










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. v.68 n.3-4 Mendoza jul./dic. 2009
TRABAJOS CIENTÍFICOS
Visitantes florales nocturnos del girasol (Helianthus annuus, Asterales: Asteraceae) en la Argentina
Nocturnal floral visitors of sunflower (Helianthus annuus, Asterales: Asteraceae) in Argentina
Torretta Juan P*, Fernando Navarro** y Diego Medan*
*Cátedra de Botánica Agrícola, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453, C1417DSE, Buenos Aires, Argentina; e-mail: torretta@agro.uba.ar; diemedan@agro.uba.ar
**Instituto Superior de Entomología "Dr. Abraham Willink", Facultad de Ciencias Naturales y Instituto Miguel Lillo, Universidad Nacional de Tucumán, Miguel Lillo 205, 4000, Tucumán, Argentina; e-mail: fnavarropaz@yahoo.com.ar
RESUMEN: El girasol (Helianthus annuus) es un cultivo oleaginoso, polinizado por Apis mellifera L. y otras abejas en distintas regiones del mundo. Sin embargo, sus flores también son visitadas por insectos de actividad nocturna. Durante tres campañas agrícolas, se determinó la diversidad de los visitantes nocturnos de capítulos de girasol, en cinco sitios de Argentina. También se estudió el comportamiento de forrajeo de los principales visitantes y la variación de la receptividad estigmática a lo largo del día, con el fin de establecer si estos visitantes contribuyen a la polinización. Al menos 67 especies o morfoespecies pertenecientes a cuatro órdenes de visitantes nocturnos fueron colectadas. El orden más rico y abundante fue Lepidoptera (44 especies o morfoespecies, cinco familias), seguido por Coleoptera (18 especies o morfoespecies, nueve familias), Orthoptera (tres morfoespecies, una familia) y Blattaria (dos especies, una familia). Los lepidópteros forrajearon exclusivamente por néctar, mientras que los individuos de los demás órdenes consumieron polen y/o partes florales. El estigma se encontró receptivo durante las horas de luz, con una receptividad máxima al mediodía (12:00 - 14:00). Llamativamente, las flores del girasol son visitadas por mayor número de polillas que de abejas. Debido a que las polillas consumen néctar y potencialmente transportan polen entre flores, en un momento del día en que los estigmas se encuentran menos receptivos, es improbable que polinicen efectivamente el cultivo.
PALABRAS CLAVE: Noctuidae; Receptividad estigmática; Visitantes florales nocturnos.
ABSTRACT: Sunflower (Helianthus annuus) is an oilseed crop pollinated by Apis mellifera L. and other diurnal bees in different regions of the world. However, their flowers are also visited by insects active at night. During three agricultural years, the diversity of nocturnal visitors to sunflower heads was assessed in five different sites in Argentina. The foraging behavior of the main visitors as well as the stigmatic receptivity variations along the day were also studied in order to determine whether or not these visitors contribute to pollination. At least 67 species or morpho-species belonging to four nocturnal visitor orders were recorded. The richest and most abundant order was Lepidoptera (44 species or morpho-species, five families), followed by Coleoptera (18 species or morpho-species, nine families), Orthoptera (three morpho-species, one family) and Blattaria (two species, one family). Lepidopterans foraged exclusively for nectar while species in other orders consumed pollen and/or floral parts. However, stigma receptivity was greater during the day, peaking at midday (12:00 - 14:00). Unexpectedly, moth species widely outnumbered bee species as sunflower visitors. Since moths potentially transfer pollen at a time of the day when stigmas are least receptive, it is unlikely that they do effectively pollinate sunflowers.
KEY WORDS: Noctuidae; Stigmatic receptivity; Nocturnal flower visitors.
Recibido: 24-IX-2009;
Aceptado: 6-XI-2009
INTRODUCCIÓN
La morfología y coloración de las flores del girasol (Helianthus annuus) sugieren que ésta es una especie polinizada por insectos diurnos. De hecho, este importante cultivo oleaginoso es polinizado principalmente por la abeja doméstica (Apis mellifera L.) en diversas regiones del mundo (McGregor, 1976; Free, 1993; Delaplane & Mayer, 2000). Otras numerosas especies de abejas también visitan y polinizan las flores de este cultivo durante el día (Hurd et al., 1980). Torretta et al. (ms. enviado) observaron 32 especies de abejas que visitan flores de girasol en la Argentina. Sin embargo, Delisle et al. (1989) reportaron que el aroma emitido por el polen del girasol es atractivo para la polilla nocturna Homoeosoma electellum (Hulst) (Pyralidae), ya que actúa como un estimulante para la oviposición.
