) large (larger than 1/3 of postocular lobe's width) and approximately oval (shared with H. chelifera and H. lindneri), sometimes with concave anterior margin and pale postocular bar present (shared with H. lindneri and H. obrieni), in heteromorphic female entire posterior area of dorsum pale (shared with all Homeoura species). Medial projection of posterior lobe of prothorax approximately rectangular (shared with H. chelifera) with posterior margin slightly concave to linear (Figs. 27-28), reduced to absent in some females (Fig. 25). Lateral margins of female mesostigmal plates (Fig. 26) bluntly rounded, anterior and posterior margins approximately parallel-sided (Fig. 26; unique). Pt (Fig. 19) quadrangular (shared with H. obrieni). Second segment of genital ligula with a dense row of strong hairs on each side (Figs. 50-52; unique), and bladed projections are proximal to flexure with bluntly pointed apex and considerably wider than high (Fig. 50; unique). Dorsum of male S8 entirely black, of S9 mostly pale with black anterior margin, of S10 entirely or mostly black (Figs. 64, 80-81; unique). Dorsal and ventral margins of male cercus (Fig. 80) in lateral view diverging apically, and dorsal margin longer than ventral, so that apical width is larger than basal width and apex is obliquely truncated dorsoventrally (shared with H. lindneri and H. obrieni).
Larval prementum with two long and two short mental setae and four palpal setae (Calvert, 1948).
Biology. Adults found at lentic environments (locality data from collections). Larva described by supposition by Calvert (1948) based on an incomplete specimen from a dam in Pirassununga, Brazil.
Distribution (Fig. 89). Brazil (Amazonas, Pará, Rondônia, São Paulo, Mato Grosso and Mato Grosso do Sul states), Paraguay (Concepción and Amambay departments) and Bolivia (Beni, Santa Cruz and Tarija departments), from 10 to 635 m a.s.l.
Type material. Holotype m from IRSNB currently on loan to MNRJ [ not examined].
Material examined (23 ![]() 14
14![]() ). PERU. Madre de Dios: Explorer's Inn on Rio Tambopata, 30 km SW Puerto Maldonado, oxbow lake, 15 /16 vi 1977, D.R. Paulson col., 1
). PERU. Madre de Dios: Explorer's Inn on Rio Tambopata, 30 km SW Puerto Maldonado, oxbow lake, 15 /16 vi 1977, D.R. Paulson col., 1 ![]() 1
1 ![]() (DRP). BRAZIL. Pará: Santarem, 1-31 v 1919, S. M. Klages col., 1
(DRP). BRAZIL. Pará: Santarem, 1-31 v 1919, S. M. Klages col., 1![]() 1
1![]() (RWG), same but iv/ vii 1919, 4
(RWG), same but iv/ vii 1919, 4 ![]() 2
2![]() (FSCA). Rondônia: 60 km S of Ariquemes, Fazenda Rancho Grande, farm of Harold Schmitz and environs within about 15 km radius, 16/24 iii 1989, S. W. Dunkle, 1
(FSCA). Rondônia: 60 km S of Ariquemes, Fazenda Rancho Grande, farm of Harold Schmitz and environs within about 15 km radius, 16/24 iii 1989, S. W. Dunkle, 1 ![]() 3
3 ![]() (SWD), same but 15/ 22 iii 1991, F. W. Skillman Jr. col., 1
(SWD), same but 15/ 22 iii 1991, F. W. Skillman Jr. col., 1 ![]() (SWD). BOLIVIA. Beni: Reyes, 27-30 xi 1957, L. Peña col., 1
(SWD). BOLIVIA. Beni: Reyes, 27-30 xi 1957, L. Peña col., 1 ![]() (RWG). Santa Cruz: Provincia del Sara: J. Steinbach col., 1
(RWG). Santa Cruz: Provincia del Sara: J. Steinbach col., 1 ![]() (FSCA), Puerto Suárez, xi 1960, 5
(FSCA), Puerto Suárez, xi 1960, 5 ![]() 4
4 ![]() (RWG), Los Lajas hacienda and lagunas, 5 km S Guarayos, 27 viii 2003, B. Mauffray col., 1
(RWG), Los Lajas hacienda and lagunas, 5 km S Guarayos, 27 viii 2003, B. Mauffray col., 1 ![]() (FSCA), 1.5 km N Urubicha, Río Blanco and adjacent laguna Guarayos, 19 viii 2003, B. Mauffray col., 1
(FSCA), 1.5 km N Urubicha, Río Blanco and adjacent laguna Guarayos, 19 viii 2003, B. Mauffray col., 1 ![]() 1
1 ![]() (FSCA), 1.7 km W Ascención de Guarayos, laguna La Tejerina, 14 viii 2003, B. Mauffray col., 2
(FSCA), 1.7 km W Ascención de Guarayos, laguna La Tejerina, 14 viii 2003, B. Mauffray col., 2 ![]() (FSCA), 15 km W Yotau, Cachuelos de Yotau, 15 viii 2003, B. Mauffray col., 1
(FSCA), 15 km W Yotau, Cachuelos de Yotau, 15 viii 2003, B. Mauffray col., 1 ![]() (FSCA), hacienda Progreso and lagunas, 13 km S on Hwy 9, 17 viii 2003, B. Mauffray col., 1
(FSCA), hacienda Progreso and lagunas, 13 km S on Hwy 9, 17 viii 2003, B. Mauffray col., 1 ![]() 1
1 ![]() (FSCA), 23.5 km N on Hwy 9 of Ascención de Guarayos, borrow pit pond and adjacent forest, 26 viii 2003, B. Mauffray col., 1
(FSCA), 23.5 km N on Hwy 9 of Ascención de Guarayos, borrow pit pond and adjacent forest, 26 viii 2003, B. Mauffray col., 1 ![]() (FSCA). PARAGUAY. Amambay: Cerro Cora, 16/17 viii 1980, L. Strickman col., 1
(FSCA). PARAGUAY. Amambay: Cerro Cora, 16/17 viii 1980, L. Strickman col., 1![]() 1
1 ![]() (DRP).
