










Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.70 no.3-4 Mendoza jul./dic. 2011
TRABAJOS CIENTÍFICOS
Fenologia de Cornops aquaticum (Orthoptera: Acrididae) associado a Eichhornia crassipes (Pontederiaceae) em um Lago de Várzea na Amazônia Central, Brasil
Phenology of Cornops aquaticum (Orthoptera: Acrididae) associated with Eichhornia crassipes (Pontederiaceae) in a Floodplain Lake in Central Amazonia, Brazil
Braga, Carlos Elias *, Ana Lúcia Nunes-Gutjahr *, José Wellington de Morais ** & Joachim Adis† ***
* Universidade do Estado do Pará, Centro de Ciências Sociais e Educação, Departamento de Ciências Naturais, Rua do Una nº 156, Telégrafo, Belém, PA, Brasil. CEP: 66113-200; e-mail: bragaelias@yahoo.com.br.
** Instituto Nacional de Pesquisas da Amazônia, Coordenação de Pesquisa em Entomologia, Avenida André Araújo nº 2936, Petrópolis, Manaus, AM, Brasil. CEP: 60083-000.
*** Max-Planck Institut Für Limnologie, Plön, Alemanha (in memoriam).
Recibido: 27-III-2011
Aceptado: 1-VII-2011
RESUMO. O gafanhoto Neotropical Cornops aquaticum (Bruner) vive associado às macrófitas da família Pontederiaceae da qual se alimenta. Nos lagos da Amazônia Central, Eichhornia crassipes (Mart.) Solms (mururé ou aguapé) é considerada a planta hospedeira de maior importante para este gafanhoto. Esse trabalho teve o objetivo de realizar um estudo fenológico de adultos e ninfas desse gafanhoto em associação a sua planta hospedeira, frente ao regime hidrológico da Amazônia Central. Tal estudo foi realizado entre os meses de abril de 2006 a agosto de 2007 no Lago Camaleão (03º17'05"S 60º11'11"W), na várzea da Amazônia Central. Para isso, utilizou-se uma rede entomológica (70 cm de diâmetro) adaptada para a captura dos gafanhotos. Onde foram capturados 850 exemplares (296 adultos e 554 ninfas). Observou-se que a abundância e a biomassa de adultos e de ninfas de C. aquaticum, bem como de sua macrófita hospedeira, estão relacionados com a oscilação sazonal do nível do rio (pulso de inundação).
PALAVRAS CHAVE. Gafanhoto semi-aquático; Estudo fenológico; Região Amazônica; Aguapé.
ABSTRACT. The Neotropical grasshopper Cornops aquaticum (Bruner) lives associated with the macrophytes of the family Pontederiaceae from which it feeds. In the lakes of Central Amazon, Eichhornia crassipes (Mart.) Solms (water hyacinth or Mururé) is considered the most important host plant for this grasshopper. This study aimed to perform a phenological study of adults and nymphs of grasshopper in association with its host plant, compared to the hydrological regime of the Central Amazon. This study was undertaken between April 2006 and August 2007 at the Chameleon Lake (03 17'05"S, 60 11'11"W) in the Central Amazon floodplain. An entomological net adapted for catching grasshoppers was used. 850 individuals (296 adults and 554 nymphs) were captured. It was observed that the abundance and biomass of adults and nymphs of C. aquaticum, as well as their macrophyte host are related to the "flood pulse", ie. to the seasonal oscillations of the Amazonian rivers.
KEY WORDS. Semi-aquatic grasshopper; Phenological study; Amazon Region; Water hyacinth.
RESUMEN. La tucura Neotropical, Cornops aquaticum (Bruner), vive asociada a las macrófitas de la familia Pontederiaceae, de las cuales se alimenta. En los lagos de la Amazonia Central, Eichhornia crassipes (Mart.) Solms (camalote o aguape) constituye la planta huésped más importante de esta tucura. El objetivo de este trabajo fue estudiar la fenología de los adultos y las ninfas de C. aquaticum en los camalotales de E. crassipes, en relación al régimen hidrológico de la Amazonia Central. Los muestreos se realizaron entre los meses de abril de 2006 a agosto de 2007, en el Lago Camaleón (03º17'05"S 60º 11 '11 "O) en la Várzea de la Amazonia Central. Los individuos fueron capturados desde una embarcación a motor, utilizando una red entomológica de 70 cm de diámetro. Durante este estudio, se capturaron un total de 850 ejemplares (296 adultos y 554 ninfas). Se observó que la abundancia y la biomasa de los adultos y de las ninfas de C. aquaticum, así como la planta huésped, están estrechamente relacionados con la oscilación estacional del nivel del río (pulso de inundación).