La observación de Delisle et al. (1989) sugiere, por lo tanto, que el girasol puede ser polinizado por insectos nocturnos además de los diurnos, tal como sucede en Catalpa speciosa (Bignoniaceae) (Stephenson & Thomas, 1977) y Hancornia speciosa (Apocynaceae) (Darrault & Schlindwein, 2005). En estas especies, al igual que en el girasol, la longevidad floral es mayor que un día y la oferta de recompensas tiene lugar durante todo el día. Torretta et al. (2005) observaron numerosas especies de insectos nocturnos (principalmente, lepidópteros) que visitan flores de girasol, en dos sitios de la provincia de Buenos Aires (Gral. Villegas [S 34° 43´ O 63° 12´] y Balcarce [S 37º 46' O 58º 18']) en las primeras horas de la noche (21:00-24:00). Aparte de este trabajo exploratorio y preliminar, y de informes aislados (Lehman et al., 1973; Del Socorro & Gregg, 2001) hasta el momento no existe ningún relevamiento sistemático sobre los visitantes florales nocturnos en ningún sitio donde se cultive comercialmente el girasol. Estos hechos indujeron a extender el área de estudio y a intentar dilucidar si los visitantes florales de actividad nocturna, se comportan de manera mutualista o antagónica en relación a la polinización (y posterior producción) del cultivo.
En este trabajo, se intenta describir el elenco de los visitantes nocturnos del girasol en un área vasta del cultivo en la Argentina, y responder las siguientes preguntas: a) ¿cuál es la principal recompensa floral por la que éstos forrajean?, b) ¿son eficaces en el transporte de polen?, c) ¿en qué momento del día se encuentra receptivo el estigma del girasol? y d) ¿contribuyen a la polinización de este cultivo oleaginoso?
MATERIAL Y MÉTODOS
Sitios de muestreo
Durante las campañas agrícolas 2004-2005, 2005-2006 y 2006-2007 se relevaron cinco sitios (Tabla I, de aquí en adelante los sitios serán nombrados por su abreviatura). En cada sitio de muestreo se eligió un lote comercial del cultivo en el cual la mayoría de las plantas ya se encontraban florecidas (estadio R-5 de Schneiter & Miller, 1981). El tamaño de los lotes sembrados con girasol varió entre 25 y 50 ha. (Tabla I). Dada la magnitud del muestreo no fue posible relevar todos los sitios durante la misma campaña agrícola, y las posibles diferencias interanuales en condiciones ambientales no pudieron tenerse en cuenta. Los lotes fueron revisados dos veces en un lapso de tres días con condiciones climáticas semejantes, durante las primeras horas de la noche (21:00-24:00). En cada visita se recorrió cada lote dos veces, se comenzó la primera vez a las 21:00 y la segunda a las 23:00 (excepto en BA y RO, en donde se realizaron solamente tres recorridos: dos a las 21:00 y uno a las 23:00). En total se acumularon 20,30 hs de captura entre todos los sitios (Tabla I).
Tabla I. Localización de los sitios estudiados, períodos en que fueron relevados, esfuerzo de muestreo (horas-hombre de captura) y tamaño de los lotes.

Especie vegetal
La variedad cultivada del girasol (Asteraceae, Heliantheae, H. annuus var. macrocarpus) es una planta no ramificada que tiene sólo un capítulo, que alcanza mayor tamaño que en la variedad silvestre. El capítulo posee dos tipos de flores. Las flores marginales, liguladas, son amarillas, llamativas y estériles. La corola de estas flores absorbe fuertemente la luz UV y contribuye posiblemente en la atracción de los polinizadores (Sammataro et al., 1983, citado en Seiler, 1997).
Las flores del disco son perfectas, están dispuestas en arcos que radian desde el centro del capítulo, y son las que forman frutos (Seiler, 1997). Entre uno y cuatro círculos de flores del disco abren por día, de manera centrípeta, la duración total de la floración del capítulo es de 10 a 15 días. La corola de las flores del disco está formada por cinco pétalos casi totalmente soldados (excepto en el ápice). Dentro del tubo formado por la corola se encuentra el androceo sinantéreo. Encerrado por el tubo formado por las anteras, está el estilo. En la base del estilo de las flores del disco se encuentran los nectarios florales (Seiler, 1997). Las anteras liberan el polen antes de que el estigma se torne receptivo (protandria). Al crecer, el estilo empuja hacia el exterior (a modo de pistón) el polen alojado en el interior del tubo anteral. El polen queda adherido a la zona no receptiva de las ramas estigmáticas. Posteriormente, los filamentos estaminales pierden turgencia y se introducen en la corola. Las ramas estigmáticas se expanden totalmente entre la noche y la mañana del día siguiente. Una vez que la superficie estigmática receptiva es expuesta, puede ocurrir la polinización (Seiler, 1997). En girasoles cultivados, la secreción de néctar ocurre principalmente (aunque no exclusivamente) durante las horas de luz del día, es decir, durante la fase estaminada (McGregor 1976; Fell 1986), aunque Smidichev (1977, citado en Neff & Simpson, 1990) indica que la producción de néctar continúa durante la noche.
Relevamiento de los visitantes florales
La captura de los visitantes florales se realizó caminando lentamente a lo largo de los surcos, mientras se iluminaban los capítulos con linternas de cabeza. Cada recorrida era realizada por dos o tres personas, separadas entre sí por aprox. 20 m. La duración de cada recorrida varió entre 25 y 40 minutos lo que dependía del tamaño del lote (longitud del recorrido: 500-800 m). Los individuos colectados se sacrificaron en frascos con acetato de etilo y se prepararon según las técnicas convencionales. Las determinaciones de los lepidópteros nocturnos se realizaron en laboratorio. Los ejemplares de Coleoptera y Blattaria capturados fueron determinados por especialistas (ver Agradecimientos). Los mismos se encuentran depositados en la Colección Entomológica de la Cátedra de Botánica Agrícola de la Facultad de Agronomía de la Universidad de Buenos Aires.