(DRP).
Remarks. As stated above, the holotype is currently deposited in the MNRJ, where RWG examined it and illustrated the head (Fig. 10), thorax (Figs. 28, 63), genital ligula (Figs. 51, 52), and S1-3, 7-10 (Fig. 64). Although the appendages are missing, comparisons of RWG illustrations of pronotum and color pattern with material I have identified as this species show that I have correctly applied the name to material discussed here. Synonymy of «Argentagrion» lindneri with «A.» nepos by Kiauta (1988) is discussed under H. lindneri above. The records of H. nepos from la Guyane department, French Guiana (Machet, 1989) need to be confirmed by examination of specimens, since they could as well correspond to H. obrieni (Fig. 89).
Homeoura obrieni sp. nov. (Figs. 8-9, 14, 20, 37-40, 53-55, 61-62, 74, 82-83, 89)
Argentagrion lindneri; Rácenis, 1958: 191- 193 (placement in Argentagrion, comparison with original description and redescription of A. lindneri by Calvert, 1948, based on male from Suripá, Estado Barinas, 14.iii.1954, Szumkowsky leg.) misidentified material.
Argentagrion nepos; Donnelly & Alayo (1966: 108-109, fig. 3; at least in part, specimen from Aracataca, Colombia; illustration of genital ligula) misidentified material. Homeoura nepos; De Marmels (1990: 335; mention from Venezuela) misidentified material.
Diagnosis. Pale postocular spots on dorsum of male head (Fig. 8) about as large as 1/3 of postocular lobe width and approximately rounded, and pale postocular bar present (shared with H. lindneri and H. nepos). Medial projection of posterior lobe of prothorax is rhomboidal in male (Figs. 37- 38) and triangular (Fig. 39) in female (shared with H. lindneri). Lateral margins of females' mesostigmal plates (Fig. 40) are bluntly pointed (shared with H. lindneri). Pt (Figs. 14, 20) quadrangular (shared with H. nepos). Dorsum of male S8 partially black, always distally pale, S9 entirely pale and S10 mostly pale, with black posterior margin (Figs. 82- 83; shared with H. chelifera) or black anterior and posterior margins, occasionally joined by black narrow mid-dorsal stripe. Dorsal and ventral margins of male cercus (Fig. 82) in lateral view diverging apically, and dorsal margin longer than ventral, so that apical width is larger than basal width and apex is obliquely truncated dorso-ventrally (shared with H. lindneri and H. nepos).
Description. Male holotype. Head: labium pale yellow; labrum, base of mandibles, anterior surface of genae and anteclypeus light blue, with a basal black stripe on labrum and two short black stripes on anteclypeus; postclypeus, dorsal surface of genae, frons, antennae and top of head black, with a small pale blue spot anterior to lateral ocellus, large pale blue postocular spot and occipital bar; back of head pale blue, with a black rounded spot to each side of occipital foramen. Frons in profile rounded. Thorax: prothorax black with pale blue spots; medial lobe of posterior prothoracic lobe trapezoidal (as in Figs. 37-38). Dorsum of pterothorax black with complete pale blue antehumeral stripe about as wide as a fourth of mesepisternum, slightly widened at anterior and posterior ends; sides pale blue with a small black spot on posterior end of metapleural suture (as in Fig. 61). Venter of thorax pale yellow, with a low postero-medial rounded tubercle. Legs yellow with black spurs, extensor surface of femora, a narrow stripe along outer side of tibiae and distal tip of tarsi and pretarsi (as in Fig. 61). Spurs on femora and tibiae shorter than intervening spaces; pretarsal claw with well developed supplementary tooth (as in Fig. 61). Wings (as in Fig. 14) hyaline; pt reddish brown, quadrangular, shorter than underlying cell, with anterior (costal) margin the shortest (as in Fig. 20); CuP reaching CuPAA distal to confluence of CuPAA with A, for a distance equal to CuP in FW, and shorter than CuP in HW, petiolation ending at about a third of the distance between Ax 1 and Ax 2; Px 7 in FW, 6 in right HW, 8 in left HW; RP2 branching between Px 3 and 4 but closer to 4 in FW and right HW, midway between Px 3 and 4 in left HW. Abdomen: S1pale blue with dorsal black spot on basal two thirds; dorsum of S2 black, sides pale blue; dorsum of S3-7 black except basal pale blue ring, sides pale blue; S8 pale blue with black spot on an antero-dorsal three fifths; S9 pale blue; S10 pale blue with narrow transverse anterodorsal and postero-dorsal black stripes. Cercus (as Figs. 74, 82-83) shorter than S10, with a ventro-basal triangular process directed anteriorly on inner margin; a distal area of differentiated scalariform like cuticle (May, 2002) on mesal surface (as in Fig. 74); dorsal margin slightly longer than and diverging gradually from ventral margin, so that posterior margin is oblique (as in Fig. 82); color black except pale brown mesal surface and distal portion of outer surface. Paraproct yellow, with a small medially directed black tooth on dorsal end (as in Fig. 74). Second segment of genital ligula with a few thin hairs on each side (as in Figs. 53-55), and bladed projections proximal to flexure in the shape of equilateral triangles, with acutely pointed apex and about as high as wide (as in Figs. 53-54).