PALABRAS CLAVE. Tucuras semiacuáticas; Estudio fenológico; Amazonia; Camalote.
INTRODUÇÃO
As planícies inundáveis da Região Amazônica cobrem uma área com cerca de 300.000 Km (Junk & & Piedade, 1993) e são definidas como áreas periodicamente inundadas (Junk et al. 1989). Na Amazônia Central, o nível dos rios e lagos sofre anualmente uma oscilação vertical média de 10 m (Schmidt, 1973). Essa variação hídrica expõe extensas áreas de floresta a períodos de inundações de cinco a seis meses (Adis, 1985) e influencia de forma diferenciada a biota da região, impondo a fauna e a flora a uma dinâmica de sobrevivência entre uma fase terrestre e outra aquática, tornando necessário que as mesmas desenvolvam adaptações para sobreviver nesses ambientes instáveis (Junk et al., 1989; Adis & Junk, 2002). Nessa região, podem-se encontrar áreas de várzea sazonal e igapó sazonal que são inundadas periodicamente por água branca e água preta, respectivamente (Prance, 1979).
Cornops aquaticum (Bruner) é um gafanhoto semi-aquático, Neotropical, que pode ser encontrado desde o sudeste do México até o centro da Argentina e Uruguai (Adis et al., 2007). Essa espécie possui porte médio de 2.5 a 3.0 cm de comprimento, com coloração verde escura e faixa pós-ocular preta e amarela que se estende nas laterais do corpo (Lhano et al., 2005). C. aquaticum desenvolve seu ciclo de vida associado às populações de macrófitas aquáticas pertencentes à família Pontederiaceae. Todavia, mostra grande especificidade alimentar e hospedeira, principalmente, aos gêneros Eichhornia Kunth e Pontederia L (Ferreira & Vasconcelos-Neto, 2001). Este trabalho objetivou realizar um estudo sobre a fenologia de adultos e ninfas de C. aquaticum em associação a E. crassipes, em um lago de várzea, frente ao regime hidrológico da Amazônia Central.
MATERIAL E MÉTODOS
Área de Estudo
O estudo foi realizado no Lago Camaleão (03º17'05"S, 60º11'11"W), na Ilha de Marchantaria, distante cerca de 20 km da cidade de Manaus (Irion et al., 1983; Waichman et al., 2002), no estado do Amazonas (Fig. 1). O Lago se encontra conectado por uma enseada do Rio Solimões e tem a forma alongada com 7 km de comprimento, 300 a 500 m de largura e 6 m de profundidade durante o nível médio das águas (Soares et al., 1986).
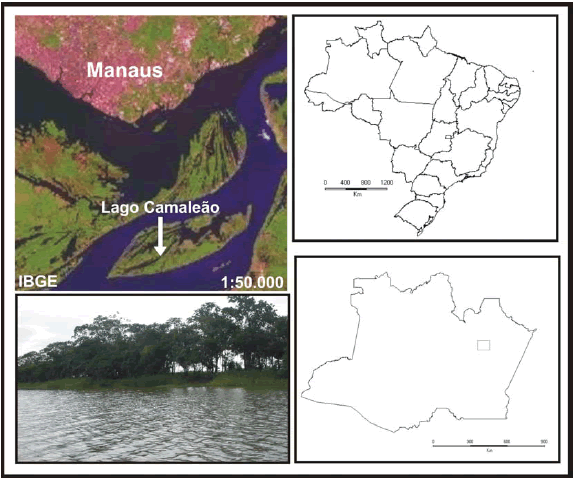
Fig. 1. Localização do Lago Camaleão (esquerda superior) e fotografia mostrando o aspecto do Lago (esquerda inferior).
Metodologia
O estudo da fenologia de ninfas e adultos de Cornops aquaticum foi realizado através de coletas mensais de abril/2006 a agosto/2007, totalizando 17 meses de amostragem, nas colônias de Eichhornia crassipes no Lago Camaleão.