Descripción del comportamiento del visitante floral
Se realizaron observaciones de los principales visitantes florales nocturnos del girasol, con especial referencia al comportamiento de forrajeo y a la recompensa buscada por los insectos, en todos los sitios muestreados excepto BA. Las observaciones del comportamiento de los visitantes nocturnos se realizaron entre las 21:00 y las 23:00 en días en que no se realizaban las capturas.
Determinación de las cargas polínicas corporales
Para determinar, si los lepidópteros nocturnos transportaban granos de polen de girasol en SU y HO, se capturaron individuos mientras libaban en flores de girasol, se sacrificaron y mantuvieron aislados (para evitar contaminación de polen entre individuos). En el laboratorio, se cortó y montó la espiritrompa de cada individuo estudiado en un portaobjetos con gelatinaglicerina coloreada con safranina. Luego, bajo microscopio se verificó la presencia/ausencia de granos de polen de girasol. En caso de presencia, se contó la totalidad de granos de polen. Para comparar el número de polillas que transportaban granos de polen de girasol en la espiritrompa, se realizó un análisis de datos categóricos (Zar, 1996). Para realizar una prueba de Bondad de Ajuste (χ2), se agruparon los individuos en categorías: carga polínica baja (1-100 granos de polen), carga polínica media (101-200 granos de polen) y carga polínica alta (>201 granos de polen). Esta prueba se realizó bajo la hipótesis nula de igualdad entre las tres categorías y valor-p de 0,05.
Receptividad estigmática del girasol
Para conocer la variación de la receptividad estigmática a lo largo del día, se evaluaron estigmas de girasol, para lo que se empleó el papel reactivo Peroxtesmo (Dafni & Motte-Maués, 1998). La prueba de Peroxtesmo se utiliza para comprobar la presencia de peroxidasas en estigmas. Cuando los estigmas están receptivos, las tiras de papel Peroxtesmo reaccionan tornándose azules. La intensidad de la coloración depende de la concentración de peroxidasas presentes en el estigma. Subjetivamente, a cada estigma tratado con Peroxtesmo se le asignó un valor de 0 (estigma no receptivo, sin coloración en la tira de Peroxtesmo), 1 (estigma muy poco receptivo, tinte azulado pálido), 2 (estigma medianamente receptivo, tinte azulado fuerte) o 3 (estigma fuertemente receptivo, color azul fuerte). Los valores asignados a los estigmas tratados en cada horario se promediaron para obtener un "índice de receptividad estigmática" a lo largo del día. Los estigmas analizados bajo lupa binocular, pertenecían a flores previamente emasculadas para evitar falsos positivos, ya que los granos de polen también reaccionan con Peroxtesmo. La receptividad fue medida en enero de 2006, en muestras de 6-15 estigmas tomados de flores de 2-5 capítulos, cada dos horas desde las 6:00 hasta las 20:00, y cada hora entre las 20:00 y las 23:00, en un cultivo experimental en la Facultad de Agronomía de la Universidad de Buenos Aires. Para analizar si hubo diferencias en el "índice de receptividad estigmática" a lo largo del día, se realizó una tabla de contingencia utilizando una prueba de Chi-cuadrado (χ2). Para este análisis, se agruparon los horarios en cuatro categorías: mañana (6:00-10:00); mediodía (12:00-14:00), tarde (16:00-18:00) y noche (20:00-23:00); y el valor del índice en tres categorías: receptividad nula (valor 0), receptividad media (valores 1 y 2) y receptividad máxima (valor 3). Las categorías de receptividad estigmática se realizaron con el fin de disminuir las casillas de la tabla de contingencia con valores debajo de 5.
RESULTADOS
Relevamiento de los visitantes florales
En los cinco sitios relevados se colectaron 637 visitantes florales nocturnos, pertenecientes a cuatro órdenes de insectos: Lepidoptera, Coleoptera, Blattaria y Orthoptera (Tablas II y III). De ellos, 577 fueron lepidópteros nocturnos de las familias Noctuidae, Pyralidae, Crambidae, Geometridae y Sphingidae. De los 60 individuos restantes, 45 fueron escarabajos, diez cucarachas y cinco langostas.
Tabla II. Lepidópteros nocturnos capturados sobre capítulos de girasol, en los cinco sitios relevados durante las campañas agrícolas 2004-2005, 2005-2006 y 2006-2007.

Tabla III. Visitantes florales nocturnos no lepidópteros capturados sobre capítulos de girasol, en los cinco sitios relevados durante las campañas agrícolas 2004-2005, 2005-2006 y 2006-2007.

Los lepidópteros nocturnos más abundantes pertenecieron a la familia Noctuidae (510 ejemplares; 34 especies), mientras que las familias Crambidae (32 ejemplares; dos especies), Pyralidae (28 ejemplares; tres morfoespecies), Geometridae (cinco ejemplares; tres morfoespecies) y Sphingidae (dos ejemplares de Hyles lineata (Fabricius)) estuvieron menos representadas.