Dimensions: total length 25.7 mm; abdomen length 20.6 mm; FW 13mm; HW 12.2 mm.
Female allotype. Head: as in holotype but postclypeus pale blue with anterior margin black; pale spot anterior to lateral ocellus larger; additional small pale spot posterior to base of antenna; pale blue postocular spot confluent with pale occipital bar. Thorax: color pattern as in holotype but pale blue areas on prothorax more extensive; and a narrow yellow stripe along humeral suture; legs as in holotype but black areas brown. Medial lobe of posterior prothoracic lobe bluntly triangular (as in Fig. 39); mesostigmal plate with basal constriction, so that anterior and posterior margins diverge basally; lateral sides bluntly rounded (as in Fig. 40). Wings as in holotype but pt yellow, Px 10 in right FW, 8 in left FW, 7 in right HW, 7 in left Hw; RP2 branching between Px 3 and 4 but closer to 4 in all wings. Abdomen: color pattern as in male holotype, except for dorsum of S1-7 black with only very narrow pale anterior ring, black on dorsum of S8 reaching posterior margin but very narrowly along posterior fourth of S8; dorsum of S9 with a black triangular spot on each side of anterior margin (as in Fig. 62). Cercus shorter than S10, conical and dark reddish brown; paraprocts yellow. Vulvar spine on S8 well developed, with black tip. Sub-basal plate of ovipositor small and triangular; outer valve of ovipositor with a single row of teeth; tip of ovipositor (excluding stylus) extending slightly beyond posterodorsal margin of S10 (as in Fig. 62).
Dimensions: total length 27.8 mm; abdomen length 22.3 mm; FW 15.3 mm; HW 14.5 mm.
Variation in paratypes. Heteromorphic females with entire posterior area of head dorsum pale (Fig. 9), and thorax entirely pale. Dorsum of S8 and S9 as holotype in some females. Male paratypes Px in FW 8-6, in HW 5-7; RP2 branching between Px 3 and 4 in FW, usually closer to 4, at Px 3 or 4 in HW, usually closer to 4. Female paratypes Px in FW 7-9, in HW 6-7; RP2 branching between Px 3 and 4 in FW, usually closer to 4, at Px 3 or 4 in HW, usually closer to 4. Dimensions: abdomen length in males 18.5-22.2 mm [ mean 20.73 mm; SD 0.79; n 40]; abdomen length in females 20-22.3 mm [ mean 21.29 mm; SD 0.61; n 23]; HW males 11.1-13.5 [ mean 12.54 mm; SD 0.52; n 40]; HW females 13.2-14.75 mm [ mean 13.9 mm; SD 0.5; n 23].
Biology. Adults found at lentic environments (locality data from collections). Larva and habits are unknown.
Distribution (Fig. 89). Venezuela (Barinas, Delta Amacuro, Guárico and Yaracuy states), Colombia (Bolívar, Magdalena and Chocó departments) and Brazil (Amazonas state), from 0 to 870 m a.s.l.
Etymology. Named after my colleague Mark O' Brien, in gratitude for his manifold assistance to students interested in the rich dragonfly collection in UMMZ.