A metodologia utilizada foi baseada em Franceschini et al. (2007), sendo a coleta do tipo varredura sobre as colônias de E. crassipes efetuada por uma pessoa com auxílio de uma rede entomológica, adaptada para esse tipo de coleta (aro com 70 cm de diâmetro e saco coletor de 100 cm de comprimento), para o deslocamento na área estudada foi utilizado um barco a motor. O esforço de amostra foi baseado na coleta de 50 exemplares de C. aquaticum (entre adultos e ninfas). Os gafanhotos capturados foram acondicionados em sacos plásticos e transportados vivos em caixa de poliestileno (isopor).
No laboratório os exemplares foram introduzidos em freezer para serem mortos, por resfriamento, após esse procedimento se fez à triagem e quantificação dos indivíduos por sexo, no caso dos adultos (machos e fêmeas) e por ninfas de categorias, de acordo com o comprimento do fêmur posterior, denominadas, tipo A (< 5 mm) e B (> 5 mm) (Franceschini et al. 2005). Em seguida, os exemplares triados foram pesados em balança analítica (Sartorius 0.0001 g) por grupos de categorias (machos, fêmeas, ninfas A e ninfas B) para determinação da biomassa (peso fresco/g). Todo o material coletado foi acondicionado e preservado em vidros (20 mL) etiquetados contendo álcool a 80%.
Os dados climatológicos (temperatura, umidade relativa, insolação e precipitação) apresentados foram gerados e fornecidos pelo Instituto Nacional de Meteorologia (INMET/1º DISME/AM/AC/RR), Seção de Observação Meteorológica (SEOMA). Os dados relativos ao nível da água do Rio Negro foram obtidos a partir das medições disponibilizadas, diariamente, pela Superintendência Estadual de Navegação, Portos e Hidrovias (SNPH) em Manaus, Governo do Estado do Amazonas, no endereço eletrônico www. snph.am.gov.br. Quanto a isso, Schmidt (1973) afirma que os dados do nível do Rio Negro podem ser utilizados para caracterizar as flutuações do nível d'água em áreas de até 100 km rio acima, com diferenças de poucos centímetros.
Dados Abióticos
Durante o período de realização do estudo, foram observados os seguintes dados climáticos: temperaturas mínima de 23.1ºC, máxima de 35.4ºC e média de 28.7ºC; umidade relativa média variando 74% a 91%; precipitação total mensal mínima de 16.7 mm, máxima de 414.4 mm e média de 196.3 mm; Insolação total mensal mínima de 117 h, máxima de 259 h e média de 174.8 h.
Análise de Dados
A análise estatística foi realizada através dos testes de Regressão Linear e Teste t, com intervalo de confiança de α = 0.05 e, também se utilizou o coeficiente de Correlação Linear de Pearson, (r), onde r = 0 (ausência de relação), r = 1 (relação positiva perfeita), r = -1 (relação negativa perfeita), 0 < r < 0.30 (relação positiva fraca), 0.30 < r < 0.60 (relação positiva moderada), 0.60 < r < 0.90 (relação positiva forte), 0.90 < r < 1 (relação positiva muito forte), 0 < r < -0.30 (negativa fraca), -0.30 < r < -0.60 (negativa moderada), -0.60 < r < -0.90 (negativa forte) e -0.90 < r < -1 (negativa muito forte) (Fonseca et al. 1991). Todos os dados foram analisados com o auxílio dos programas estatísticos R 2.5.1 e SYSTAT 11.
RESULTADOS
Durante o período de desenvolvimento do estudo, o nível da água do Rio Negro, durante os dias de coletas em campo, oscilou de 28.79 m (cota máxima) a 17.10 m (cota mínima), ocasionando uma diferença entre a cota mínima e máxima de 11.69 m (Tabela I).
Tabela I. Abundância de adultos (machos e fêmeas) e ninfas (A e B) de Cornops aquaticum, coletados no Lago Camaleão durante o período de abril/2006 - agosto/2007 e o nível Rio Negro (m).

Durante o período de amostragem, foram coletados 850 exemplares de C. aquaticum entre adultos (n = 296) e ninfas (n = 554) (Tabela I). A abundância de ninfas e adultos variou em relação à oscilação no nível da água dos rios Negro e Solimões e conseqüentemente no lago estudado (regressão linear: p < 0.001). Durante o período de cheia foi observada a maior abundância de ninfas (n = 364) e na fase de seca os adultos foram mais abundantes (n = 160) (Fig. 2).