Los noctuidos representaron el 88,4% de los lepidópteros nocturnos y el 80% de todos los visitantes florales nocturnos del girasol. El género más abundante fue Feltia Walker, con 155 individuos pertenecientes a dos especies, seguido por Pseudaletia Franclemont (97 individuos y dos especies) y Spodoptera Guenée (67 individuos y tres especies). Las especies más comunes fueron: Feltia deprivata (Walker) 96 individuos, Spodoptera frugiperda (J.E. Smith) 63, Feltia subterranea (Fabricius) 59, Pseudaletia unipuncta (Haworth) 56, y P. adultera (Schaus) 41. Estas cinco especies corresponden a 315 individuos (54,6% de los lepidópteros nocturnos capturados). Otras especies de noctuidos comunes fueron: Rachiplusia nu (Guenée) 33 individuos, Peridroma saucia (Hübner) 29, Faronta albilinea (Hübner) 23 y Pseudoleucania bilitura (Guenée) 20.
Las familias Crambidae y Pyralidae estuvieron representadas por un bajo número de ejemplares. La especie más abundante de Crambidae fue Achyra bifidalis (Fabricius) con 30 individuos, mientras que Nomophila indistinctalis (Walker), solamente por dos individuos. Entre los Pyralidae, la especie Pyralidae sp. 1 fue la más abundante, con 17 ejemplares capturados.
En cuanto al área de distribución de las especies, solamente tres especies (Feltia subterranea, Rachiplusia nu y Achyra bifidalis) estuvieron presentes en todos los sitios muestreados (Tabla II). Tres especies de noctuidos (Faronta albilinea, Peridroma saucia y Pseudaletia unipuncta) estuvieron en cuatro sitios (Tabla II). Numerosas especies, representadas sólo en un sitio y con un número pequeño de individuos (Tabla II).
Cuarenta y cinco individuos fueron coleópteros de nueve diferentes familias (Tabla III): Elateridae (Conoderus alfredoi Guzmán de Tomé y cinco morfoespecies más), Carabidae (Metius circumfusus (Germar)), Scarabaeidae (Anomala testaceipennis Blanchard, Euphoria lurida (Fabricius) y Bothinus sp.), Chrysomelidae (Amblycerus dispar Sharp y Spintherophyta sp.), Coccinelidae (Eriopis connexa (Germar)), Curculionidae (Naupactus verecundus Hustache), Melyridae (Astylus atromaculatus Blanchard), Nitidulidae (Lobiopa sp.) y Tenebrionidae (Hylithus tentyroides Lacordaire y Epitragus sp.). En general, excepto Astylus atromaculatus que fue muy abundante en RO, los coléopteros estuvieron presente en número bajo. Además, en SU se capturaron nueve individuos de la cucaracha Ischnoptera bilunata Saussure y en HO un individuo de I. argentina Hebard (Blatellidae). En SU y RO se capturaron cinco individuos de tres especies de ortópteros (Tabla III).
Comportamiento de los visitantes florales
Tanto las polillas como los demás insectos nocturnos se movilizaban poco dentro del capítulo o entre capítulos. Las polillas libaban posadas en los mismos, pero el cuerpo quedaba separado de las flores. Individuos de algunas especies, como Rachiplusia nu, aleteaban constantemente mientras libaban néctar. El largo de la espiritrompa de todos los lepidópteros observados superaba el tamaño del tubo floral, por lo que el animal no introducía su cabeza en la flor. De tal manera, las polillas no contactaban las anteras y/o estigmas con sus cabezas. Incluso, un mismo individuo podía libar de varias flores sin necesidad de desplazarse por el capítulo, moviendo solamente su aparato bucal. Sin embargo, mientras se alimentaban del néctar del girasol (tanto en flores en fase masculina como en flores en fase femenina), las polillas tocaban las partes florales con los palpos labiales y con las antenas que son sensoriales y detectan olores relacionados con el apareamiento y la alimentación (Scout, 1986; Krenn et al., 2005).
Si bien no se realizó un muestreo exhaustivo de los estadios preimaginales ni adultos que no se encontraban forrajeando en las flores del cultivo, durante las observaciones comportamentales no se observaron polillas copulando, ni hembras oviponiendo en las flores del girasol, así como tampoco se hallaron larvas comiendo polen. A diferencia de la mayoría de las polillas, los dos ejemplares de esfíngidos capturados libaban néctar en vuelo sostenido frente al capítulo, comportamiento bien conocido para la familia (Moré et al., 2005).
Entre los coleópteros, algunas especies comían polen y otras se alimentaban de tejidos florales o, incluso, del receptáculo común del capítulo. La abundancia de los escarabajos en los capítulos del girasol fue baja. Ocasionalmente, se observaron grupos de individuos copulando sobre los capítulos (p.e., Astylus atromaculatus en RO) mientras se alimentaban de granos de polen.
Las langostas y, posiblemente, también las cucarachas, se alimentaban de granos de polen que forrajeaban en las anteras o sobre los estigmas.