Type material. Holotype m : COLOMBIA. Bolívar: Palermo, 19 i 1917, E.B. & J.H. Williamson col. (UMMZ). Allotype f: COLOMBIA. Magdalena: Aracataca, 9 i 1917, E.B. & J.H. Williamson col. (UMMZ). Paratypes (40 ![]() 23
23![]() ): VENEZUELA. Yaracuy: Nirgua, 28 ii 1920, J.H. & E.B. Williamson col., 1
): VENEZUELA. Yaracuy: Nirgua, 28 ii 1920, J.H. & E.B. Williamson col., 1![]() (RWG). COLOMBIA. Magdalena: same as allotype but 6
(RWG). COLOMBIA. Magdalena: same as allotype but 6![]() 6
6![]() (UMMZ), 2
(UMMZ), 2![]() 2
2![]() (FSCA), 2
(FSCA), 2 ![]() (RWG), 2
(RWG), 2 ![]() 1
1![]() (MLP), 15 xii 1916, 2
(MLP), 15 xii 1916, 2 ![]() (UMMZ), Fundación, S of Aracataca, 12-14 i 1917, E.B. & J.H. Williamson col., 1
(UMMZ), Fundación, S of Aracataca, 12-14 i 1917, E.B. & J.H. Williamson col., 1![]() (RWG), same but 1
(RWG), same but 1![]() (UMMZ), El Banco, 24 I 1917, E.B. & J.H. Williamson col., 1
(UMMZ), El Banco, 24 I 1917, E.B. & J.H. Williamson col., 1![]() 1
1![]() (UMMZ). Bolívar: same as holotype but 5
(UMMZ). Bolívar: same as holotype but 5 ![]() 2
2 ![]() (UMMZ), 6
(UMMZ), 6![]() 1
1![]() (RWG), Zambrano, 20 i 1917, E.B. & J.H. Williamson col., 3
(RWG), Zambrano, 20 i 1917, E.B. & J.H. Williamson col., 3![]() 1
1![]() (RWG), same but 1
(RWG), same but 1 ![]() 1
1 ![]() (UMMZ), near Magangue, 21 i 1917, E.B. & J.H. Williamson col., 1
(UMMZ), near Magangue, 21 i 1917, E.B. & J.H. Williamson col., 1 ![]() 5
5 ![]() (RWG), same but 2
(RWG), same but 2![]() 2
2![]() (UMMZ). Chocó: Sautatá, 6 ii 1918, M.A. Carriker col., 3
(UMMZ). Chocó: Sautatá, 6 ii 1918, M.A. Carriker col., 3 ![]() (UMMZ).
(UMMZ).
Other material examined. BRAZIL. Amazonas: Manaus, 16 vi 1922, J.H. Williamson & J.W. Strohm col., 1 incomplete ![]() , lacking S8-10 (FSCA).
, lacking S8-10 (FSCA).
Remarks. This species most closely approaches H. nepos and was so confused with it in the past, but it differs (alternative characters for H. nepos in square brackets) by medial portion of posterior lobe of prothorax rhomboidal or triangular, roundly (Fig. 37) to angularly (Fig. 38) convex in male [ rectangular with posterior margin linear to concave, Figs. 27-28], forming a triangular lobe in female (Fig. 39) [ slightly sinuate, Fig. 25]; female mesostigmal plates with anterior and posterior margins not parallel sided, and lateral margins bluntly pointed (Fig. 40) [ anterior and posterior margins approximately parallel-sided, and lateral margins bluntly rounded, Fig. 26]; second segment of genital ligula with a few thin hairs on each side (Figs. 53-55) or lacking hairs [ second segment of genital ligula with a dense row of strong hairs on each side (Figs. 50-52)], and bladed projections proximal to flexure in the shape of equilateral triangles, with acutely pointed apex and about as high as wide (Figs. 53- 54)[ bladed projections with bluntly pointed apex and much wider than high (Fig. 50)]; dorsum of male S8 distally pale (Figs. 82- 83) [ dorsum of male S8 distally black, Figs. 80-81]; larger species: male abdomen length 18.5-22.2 mm, mean 20.73 mm, SD 0.79; female abdomen length 20-22.3 mm, mean 21.29 mm, SD 0.61 [ smaller species: male abdomen length 16.5-18.5 mm, mean, 17.74 mm, SD 0.64; female abdomen length 16.9-19.8 mm, mean 18.61 mm, SD 1.05].
The distributions of H. nepos and H. obrieni are apparently allopatric (Fig. 89) along most of their ranges, meeting only in Amazonas state, Brazil. The only male I examined from Manaus, Amazonas state (UMMZ) agrees with H. obrieni in the shape of its pronotal posterior lobe, larger size (HW of 13.5 mm; range in males of H. nepos (n 17): 10.2-12 mm; in males of H. obrieni (n 42): 11.1-13.5 mm), and genital ligula characters (second segment devoid of evident hairs on each side, and bladed projections proximal to flexure with acutely pointed apex and about as high as wide).Since S8-10 are missing, I did not include it in the type series. J. De Marmels (pers. comm.) kindly shared his records (MIZA) and confirmed the presence of H. nepos in Manaus, Amazonas state. The records of H. nepos from French Guiana (Machet, 1989) could correspond to this species (see remarks under H. nepos).
Homeoura sobrina (Schmidt, 1942) comb. nov. (Figs. 11-12, 72, 84-85, 89)
Ischnura sobrina Schmidt, 1942: 237-239, figs. 7a-f (description of male and female, illustrations of male and female head, male S8-10); Rácenis, 1959: 477 (mention from Peru); Garrison, 1991: 12 (synonymic list); Bridges, 1994: VII.220 (list); Steinmann, 1997: 277 (synonymic list); Tsuda, 2000: 37 (synonymic list).
Diagnosis [ based on original description]. Pale postocular spots on dorsum of male head (Fig. 11) rounded and minute (smaller than 1/6 of postocular lobe's width; unique), and no pale postocular bar present (shared with H. chelifera); in heteromorphic female (Fig. 12) entire posterior area of head dorsum pale (shared with all Homeoura species). Male pterothorax with black stripe along metapleural suture (unique); in heteromorphic female there is a black narrow stripe along metapleural suture (unique) beside the medio-longitudinal black stripe on dorsum. Dorsum of male S8-9 pale and sides black (unique), both dorsum and sides of S10 black (Fig. 84). Dorsal and ventral margins of male cercus (Fig. 84) approximately parallel and of equal length in lateral view, so that apical width is approximately the same as basal width, and apex is roundly truncated (shared with H. chelifera); apex slightly bent ventrally. Dorso-posterior margin of male S10 (Figs. 72, 84-85) with u-shaped cleft and two marginal tubercles situated on apex of a projection (unique).