Fig. 2. Correlação da abundância para adultos e para ninfas de Cornops aquaticum com o nível da água do Rio Negro (m).
A correlação estabelecida, de forma geral, entre a abundância de indivíduos de C. aquaticum e a variação do nível da água do Rio Negro, foi considerada negativa para adultos (r = -0.61) e positiva para ninfas (r = 0.61), indicando que com a diminuição do nível da água (vazante: período de seca), houve uma tendência de aumento do número de exemplares adultos e diminuição da quantidade de ninfas. De modo contrário quando houve a elevação do nível da água (enchente: período de cheia), foi observado a diminuição de indivíduos adultos e o aumento do número de ninfas (Fig. 2). Vale ressaltar que outros fatores abióticos como, temperatura, umidade relativa, insolação e precipitação também foram relacionadas com os resultados obtidos, mas nenhum deles se mostrou significativo em relação à variação da população de C. aquaticum.
Para os imaturos a maior abundância mensal ocorreu em maio/2006 e junho/2007, quando foram registrados em ambos os meses 47 ninfas, correspondendo a 94% dos exemplares coletados no mês, conseqüentemente foi observado o menor número de adultos (correspondente a 6%). De forma contrária, no mês de janeiro/2007 os resultados mostraram a maior abundância de adultos com 40 indivíduos (80% dos exemplares no mês) e menor de ninfas (correspondente a 20%) (Tabela I).
Os resultados mostram que o total de machos e fêmeas durante o período de amostragem apresentou valores muito próximos (machos: n = 151 e fêmeas n = 145). Observou-se que em 14 dos 17 meses de coleta a diferença entre o número de machos e fêmeas não passou de três indivíduos, sendo que na maioria dos casos o número de machos ficou levemente acima da quantidade de fêmeas (Teste t: p > 0.05).
No mês de novembro de 2006, registrou-se, a maior diferença, quando foram coletadas 20 fêmeas e 12 machos. Vale ressaltar que no mês de maio/2006, devido ao baixo número de adultos (três exemplares), não foram capturadas fêmeas adultas. (Fig. 3 e Tabela I).

Fig. 3. População de e fêmeas de Cornops aquaticum, frente à oscilação do nível da água do Rio Negro durante o período de abril/2006 a agosto/2007.
A quantidade de Ninfas A (n = 374) em quase todos os meses de coleta foi superior a de Ninfas B (n = 180), exceto em janeiro/2007. A análise através do Teste t indicou haver diferenças significativas entre o número ninfas A e B (P < 0.001) (Fig. 4 e Tabela I).
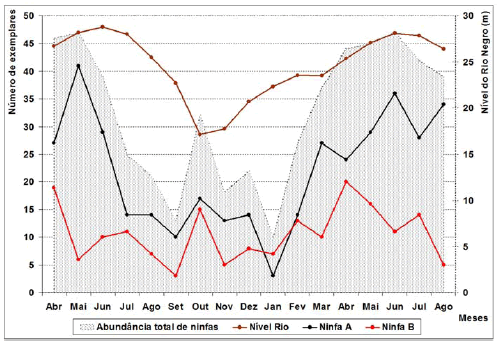
Fig. 4. População Ninfas A e B de Cornops aquaticum, frente à oscilação do nível da água do Rio Negro durante o período de abril/2006 a agosto/2007.
Os dados referentes à biomassa (peso fresco em gramas) de adultos e ninfas de C. aquaticum observados, também variou com a oscilação do nível do rio, quando se registraram os maiores índices de biomassa total (adultos + ninfas) no período em que o nível da água estava mais baixo (vazante/ seca) e os menores valores de biomassa quando o nível da água estava mais alto (enchente/cheia) (Fig. 5 e Tabela II).

Fig. 5. Biomassa (g) de adultos e ninfas de Cornops aquaticum e nível do Rio Negro, no Lago Camaleão durante o período de abril/2006 a agosto/2007.
Tabela II. Biomassa (g) de adultos (macho e fêmea) e ninfas (A e B) de Cornops aquaticum, coletados no Lago Camaleão durante o período de abril/2006 - agosto/2007.