Cargas polínicas corporales de los lepidópteros nocturnos
Se revisaron las cargas polínicas de 23 polillas: 21 noctuidos (nueve individuos de Pseudaletia unipuncta, tres de P. adultera, dos de Peridroma saucia, dos de Rachiplusia nu, dos de Heliothis molochitina Berg y tres de Spodoptera frugiperda) y dos crámbidos (Achyra bifidalis). En las espiritrompas estudiadas, las polillas transportaban 78,34 ± 98,72 (rango = 1-320) granos de polen de girasol (Fig. 1). La mayoría de los individuos procesados transportaban hasta 100 granos de polen de girasol (χ2=22,54, P<0,000) y solamente cinco individuos transportaban cargas polínicas superiores a 200 granos de polen.
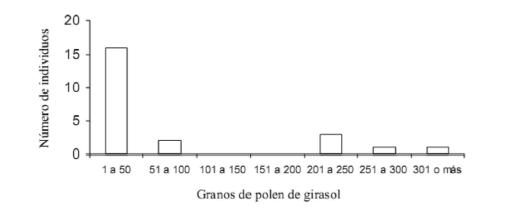
Fig. 1. Distribución de frecuencias de granos de polen de girasol en espiritrompas de lepidópteros nocturnos, capturados sobre capítulos de girasol (N=23 polillas pertenecientes a ocho especies).
Receptividad estigmática del girasol
El girasol exhibe una marcada diferencia de la receptividad estigmática a lo largo del día (χ2=133,16, P<0,000). Las mediciones de la receptividad estigmática mostraron que el estigma del girasol se encuentra receptivo durante las horas de luz del día (Fig. 2). A la mañana, la receptividad fue intermedia y alcanzó su máximo hacia el mediodía. Durante la tarde, se observó una marcada disminución y a la noche, si bien las ramas estigmáticas estaban abiertas, la superficie del estigma no se encontraba receptiva.

Fig. 2. Variación del índice de receptividad estigmática en flores emasculadas de girasol (Helianthus annuus) a lo largo del día. Se representa la media ± desvío estándar.
DISCUSIÓN
El presente trabajo es el primero en relevar sistemáticamente las especies de visitantes nocturnos del girasol en un amplio rango del área del cultivo de esta especie, en uno de los principales países productores del cultivo, la Argentina. Los resultados mostraron que, llamativamente, los capítulos de girasol son visitados por más especies de lepidópteros nocturnos que de abejas (32 especies), sus principales polinizadores (Torretta et al., enviado).
En la literatura existen muy pocos datos acerca de las visitas de lepidópteros nocturnos a las flores de girasol. Lehman et al. (1973) informaron acerca de grandes poblaciones de dos especies de noctuidos, "beet army worm" y "cabbage looper moth", que probablemente correspondan a las especies Spodoptera exigua (Hübner) y Trichoplusia ni (Hübner), respectivamente, en capítulos de girasoles en áreas desérticas del sur de California. En otro estudio, Del Socorro & Gregg (2001) informaron que Helicoverpa armigera (Hübner) (Noctuidae) se alimenta de girasol en Queensland, Australia y utilizan los granos de polen del girasol (un cultivo menor dentro del área de estudio; <1% del área cultivada) como marcador en un estudio acerca del desplazamiento local de este lepidóptero nocturno, cuyas larvas consumen dicho polen. Por otro lado, como se mencionó anteriormente, el olor de los granos del polen del girasol estimula las hembras de H. electellum a oviponer (Delisle et al., 1989). Si bien en este trabajo no se estudió específicamente la oviposición de polillas en las flores del cultivo, la falta de observaciones casuales de hembras oviponiendo sugiere que el fenómeno, si existía, no era frecuente.
En un trabajo acerca de los insectos asociados al girasol en diversas regiones del mundo, Charlet et al. (1997) citan numerosas especies de lepidópteros nocturnos cuyas larvas consumen diversas partes (capítulo, hojas, raíces, semillas, etc.) del cultivo, pero no mencionan ninguna como consumidora de néctar durante su estadio adulto.
Varias especies de polillas capturadas sobre las flores de girasol son consideradas plagas en sus estadios larvales. En Argentina, Pastrana et al. (2004) citan numerosas especies pertenecientes a las familias Arctiidae, Chouretidae, Crambidae, Noctuidae, y Pyralidae como especies fitófagas del girasol, aunque no han sido citadas en la bibliografía como consumidoras de néctar. De las 34 especies de Noctuidae registradas en este trabajo, nueve (Spodoptera frugiperda, Helicoverpa gelotopoeon (Dyar), H. zea (Boddie), Feltia subterranea, F. cf. gypaetina Guenée, Agrotis ipsilon (Hufnagel), A. malefida Guenée, Peridroma saucia y Rachiplusia nu) han sido citadas como especies fitófagas (Pastrana et al. 2004). Otra especie registrada como visitante floral y que, posiblemente, es fitófaga en girasol es Spodoptera cosmiodes (Walker), mencionada a menudo de manera incorrecta como S. latifascia (Walker) (Pogue, 2002).