Biology. Larva and habits are unknown.
Distribution (Fig. 89). Peru (Pasco department) at 3000 m a.s.l.
Remarks. Based on its description, this species belongs in Homeoura, displaying diagnostic characters of this genus as regards male cerci morphology. Schmidt (1942) already noted its similarity to 'Ischnura' nepos. He did not describe its genital ligula, and the only specimens so far known comprise the type series originally deposited in Schmidt's collection. Unfortunately, their current location is unknown. Dr. E. Schmidt's collection was inherited by Dr. S. Asahina (Dr. W. Schneider pers. comm.), and Dr. Asahina's collection was in turn donated to the National Museum of Nature and Science in Tokyo, Japan (Dr. H. Ubukata pers. comm.). However, no type specimens from Schmidt are found in this collection (http://www.type.kahaku.go.jp/TypeDB).
Key to species of Argentagrion and Homeoura
1- Posterior lobe of prothorax in both sexes broadly bilobate (Figs. 21, 23) or with a medial triangular apophysis (Fig. 22); postero-dorsal margin of male S10 entire (Figs. 65, 68-70), usually with a small medio-dorsal apophysis (Figs. 69, 70); inner surface of genital ligula proximal to flexure smooth (Figs. 41, 43); distal portion of male cercus bent ventrally at a right angle (Figs. 65-70) …………….. Argentagrion .................................... 2
1'- Posterior lobe of prothorax trilobate with a differentiated medial process, which is trapezoidal (Figs. 33, 37-38) or rectangular (Figs. 27-30) in male, triangular (Figs. 34-35, 39), rectangular (as in Fig. 30) or barely bilobate(Fig. 31) in female; postero-dorsal margin of male S10 with a u-shaped medial cleft with a tubercle on each side (Figs. 71-85); inner surface of male genital ligula proximal to flexure provided with a pair of sclerotized projections shaped as triangular blades (Figs. 46-47, 49-50, 53-54, 56, bl); male cercus approximately horizontal with a ventrally directed laminar branch at base (Figs. 71-85 ……. Homeoura … 3
2- Posterior lobe of prothorax of both sexes broadly bilobate (Figs. 21, 23); male cercus about twice as long as wide in dorsal view (Fig. 66); dorsal branch of male paraproct rounded (Fig. 67); dorsum of S3-8 mostly black (Fig. 58); larger species (HW 12.5-15mm; abdomen 19-21.5 mm)........................ ................. Argentagrion ambiguum Ris
2'- Posterior lobe of prothorax with a medial triangular apophysis (Fig. 22; female unknown); male cercus about as long as wide in dorsal view (Fig. 69); dorsal branch of male paraproct ending in a sclerotized tooth (Fig. 70); dorsum of S3-6 with black restricted to narrow medio-longitudinal stripe, limited to basal half on S7, and dorsum of S8 entirely pale (Fig. 59); smaller species (HW 10.4 mm; abdomen 17.5 mm) [ female unknown, but probably similar to male in more limited extension of black pattern and smaller size] ..................... Argentagrion silviae Bulla
3- Pale postocular spots of male (Fig. 11) minute (less than 1/6 of postocular lobe's width); complete black stripe on male metapleural suture, and narrow black stripe on metapleural suture of heteromorphic females; lateral sides of male S9 black (Fig. 84) [ based on original description] ................ ............... Homeoura sobrina (Schmidt)
3'- Pale postocular spots of male (Figs. 3- 4, 7-8, 10) large (about as wide as 1/3 of postocular lobe's width); no complete black stripe on male metepisternalmetepimeral suture, and no narrow black stripe on mesepimeralmetepisternal suture of heteromorphic females; lateral sides of male S9 pale (Figs. 64, 77-78, 80, 82) .................... 4
4- Pt approximately quadrangular, with costal side the shortest (Figs. 14, 19-20) ....................................................... 5
4'- Pt approximately rhomboidal, with costal side the longest (Figs. 16-17) ... 6
5- Medial portion of posterior lobe of prothorax approximately rectangular, with posterior margin slightly concave (Fig. 28) to linear (Fig. 27) in male, and slightly sinuate in female (Fig. 25); female mesostigmal plates with anterior and posterior margins approximately parallel sided, and lateral margins bluntly rounded (Fig. 26); second segment of genital ligula with a dense row of strong hairs on each side (Figs. 50-52), and bladed projections proximal to flexure with bluntly pointed apex and much wider than high (Fig. 50); dorsum of male S8 entirely black (Figs. 80, 81); smaller species (males' abdomen length 16.5-19.1 mm, mean, 17.82 mm, SD 0.7; females' abdomen length 16.9-19.8 mm, mean 18.61 mm, SD 1.05) ...................... Homeoura nepos (Selys)
5'- Medial portion of posterior lobe of prothorax rhomboidal or triangular, with posterior margin roundly (Fig. 37) to angularly (Fig. 38) convex in male, forming a triangular lobe in female (Fig. 39); female mesostigmal plates with anterior and posterior margins not parallel sided, and lateral margins bluntly pointed (Fig. 40); second segment of genital ligula with a few thin hairs on each side (Figs. 53-55) or lacking hairs, and bladed projections proximal to flexure in the shape of equilateral triangles, with acutely pointed apex and about as high as wide (Figs. 53-54); dorsum of male S8 distally pale (Figs. 82-83); larger species (males' abdomen length 18.5-22.2 mm, mean 20.73 mm, SD 0.79; females' abdomen length 20-22.3 mm, mean 21.29 mm, SD 0.61) ....... Homeoura obrieni sp. nov.