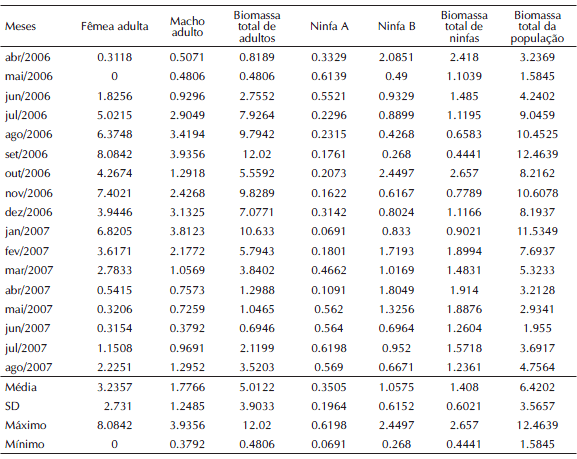
A biomassa média de um gafanhoto C. aquaticum (biomassa total/ abundância total) por estágio e sexo, foi a seguinte: adultos machos = 0.2000 g; adultos fêmeas = 0.3794 g; ninfa A = 0.0159 g; ninfa B = 0.0999 g. Os valores máximos de biomassa registrados aconteceram nos meses de setembro/2006 (biomassa total: n = 12.4639 g e de adultos: n = 12.0198 g) e outubro/2006 no caso das ninfas (n = 2.6570 g). Com relação aos valores mínimos de biomassa, esses foram observados em maio/2006 para biomassa total (n = 1.5845 g) e de adultos e em setembro/2006 (n = 0.4806 g), para ninfas (n = 0.4441 g) (Fig. 5 e Tabela II).
Os índices de biomassa, assim como ocorreu com a abundância, registraram maiores valores para adultos quando o nível da água baixou e vice-versa, exceto para as ninfas onde essa variação não foi tão evidenciada, apesar de haver uma tendência de aumento e diminuição de biomassa conforme a elevação e redução do nível da água (Fig. 5 e Tabela II). Quanto a isso, o teste de Regressão Linear, mostrou valores significativos para adultos (p < 0.05) e não significativos (p> 0.05) para as ninfas.
Os resultados mostraram que, em relação às ninfas A, as variações da biomassa não foram tão extremas entre os índices mais altos e baixos, quando em comparação às observadas para adultos (machos e fêmeas) e ninfas B, durante o período de amostragem (Figs 6 e 7).

Fig. 6. Biomassa (g) de machos e fêmeas de Cornops aquaticum, frente à oscilação do nível da água do Rio durante o período de abril/2006 a agosto/2007.

Fig. 7. Biomassa (g) de Ninfas A e B de Cornops aquaticum, frente a oscilação do nível da água do Rio Negro durante o período de abril/2006 a agosto/2007.
DISCUSSÃO
Estudos realizados na Amazônia Central demonstram que a flutuação do nível das águas de seus rios, lagos e demais corpos d'água (pulso de inundação), exerce influência direta ou indireta na fauna e flora dessa região (Junk, 1980; Nunes, 1989). Essa dinâmica se manifesta através de estratégias adaptativas necessárias para a sobrevivência desses animais e plantas aos períodos de seca (fase terrestre) e cheia (fase aquática) (Junk, 1980; Adis, 1997). O pulso de inundação exerce ação direta sobre a macrófita aquática Eichhornia crassipes que desenvolve formas adaptadas ao período de seca e cheia dos rios. Essas modificações morfo-fisiológicas permitem que essa planta sobreviva às oscilações hidrológicas sazonais nessa região (Junk & Piedade, 1993). Todavia, a influência desse fenômeno é considerada indireta sobre Cornops aquaticum, visto que, esse gafanhoto desenvolve todo seu ciclo de vida associado às populações de macrófitas aquáticas dos gêneros Eichhornia e Pontederia (Ferreira & Vasconcelos-Neto, 2001), portanto sua dinâmica populacional depende da maior ou menor abundância (disponibilidade) dessas macrófitas aquáticas nesses ambientes (Nunes, 1989).
Os resultados obtidos neste trabalho mostraram que a variação hídrica (pulso de inundação) exerce forte influência sobre a população de C. aquaticum do Lago Camaleão, na várzea da Amazônia Central. Contudo, notou-se certa tendência na população de ninfas e adultos de C. aquaticum variar conforme a oscilação no nível da água dos rios Solimões e Negro e, conseqüentemente, desse lago. Essa constatação foi decorrente da maior abundância de ninfas durante a época de cheia e enchente e no período de seca e vazante a maior ocorrência de adultos e vice-versa, cuja correlação estabelecida, em ambos os lagos, foi positiva para ninfas e negativa para adultos e também considerada como forte (Fonseca et al., 1991).