El hecho que los adultos de polillas visiten flores del girasol en busca de néctar tendría dos consecuencias contrapuestas: por un lado, el consumo de esta recompensa floral podría acentuar el antagonismo de estos visitantes florales hacia el girasol, ya que además de dañar hojas y/o flores durante los estadios preimaginales, los lepidópteros nocturnos adultos consumen néctar sin contribuir a la polinización efectiva (puesto que actúan en un momento del día en el cual el estigma no se encuentra receptivo). Este resultado es congruente con los
obtenidos por Chamer et al. (2004), quienes no encontraron diferencias significativas en la tasa de formación de frutos entre capítulos de girasol expuestos exclusivamente a visitantes nocturnos (embolsados durante el día y expuestos a los visitantes durante la noche), y capítulos privados completamente de visitas de insectos (embolsados día y noche). Otro hecho que sugiere que las polillas no son polinizadores eficientes del girasol es la baja carga polínica comparada con la transportada por las abejas (Apis mellifera, 1183,93 ± 743,95 y Melissodes spp. y Melissoptila tandilensis Holmberg, 1035,88 ± 806,37 granos de polen; Torretta, 2007). Por otro lado, el consumo de néctar durante la noche podría determinar que la disponibilidad de esta recompensa en las flores del capítulo resultara inconstante. Ello obligaría a los visitantes diurnos a moverse más frecuentemente entre capítulos, lo que aumentaría la eficiencia de la polinización. La existencia de diferencias interflorales considerables en la cantidad de néctar por flor fue postulada por Frankie & Haber (1983), para explicar el movimiento de grandes abejas que polinizan árboles con floración explosiva (mass-flowering) en bosques neotropicales.
Al igual que las polillas, muchos coleópteros son consumidores de hojas, flores, receptáculos florales y raíces del girasol, y muchos son considerados plagas del cultivo, al que atacan en distintas etapas de su desarrollo (Saini, 2004). Numerosas especies de coleópteros de la familias Melyridae, Scarabaeidae y Elateridae se citan como insectos fitófagos del girasol (Di Ioirio, 2004a, b, Saini, 2004). Por otro lado, las especies de Tenebrionidae son conocidas por alimentarse de sustancias vegetales y ser polilécticos (Flores & Debandi, 2004). Otros coleópteros nocturnos capturados (especies de Carabidae) son predadores y podrían ser benéficos para el cultivo, ya que consumen insectos perjudiciales (Saini, 2004).
Como se mencionó anteriormente, la expansión de las ramas estigmáticas ocurre entre la noche y la mañana siguiente, aunque los resultados obtenidos en este trabajo, demuestran que la receptividad estigmática comienza en las primeras horas de luz. La marcha de la receptividad estigmática del girasol concuerda con el momento de mayor actividad de los principales polinizadores del girasol, las abejas. La ausencia de receptividad, en el período de actividad de las polillas, sería otro factor que demostraría la baja eficiencia de estos insectos como polinizadores del cultivo.
Los resultados acerca de la receptividad estigmática del girasol en este trabajo fueron obtenidos de plantas de una única población y mostraron un patrón marcadamente homogéneo. Sin embrago, Chamer (datos no publicados), estudiando la receptividad estigmática en condiciones similares a las de este trabajo durante tres años consecutivos, encontró que si bien al atardecer se observa una disminución en la receptividad estigmática, durante la noche ésta aumenta, aunque el patrón observado no es coincidente entre años. Por lo tanto, es necesario realizar trabajos experimentales de polinización nocturna controlada, observación de crecimientos de tubos polínicos y formación de frutos, que permitan esclarecer fehacientemente la dinámica de la receptividad estigmática de este cultivo.
Si bien en este trabajo no se estudió la marcha diaria de la secreción de néctar, las flores de girasol ofrecían néctar a los visitantes durante la noche. Este hecho, puede deberse a que la producción de néctar, durante la fase estaminada, sea mayor al consumo de este recurso por visitantes diurnos, o bien, que durante la noche también ocurra secreción de néctar.
Torretta et al. (ms. enviado) reportan que el 50% de los capítulos de girasol (observaciones directas de 10 capítulos en censos de 10 minutos de duración, durante tres momentos del días y a tres distancias del borde del cultivo, en ocho sitios de muestreo) de lotes agrícolas de Argentina no reciben visitas de insectos diurnos, lo cual podría indicar que el consumo de néctar en las flores de girasol por abejas (y otros insectos de actividad diurna) es bajo.
Si se consideran los resultados de este trabajo, los numerosos visitantes florales nocturnos del girasol podrían comportarse como ladrones de néctar y/o polen, y no contribuirían positivamente en la polinización de este importante cultivo oleaginoso. A diferencia de especies como Lonicera japonica (Caprifoliaceae) (Miyake & Yahara, 1998), Manfreda virginica (Agavaceae) (Groman & Pellmyr, 1999), Silene alba (Caryophyllaceae) (Young, 2002) e Isertia laevis (Rubiaceae) (Wolff et al., 2003), donde individuos de actividad nocturna y diurna aportan a la formación de frutos y/o semillas, en las flores de girasol, la actividad de los visitantes nocturnos parece no ser importante, al menos de manera directa.