6- Medial portion of posterior lobe of prothorax in both sexes is approximately rectangular, with posterior margin linear (Figs. 29-30), to narrowly bilobate in some females (Fig. 31); lateral margins of female mesostigmal plates truncated (Fig. 32); in lateral view dorsal and ventral margins of male cercus approximately parallel and of equal length, so that apical width is approximately the same as basal width and apex is roundly truncated (Figs. 71, 76), cerci distinctly longer than S10 (Figs. 76-77) ................................ .................. Homeoura chelifera (Selys)
6'- Medial portion of posterior lobe of prothorax approximately rhomboidal with posterior margin convex in male (Fig. 33, and as in Fig. 38), and triangular in female (Figs. 34-35); lateral margins of female mesostigmal plates bluntly pointed (Fig. 36); in lateral view dorsal and ventral margins of male cercus diverging apically, and dorsal margin longer than ventral, so that apical width is larger than basal width, and apex is obliquely truncated dorsoventrally (Figs. 75, 78), cerci about as long as or shorter than S10 (Figs. 78-79) ....................... Homeoura lindneri (Ris)
DISCUSSION
As pointed out by Bulla (1971b), Rácenis' (1958) rediagnosis of Argentagrion (venation reduced, 6-10 postnodals, male paraprocts shorter than half of cerci, and smaller size: abdomen 18-23 mm; HW 12.5-15 mm) will not allow for the inclusion of A. silviae (with paraprocts almost as long as cerci), and both A. ambiguum and A. silviae differ in several characters from H. cheliferum and H. lindneri, precluding their placement in the same genus. Although he did not study all species involved, when redefining Cyanallagma De Marmels (1989, 1997) advocated the same placement I am proposing here for these species: Argentagrion ambiguum, Homeoura chelifera, H. nepos and H. lindneri. Unfortunately these observations went unnoticed, and the treatment of all species as congeneric by Rácenis (1958) and Davies & Tobin (1984) was instead followed (Garrison, 1991; Steinmann, 1997; Muzón & von Ellenrieder, 1998; Costa et al. 2000; Tsuda, 2000; Lencioni, 2006).
In their cladistic analysis designed to test sub-familiar divisions within Coenagrionidae, O'Grady & May (2003) included one species, each of Argentagrion (A. ambiguum as Homeoura ambigua) and Homeoura (H. lindneri as Ischnura sobrina). These two species clustered together only in the analysis based solely on sub-familiar characters used by Fraser (1957) and Davies & Tobin (1984), but not in their analyses when based on all characters and on non-continuous and unambiguously defined characters (O'Grady & May, 2003: 2820-2821), supporting my hypothesis that Homeoura sensu lato (including all species treated here under Argentagrion and Homeoura) constitutes a polyphyletic assemblage.
The last larval instar of only one species is known satisfactorily for each Argentagrion and Homeoura, differing by the shape of caudal lamellae: in A. ambiguum, the base of the caudal lamella is dark with nondiscernible tracheae, and primary and secondary tracheae on the distal portion are equally pigmented (Fig. 86), whereas in H. chelifera, the base of the caudal lamella is not differentiated from the distal portion: there are three diffuse dark spots along its midline, and secondary tracheae are much darker than primary (Fig. 87). Whether these differences are of generic value will be established only when larvae of more species are known.
ACKNOWLEDGEMENTS
I thank Rosser Garrison, Jürg De Marmels, Javier Muzón and Kenneth Tennessen for critically reading the manuscript and for their valuable suggestions, Rosser Garrison (RWG), Javier Muzón (MLP), Mark O'Brien (UMMZ), Sidney W. Dunkle (SWD), Bill Mauffray (FSCA) and Dennis R. Paulson (DRP) for the loan and gift of material, Rosser Garrison (RWG) for his illustrations of the holotype of Agrion nepos that accompany this paper, and Jürg De Marmels (MIZA) for confirming the identity of Homeoura specimens and sharing unpublished distribution records of Homeoura chelifera, H. nepos and H. obrieni in his collection. This study was supported by the Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina (CONICET) and PICT 2006-1378 (FONCyT).
LITERATURE CITED
1. BRIDGES, C. A. 1994. Catalogue of the family-group, genusgroup, and species-group names of the Odonata of the world (Third Edition). Urbana, xlvi+905 pp.
2. BULLA, L. A. 1970. Contribución al conocimiento de los Zygoptera Argentinos. I. Las ninfas de Telebasis willinki Fraser y Argentagrion ambiguum (Ris.) (Insecta, Odonata). Rev. Soc. Entomol. Argent. 32(1- 4): 91-98.