Pelo exposto anteriormente, acreditase que há uma expressiva ligação de C. aquaticum com sua macrófita hospedeira frente à variação hídrica sazonal existente nesse lago, cuja variação das populações do gafanhoto, está relacionada à dinâmica populacional de E. crassipes nesses ambientes. Tal relação foi demonstrada em estudo de Silva et al. (2010) realizado no Pantanal do Mato Grosso entre 2006 e 2007, que apresentou resultados similares aos encontrados neste trabalho. Braga (2008) estudou no mesmo período as populações de C. aquaticum em associação com E. crassipes no Lago Janauari (água mista: várzea & igapó), também na Amazônia Central, e obteve resultados relativamente próximos aos encontrados neste estudo, corroborando com as observações verificadas neste trabalho.
Franceschini et al. (2007), determinou que a variação populacional de C. aquaticum, no Noroeste da Argentina, estava diretamente relacionada à maior ou menor quantidade de sua macrófita hospedeira (Eichhornia azurea L. e E. crassipes), no entanto, tais relações estavam estreitamente influenciadas pelas estações do ano naquela região, onde as máximas populacionais das plantas e dos gafanhotos ocorriam no verão e as mínimas no inverno.
A maior abundância de ninfas de C. aquaticum na época de cheia (enchente) no lago estudado, é devida provavelmente a melhor disponibilidade alimentar, pois nesse período existem grandes colônias de E. crassipes. Quanto a isso, Junk (1980) afirma que as macrófitas aquáticas no período de cheia e enchente, na Amazônia Central, expressam suas máximas populacionais em decorrência da boa condição ambiental. Durante esse período a competição por alimento entre os imaturos (ninfas) e os adultos de C. aquaticum é minimizada, ocasionando a diminuição da mortalidade das ninfas. Outro fato a considerar, refere-se as oviposições endofíticas de C. aquaticum, pois estando a planta hospedeira em abundância e em boa condição morfológica, há maior disponibilidade de plantas aptas para as oviposições das fêmeas, contribuindo para o aumento na taxa de natalidade desses gafanhotos.
O fato de se encontrar ninfas A (pequenas) durante todo o período de amostragem, demonstra que a reprodução desses gafanhotos é contínua durante o ano todo, com visível, tendência, de picos reprodutivos durante a época de cheia e enchente, quando suas macrófitas hospedeiras também estão em máximas populacionais.
Por outro lado, a maior quantidade de adultos na época seca e vazante, pode ser considerada uma estratégia de sobrevivência da espécie, visto que, no estágio adulto C. aquaticum pode dispersar por vôo em busca de áreas com maior disponibilidade de sua planta hospedeira. Quanto a isso Nunes et al. (1992), fez a mesma observação para Stenacris fissicauda fissicauda Bruner que é outra espécie de gafanhoto semi-aquático, da mesma área de estudo.
Entretanto, vale destacar que durante o período de amostragem, verificou-se uma tendência de haver uma maior abundância de machos em relação à quantidade de fêmeas. Tal acontecimento pode estar relacionado à duração do período ninfal, visto que, as fêmeas tendem a apresentar um estádio ninfal extra a mais, que os machos (Zolessi, 1956; Adis & Junk, 2003). Dessa forma, apresenta um tempo maior para o desenvolvimento ninfal, permitindo que sempre um número suficiente de machos sexualmente maturos estejam presentes para copular tão logo ocorra à emersão de fêmeas (Nunes et al., 1992; Braga, 2008).
Quanto à biomassa de C. aquaticum, estudos de Franceschini et al. (2007) e Adis & Junk (2003) indicam que sua variação está influenciada pelos estágios de desenvolvimento. Dessa forma, os picos máximos de biomassa são decorrentes da predominância de adultos, que são maiores e conseqüentemente mais pesados, e os mínimos dependem do maior número de ninfas. Por essa razão, observou-se que os maiores valores de biomassa total (adultos + ninfas) tenderam a ocorrer durante o período de seca ou vazante no Lago Camaleão, em virtude de haver nessa época a maior abundância ou predominância de indivíduos adultos em relação a menor quantidade de ninfas. De modo contrário, durante a cheia (enchente) quando há grande abundância de ninfas se observa os menores índices de biomassa geral. Portanto, os valores de biomassa seguem a mesma tendência de abundância de adultos ou de ninfas.