Por lo mencionado anteriormente, aún es necesario demostrar fehacientemente (mediante estudios experimentales) si los visitantes nocturnos favorecen (o no) indirectamente la polinización, a través del redireccionamiento de la actividad de los visitantes diurnos.
En las especies vegetales arriba señaladas, las flores exhiben claras adaptaciones a la atracción de visitantes nocturnos, pero la presencia de recompensas (néctar y/o polen), durante el día, explica que también acudan a las flores los visitantes diurnos. En el girasol, parece ocurrir el fenómeno opuesto: tanto las características de las flores individuales como las de la inflorescencia parecen adaptadas principalmente a la atracción de insectos diurnos (ver Delisle et al., 1989), pero el gran tamaño del capítulo y la gran abundancia de plantas en los lotes cultivados determinarían que insectos presentes en el área cultivada en estadios preimaginales (incluso antes de que el cultivo florezca) resulten atraídos, si al emerger como adultos el cultivo se encuentra en floración. Otra explicación posible, que podría ser puesta a prueba con estudios de sitios de oviposición en distintas especies de polillas, es que el aroma emitido por el polen del girasol atraiga polillas migrantes, de manera similar a lo que ocurre con H. electellum (Delisle et al., 1989).
Futuros trabajos experimentales deberán esclarecer qué proporción del rendimiento es atribuible a la actividad de los visitantes nocturnos (directa o indirectamente), en este importante cultivo oleaginoso.
AGRADECIMIENTOS
A los propietarios de lotes de girasol que nos permitieron desinteresadamente trabajar en sus campos. A Gabriela Cilla, Norberto Montaldo y Mariano Devoto por la ayuda en el campo. A Francisco Crespo y Alejandra Valverde (Blattaria), Susana Aranda (Elateridae), Analía Lanteri (Curculionidae), Armando Cicchino (Carabidae), Gustavo Flores (Tenebrionidae) y Osvaldo Di Dorio (otros Coleoptera) que colaboraron en la determinación de diversos insectos. A Karina Hodara por la ayuda con el análisis estadístico de los datos. A la ANPCyT y a la Universidad de Buenos Aires por el apoyo económico (subsidios PICT 08-12504 y G065, respectivamente). Este estudio formó parte de la tesis doctoral de J.P.T.. A Rubén Alarcón, Diego Vázquez y a un revisor anónimo por las sugerencias que permitieron mejorar la calidad de este trabajo. J.P.T., F.N. y D.M. son investigadores del CONICET, Argentina, entidad a la que se agradece el continuo apoyo.
BIBLIOGRAFÍA CITADA
1. CHAMER, A. M., M. DEVOTO, J. P. TORRETTA, D. MEDAN, A. I. MANTESE, N. H. MONTALDO & N. BARTOLONI. 2004. Polinización del girasol: efecto sobre el rendimiento. II Reunión Binacional de Ecología: 395. [ Links ]
2. CHARLET, L. D., G. J. BREWER & B. A. FRANZMANN. 1997. Sunflower insects. En: Schneiter A. (ed.), Sunflower technology and production. American Society of Agronomy, Madison, pp.183-261. [ Links ]
3. DAFNI, A. & M. MOTTÉ-MAUÉS. 1998. A rapid and simple procedure to determine stigma receptivity. Sex Plant Reprod. 11: 177-180. [ Links ]
4. DARRAULT, R. O. & C. SCHLINDWEIN. 2005. Limited fruit production in Hancornia speciosa (Apocynaceae) and pollination by nocturnal and diurnal insects. Biotropica 37: 381-388. [ Links ]
5. DELAPLANE, K. S. & D. F. MAYER. 2000. Crop pollination of bees. CAB International, Cambridge. [ Links ]
6. DELISLE, J., J. N. MCNEIL, E. W. UNDERHILL & D. BARTON. 1989. Helianthus annuus pollen, an oviposition stimulant for the sunflower moth, Homoeosoma electellum. Entomol. Exp. Appl. 50: 53-60. [ Links ]
7. DEL SOCORRO, A. P. & P. C. GREGG. 2001. Sunflower (Helianthus annuus L.) pollen as a marker for studies of local movement in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Aust. J. Entomol. 40: 257-263. [ Links ]
8. DI IORIO, O. R. 2004a. Melyridae. En: Cordo, H. A., G. Logarzo, K. Braun & O. R. Di Iorio (eds.), Catálogo de los insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina ediciones, Buenos Aires, pp. 175-179. [ Links ]
9. DI IORIO, O. R. 2004b. Scarabaeidae. En: Cordo, H. A., G. Logarzo, K. Braun & O. R. Di Iorio (eds.), Catálogo de los insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina ediciones, Buenos Aires, pp.183-196. [ Links ]