3. BULLA, L. A. 1971a. Contribución al conocimiento de los Odonata Zygoptera de la provincia de Buenos Aires. Tesis Doctoral Universidad Nacional de La Plata, 223 pp.
4. BULLA, L. A. 1971b. Consideraciones sobre el género Argentagrion Fraser, 1947 con la descripción de una nueva especie (Odonata, Coenagriidae). Rev. Soc. Entomol. Argent. 33(1-4): 49-55.
5. BULLA, L. A. 1973. Revisión de dos especies Argentinas del género Cynallagma Kennedy (Odonata, Coenagriidae). Rev. Soc. Entomol. Argent. 34(1-2): 95-105.
6. CALVERT, P. P. 1909. Contributions to a knowledge of the Odonata of the Neotropical region, exclusive of Mexico and Central America. Ann. Carnegie Mus. 6: 73-280.
7. CALVERT, P. P. 1948. Odonata from Pirassununga (Emas), state of São Paulo, Brazil: ecological and taxonomic data. Bol. Mus. Nac., N. S. 87: 1-34.
8. COSTA, J. M., A. B. M. MACHADO, F. A. A. LENCIONI & T. C. SANTOS. 2000. Diversidade e distribuição dos Odonata (Insecta) no estado de São Paulo, Brasil: Parte I - Lista das espécies e registros bibliográficos. Publ. Avul. Mus. Nac., Rio de Janeiro 80: 1-27.
9. CRUZ, L. F. 1986. Contribución a los estudios taxonómicos de Odonata-Zygoptera de Colombia: descripción de una nueva especie de Cianallagma (Odonata: Coenagrionidae). Caldasia 14(68-70): 743-747.
10. DAVIES, D. A. L. & P. TOBIN. 1984. The dragonflies of the world: a systematic list of the extant species of Odonata. Volume 1. Zygoptera, Anisozygoptera. S. I. O. Rapid Comm. (Suppl.), Utrecht, 3: ix + 127 pp.
11. DE MARMELS, J. 1989. Notes on Acanthagrion acutum Ris, Enallagma occultum Ris, and E. ovigerum Calvert (Zygoptera: Coenagrionidae). Odonatologica 18(3): 245-252.
12. DE MARMELS, J. 1990. An updated checklist of the Odonata of Venezuela. Odonatologica 19(4): 333-345.
13. DE MARMELS, J. 1997. New and little-known species of Cyanallagma Kennedy, 1920 from the Andes and from Pantepui (Zygoptera: Coenagrionidae). Odonatologica 26: 135-157.
14. DE MARMELS, J. 2007. Tepuibasis gen. nov. from the Pantepui region of Venezuela, with descriptions of four new species and with biogeographic, phylogenetic and taxonomic considerations on the Teinobasinae (Zygoptera: Coenagrionidae). Odonatologica 36(2): 117-146.
15. DE MARMELS, J. & R. W. GARRISON. 2005. Comments on the Genus Leptagrion Selys, 1876 in Venezuela and on other poorly known species, with a description of Bromeliagrion gen. nov. (Zygoptera: Coenagrionidae). Can. Entomol. 137(3): 257 273.
16. DONNELLY, T. W. & D. A. PASTOR ALAYO. 1966. A new genus and species of damselfly from Guatemala and Cuba (Odonata: Coenagrionidae). Florida Ent. 49(2): 107-114.
17. FRASER, F. C. 1947. The Odonata of the Argentine Republic I. Acta Zool. Lilloana 4: 427-462.
18. FRASER, F. C. 1948. The Odonata of the Argentine Republic II. Acta Zool. Lilloana 5: 47-67.
19. FRASER, F. C. 1957. A reclassification of the order Odonata. Roy. Zool. Soc. N.S.W., Sydney, 133 pp.
20. GARRISON, R. W. 1991. A synonymic list of the New World Odonata. Argia 3(2): 1-30.
21. GLOGER, H. 1967. Sobre Acanthagrion (Odonata, Coenagrionidae) y géneros próximos. Acta Zool. Lilloana 21: 45-58.
22. KENNEDY, C. H. 1916. Notes on the penes of Zygoptera (Odonata) No.1 Species limits in the genus Acanthagrion. Ent. News 27: 323-330.
23. KENNEDY, C. H. 1920. Forty-two hitherto unrecognized genera and subgenera of Zygoptera. Ohio J. Science 21: 83-88.
24. KIAUTA, B. 1988. Odonatological Abtract 6314. Odonatologica 17(4): 437-438.
25. KIRBY, W. F. 1890. A synonymic catalogue of Neuroptera Odonata, or dragonflies, with an appendix of fossil species. Gurney & Jackson, London, ix + 202 pp.
26. LENCIONI, F. A. A. 2006. Damselflies of Brazil. An illustrated identification guide. 2 - Coenagrionidae. All Print Editora, São Paulo, viii + 419 pp.
27. MACHET, P. 1989. Contribution à l'étude des odonates de Guyane Française. 1. Zygoptera. Opusc. zool. flumin. 40: 1-16.