Entretanto, verificou-se que a biomassa das fêmeas adultas, no Lago Camaleão, apresentou valores superiores aos dos machos adultos na maioria das amostras. Essa tendência pode ter ocorrido devido ao fato das fêmeas serem em média 1,88 vezes mais pesadas que os machos, o que é devido ao tamanho das fêmeas que são sempre maiores que os machos. Quanto às ninfas foi observado que a biomassa das Ninfas B se manteve mais elevada do que a verificada para ninfas A durante a maioria dos meses, o que foi decorrente do tamanho e consequentemente a grande massa corporal dessas ninfas. Dessa forma, nem mesmo a numerosa quantidade de ninfas A, observada no período de cheia e enchente, pôde influenciar no resultado da biomassa geral.
CONCLUSÃO
Conclui-se que o pulso de inundação constitui o principal fator que influência a dinâmica populacional de C. aquaticum. Ele atua de forma indireta na população dos gafanhotos e direta na sua macrófita hospedeira E. crassipes. A influência do pulso e inundação sobre os gafanhotos C. aquaticum e sua respectiva macrófita hospedeira pode ser verificada através da variação da abundância e biomassa tanto das plantas quantos dos gafanhotos estudados no Lago Camaleão.
AGRADECIMENTOS
Os autores agradecem ao Instituto Nacional de Pesquisas da Amazônia (INPA) e ao Programa Insect Coevolution on Waterhyacinth (HICWA), através do Projeto INPA/Max-Planck pelo apoio financeiro, logístico e laboratorial que proporcionou a realização deste trabalho. Ao Instituto Nacional de Meteorologia (INMET/1° DISME/ AM/AC/RR) pelo fornecimento dos dados meteorológicos.
BIBLIOGRAFIA CITADA
1. ADIS, J. 1985. Adaptations of arthropods to Amazonian inundation-forests. - In: Misra, K.C. et al. (eds). Ecology and resource management in tropics. Vol.1. Silver Jubilee Symposium, Int. Soc. Trop. Ecology, Bhargava Book Depot Chowk, Varanasi, Índia. p. 29-40. [ Links ]
2. ADIS, J. 1997. Estratégias de sobrevivência de invertebrados terrestres em florestas inundáveis da Amazônia Central: uma resposta à inundação de longo período. Acta Amazonica 27(1): 43-54. [ Links ]
3. ADIS, J. & W. J. JUNK. 2002. Terrestrial invertebrates inhabiting lowland river floodplains of Central Amazonia and Central Europe: A Review. Freshwater Biology 47: 711-731. [ Links ]
4. ADIS, J. & W. J. JUNK. 2003. Feeding impact and bionomics of the grasshopper Cornops aquaticum on the water hyacinth Eichhornia crassipes in Central Amazonian Floodplains. Studies on Neotropical Fauna and Environment 38(3): 245-249. [ Links ]
5. ADIS, J., E. BUSTORF, M. G. LHANO, C. AMÉDÈGNATO & A. L. NUNES. 2007. Distribution of Cornops grasshoppers (Leptysminae: Acrididae: Orthoptera) in Latin America and the Caribbean Islands. Studies on Neotropical Fauna and Environment, 42(1): 11-24. doi: 10.1080/01650520600931719 [ Links ]
6. BRAGA, C. E. 2008. Aspectos fenológicos e consumo alimentar de Cornops aquaticum (Bruner, 1906) (Orthoptera: Acrididae: Leptysminae) associado a Eichhornia crassipes (Mart.) Solms (Pontederiaceae) em dois lagos da Amazônia Central, Brasil. Dissertação de Mestrado. Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do Amazonas. Manaus, AM, Brasil. 110 pp. [ Links ]
7. FERREIRA, S. A. & J. VASCONCELOS-NETO. 2001. Host plants of the grasshopper Cornops aquaticum (Bruner) (Orthoptera: Acrididae) in the wetland of Poconé, MT, Brazil. Neotropical Entomology 30: 523-533. [ Links ]