10. FELL, R. D. 1986. Foraging behaviors of Apis mellifera L. and Bombus spp. on oilsedd sunflower (Helianthus annuus L.). J. Kans. Entomol. Soc. 59: 72-81. [ Links ]
11. FLORES, G. E. & G. O. DEBANDI. 2004. Tenebrionidae. En: Cordo, H. A., G. Logarzo, K. Braun & O. R. Di Iorio (eds.), Catálogo de los insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina ediciones, Buenos Aires, pp. 197-201. [ Links ]
12. FRANKIE, G. W. & W. A. HABER. 1983. Why bees move among mass-flowering neotropical trees. En: Jones, C. E. & R. J. Little (eds.), Handbook of experimental pollination biology. Van Nostrand Reinhold, New York, pp. 360-372. [ Links ]
13. FREE, J. B. 1993. Insect pollination of crops. Academic Press, San Diego. [ Links ]
14. GROMAN, J. D. & O. PELLMYR. 1999. The pollination biology of Manfreda virginica (Agavaceae): relative contribution of diurnal and nocturnal visitors. Oikos 87: 373-381. [ Links ]
15. HURD, P. D., W. E. LABERGE & E. G. LINSLEY. 1980. Principal sunflower bees of North American with emphasis on the southwestern United States (Hymenoptera: Apoidea). Smithson. Contrib. Zool. 310: 1-158. [ Links ]
16. KRENN, H. W., J. D. PLANT, N. U. SZUCSICH. 2005. Mouthparts of flower-visiting insects. Arthropod. Struct. Dev. 34: 1-40. [ Links ]
17. LEHMAN, W. F., F. E. ROBINSON, P. F. KNOWLES & R. A. FLOCK. 1973. Sunflowers in the desert valley areas of Southern California. Calif. Agric. 27: 12-14 [ Links ]
18. MCGREGOR, S. E. 1976. Insect pollination of cultivated plants. Agriculture Handbook Nº 496. Agricultural Research Service, Washington. [ Links ]
19. MIYAKE, T. & T. YAHARA. 1998. Why does the flower of Lonicera japonica open at dusk? Can. J. Bot. 76: 1806-1811. [ Links ]
20. MORÉ, M., I. J. KITCHING & A. A. COCUCCI. 2005. Sphingidae: Esfíngidos de Argentina. Editorial L.O.L.A., Buenos Aires. [ Links ]
21. NEFF, J. L. & B. B. SIMPSON. 1990. The roles of phenology and reward structure in the pollination biology of wild sunflower (Helianthus annuus L., Asteraceae). Israel J. Bot. 39: 197-216. [ Links ]
22. PASTRANA, J. A., O. R. DI IORIO, F. NAVARRO, A. CHALUP & M. E. VILLAGRÁN. 2004. Lepidoptera. En: Cordo, H. A., G. Logarzo, K. Braun & O. R. Di Iorio (eds.), Catálogo de los insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina ediciones, Buenos Aires, pp. 416-515. [ Links ]
23. POGUE, M. G. 2002. A world revision of the genus Spodoptera Guenée (Lepidoptera Noctuidae). Mem. Am. Entomol. Soc. 43: 1-202. [ Links ]
24. SAINI, E. D. 2004. Insectos y ácaros perjudiciales al cultivo del girasol y sus enemigos naturales. Publicación del Instituto de Microbiología y Zoología Agrícola Nº 8. INTA. Buenos Aires. [ Links ]
25. SCHNEITER, A. A. & J. F. MILLER. 1981. Description of sunflower growth stages. Crop Sci. 21: 901-903. [ Links ]
26. SCOTT, J. A. 1986. The butterflies of North America. A natural history and field guide. Stanford University Press. California. [ Links ]
27. SEILER, G. J. 1997. Anatomy and morphology of sunflower. En: Schneiter, A. (ed.), Sunflower technology and production. American Society of Agronomy, Madison, pp. 67-111. [ Links ]
28. STEPHENSON, A. G. & W. W. THOMAS. 1977. Diurnal and nocturnal pollination of Catalpa speciosa (Bignoniaceae). Syst. Bot. 2: 191-198. [ Links ]
29. TORRETTA, J. P., M. DEVOTO, A, M. CHAMER, D. MEDAN, N. H. MONTALDO & F. NAVARRO. 2005. Visitantes florales nocturnos del girasol (Helianthus annuus L.). Bol. Soc. Arg. Bot. 40 (supl): 39. [ Links ]
30. TORRETTA, J. P. Inéd. Entomofauna relacionada a la polinización del girasol (Helianthus annuus L.) en Argentina. Tesis Doctoral, Universidad de Buenos Aires, Buenos Aires, 2007, 91 pp. [ Links ]
31. TORRETTA, J. P., D. MEDAN, A. ROIG ALSINA & N. H. MONTALDO. Visitantes florales diurnos del girasol (Helianthus annuus, Asterales: Asteraceae) en Argentina. Enviado a Revista de la Sociedad Entomológica Argentina. [ Links ]
32. WOLFF, D., M. BRAUN & S. LIEDE. 2003. Nocturnal versus diurnal pollination success in Isertia laevis (Rubiaceae): a sphingophilous plant visited by hummingbirds. Plant biol. 5: 71-78. [ Links ]
33. YOUNG, H. J. 2002. Diurnal and nocturnal pollination of Silene alba (Caryophyllaceae). Am. J. Bot. 89: 433-440. [ Links ]
34. ZAR, 1996. Biostatistical analysis. Prentice Hall. New Jersey. [ Links ]