28. MAY, M. 2002. Phylogeny and taxonomy of the damselfly genus Enallagma and related taxa (Odonata: Zygoptera: Coenagrionidae). Syst. Ent. 27: 387-408.
29. MUZÓN, J. & N. VON ELLENRIEDER. 1998. Odonata, pp. 14-21. In: Morrone, J.J. & S. Coscarón (eds.), Biodiversidad de Artrópodos argentinos. Una perspectiva biotaxonómica. Ediciones Sur, La Plata, Argentina, 599 pp.
30. MUZÓN, J., P. PESSACQ & L. RAMOS. 2007. Odonata type specimens preserved in the Museo de La Plata. Odonatologica 36(3): 301-306.
31. NEEDHAM, J. G. 1904. New dragonfly nymphs in the United States National Museum. Proceedings of the United States National Museum 27: 685-720.
32. O'GRADY, E. W. & M. L. MAY. 2003. A phylogenetic reassessment of the subfamilies of Coenagrionidae (Odonata: Zygoptera). J. Nat. Hist. 37: 2807-2834.
33. RÁCENIS, J. 1958. Los Odonatos Neotropicales en la colección de la Facultad de Agronomía de la Universidad Central de Venezuela. Acta Biol. Venez. 2(19): 179-226.
34. RÁCENIS, J. 1959. Lista de los Odonata del Peru. Acta Biol. Venez. 2(34): 467-522.
35. RIEK, E. J. & J. KUKALOVÁ-PECK. 1984. A new interpretation of dragonfly wing venation based upon Early Upper Carboniferous fossils from Argentina (Insecta: Odonatoidea) and basic character states in pterygote wings. Can. J. Zool. 62: 1150-1166.
36. RIS, F. 1904. Odonaten, 44 pp. In: Friederichsen & Co., Hamburg Magallaenischen Sammelreise 1892/93, Hamburg.
37. RIS, F. 1913. Neuer Beitrag zur Kenntnis der Odonatenfauna von Argentina. Mém. Soc. Belg. 22: 55-102 (1-48 separate).
38. RIS, F. 1918. Libellen (Odonata) aus der Region der amerikanischen Kordilleren von Costarica bis Catamarca. Arch. Naturgesch. A(9): 1-197.
39. RIS, F. 1928. Die Ausbeute der Deutschen Chaco- Expedition 1925-26. Odonata. Konowia 7(1): 40-49.
40. RODRIGUES CAPÍTULO, A. 1992. Los Odonata de la República Argentina (Insecta). Fauna de agua dulce de la Republica Argentina 34(1): 1-91.
41. RODRIGUES CAPÍTULO, A., L. M. MOLA & S. S. AGOPIAN. 1991. Species catalogue and chromosomal data of Odonata from Argentina. Rev. Soc. Entomol. Argent. 49(1-4): 59-72.
42. SANTOS, N. D. 1956. Contribuição ao conhecimento da fauna do Distrito Federal. XLVII - Enallagma cheliferum (Selys, 1876) nova combinação (Coenagriidae: Odonata). An. Acad. Bras. Cs. 28(4): 571-576.
43. SCHMIDT, E. 1942. Odonata nebst Bemerkungen über die Anomisma und Chalcopteryx des Amazonas- Gebiets. In: E. Titschack (ed.), 1941-1942 Beiträge zur Fauna Perus nach der Ausbeute der Hamburger Sudperu Expedition 1936 2: 225-276.
44. SELYS LONGCHAMPS, E. de. 1876. Synopsis des Agrionines, 5me légion: Agrion (suite). Le genre Agrion. Bull. Acad. r. Belg. (2)41: 247-322,496-539, 1233-1309.
45. STEINMANN, H. 1997. World catalogue of Odonata. Volume I. Zygoptera. In: Wermuth, H. & M. Fischer (eds.). Das Tierreich. The Animal Kingdom. Eine zusammenstellung und Kennzeichnung der rezenten Tierformen, Walter de Gruyter, Berlin, 110: xxi + 500 pp.
46. ST. QUENTIN, D. 1960. Zur Kenntnis der Agrioninae (Coenagrioninae) Südamerikas (Odonata). Beitr. Neotr. Fauna 2(1): 45-64.
47. TSUDA, S. 2000. A distributional list of world Odonata 2000. Privately published, Osaka, vi + 430 pp.
48. VON ELLENRIEDER, N. 2000. Species composition and temporal variation of odonate assemblages in the subtropical-pampasic ecotone, Buenos Aires, Argentina. Odonatologica 29(1): 17-30.
49. VON ELLENRIEDER, N. & R. W. GARRISON. 2007. Libélulas de las Yungas (Odonata). Una Guía de Campo para las especies de Argentina / Dragonflies of the Yungas. A Field Guide to the Species from Argentina. Pensoft Series Faunistica 67, Pensoft Publishers, Sofia-Moscow, iv + 116 pp.
50. VON ELLENRIEDER N. & F. LOZANO. 2008. Blues for the red Oxyagrion: a redefinition of the genera Oxyagrion Selys, 1876 and Acanthagrion Selys, 1876 (Odonata: Coenagrionidae). IJO 11(1): 95-113. [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ] [ Links ]
Recibido: 1-02-2008;
Aceptado: 26-02-2008