8. FONSECA, J. S., G. A. MARTINS, G. L. TOLEDO. 1991. Estatística aplicada. 2ª ed. Editora Atlas, São Paulo-SP. [ Links ]
9. FRANCESCHINI, M. C., J. ADIS, A. POI DE NEIFF & M. L. WYSIECKI. 2007. Fenología de Cornops aquaticum (Orthoptera: Acrididae) en un camalotal de Eichhornia azurea (Pontederiaceae) en Argentina. Amazoniana 19 (3/4): 149-158. [ Links ]
10. FRANCESCHINI, M. C., S. CAPELLO, M. LHANO, J. ADIS & M. L. WYSIECKI. 2005. Morfometría de los estadios ninfales de poblaciones de Cornops aquaticum Bruner (Acrididae: Leptysminae) en Argentina. Amazoniana 18 (3/4): 373-386. [ Links ]
11. IRION, G., J. ADIS, W. J. JUNK & F. WUNDERLICH. 1983. Sedimentological studies of the "Ilha de Marchantaria" in the Solimões/Amazon River near Manaus. Amazoniana 8 (1): 1-18. [ Links ]
12. JUNK, W. J. 1980. Áreas inundáveis - Um desafio para limnologia. Acta Amazonica 10(4): 775-795. [ Links ]
13. JUNK, W. J., P. B. BAYLEY & R. E. SPARKS. 1989. The floodpulse concept in river-floodplains systems. In: Dodge, D. P. (Ed). Proceedings of the International Large River Syposium. Canadian Special Publication of Fisheries and Aquatic Sciences 106: 110-127. [ Links ]
14. JUNK, W. J. & M. T. F. PIEDADE. 1993. Herbaceous plants of the Amazon floodplain near Manaus: Species diversity and adaptations to the flood pulse. Amazoniana 12 (3/4): 467-484. [ Links ]
15. LHANO, M. G., J. ADIS, M. I. MARQUES & L. D. BATTIROLA. 2005. Cornops aquaticum (Orthoptera, Acrididae, Leptysminae): aceitação de plantas alimentares por ninfas vivendo em Eichhornia azurea (Pontederiacae) no Pantanal Norte, Brasil. Amazoniana 18 (3/4): 397-404. [ Links ]
16. NUNES, A. L. 1989. Estudo sobre o ciclo de vida e fenologia de Stenacris fissicauda fissicauda (Bruner, 1908) (Orthoptera-Acrididae) em um lago de várzea da Amazônia Central, Brasil, Manaus. Tese de Mestrado. Instituto Nacional de Pesquisas da Amazônia/ Universidade Federal do Amazonas. Manaus, AM, Brasil. 122pp. [ Links ]
17. NUNES, A. L., J. ADIS & J. A. NUNES de MELLO. 1992. Estudo sobre o ciclo de vida e fenologia de Stenacris fissicauda fissicauda (Bruner, 1908) (Orthoptera: Acrididae) em um lago de várzea da Amazônia Central, Brasil. Boletim do Museu Paraense Emílio Goeldi 8 (2): 349-374. [ Links ]
18. PRANCE, G. T. 1979. Notes on the vegetation of Amazonia III. The terminology of Amazonian forest types subject to inundation. Brittonia 31 (1): 26-38. [ Links ]
19. SILVA, F. R. J., M. I. MARQUES, L. D. BATTIROLA & M. G. LHANO. 2010. Fenologia de Cornops aquaticum (Bruner) (Orthoptera: Acrididae) em Eichhornia azurea (Pontederiaceae) no Norte do Pantanal de Mato Grosso. Neotropical Entomology 3 (94): 535-542. [ Links ]
20. SCHMIDT, G. W. 1973. Primary production of phytoplancton in the three types of Amazonian waters. II. Limnology of a tropical flood-plain lake in Central Amazônia (lago Castanho). Amazoniana 4 (2): 139-203. [ Links ]
21. SOARES, M. G. M., R. G. ALMEIDA & W. J. JUNK. 1986. The trophic status of the fish fauna in Lago Camaleão, a macrophyte dominated floodplain lake in the moddle Amazon. Amazoniana 11 (4): 511-526. [ Links ]
22. WAICHMAN, A. V., C. R. GARCÍA-DÁVILA, E. R. HARDY & A. ROBERTSON. 2002. Composição do zooplâncton em diferentes ambientes do Lago Camaleão, na Ilha da Marchantaria, Amazonas, Brasil. Acta Amazonica 32 (2): 339-347. [ Links ]
23. ZOLESSI, L. C. 1956. Observaciones sobre Cornops aquaticum Br. (Acridoidea, Catantopinae) en el Uruguay. Revista de la Sociedad Uruguaya de Entomologia 1: 3-28. [ Links ]

