










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.70 no.3-4 Mendoza jul./dic. 2011
TRABAJOS CIENTÍFICOS
Sarcophagidae (Diptera) de importancia forense en la puna de Catamarca, Argentina: la ovoviviparidad como ventaja en condiciones de extrema aridez
Sarcophagidae (Diptera) of forensic importance at a high altitude desert in Catamarca, Argentina: ovoviviparity as an advantage under extreme arid conditions
Aballay, Fernando H. *, Florencia Fernández Campón*, Pablo R. Mulieri** y Silvana V. Urquiza***
*CONICET, Laboratorio de Entomología, Instituto Argentino de Investigaciones de las Zonas Áridas (IADIZA, CCT Mendoza), Casilla de correo 507, 5500 Mendoza, Argentina; Autor para correspondencia: faballay@mendoza-conicet.gob.ar.
** CONICET; ANLIS "Dr. Carlos G. Malbrán", Depto. Vectores. Av. Vélez Sársfield 563, (C1281AFR) Buenos Aires, Argentina.
***CONICET, Instituto Superior de Estudios Sociales. Instituto de Arqueología y Museo, Facultad de Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, San Martín 1545, (4000) San Miguel de Tucumán, Argentina.
Recibido: 5-V-2010
Aceptado: 13-V-2010
RESUMEN. A pesar de ser fauna necrófaga y de estar presentes en cadáveres humanos, los Sarcophagidae no suelen utilizarse en estudios forenses debido a la dificultad en su identificación y la poca información sobre su biología. En este trabajo, se identificaron las especies de Sarcophagidae asociadas a cadáveres y se estudió su relación con los estados de descomposición. Se analizaron las preferencias por los sustratos cadavéricos (cerdo y llama) y las condiciones microambientales (sol y sombra) en un ambiente de altura (3.600 msnm), durante la primavera. Se utilizaron dos cadáveres de cerdo dispuestos al sol y a la sombra y un cadáver de llama dispuesto al sol. Se colectaron 597 individuos pertenecientes a cinco especies de Sarcophagidae: M. antofagastensis Mulieri, Mariluis & Aballay (n=347), M. quimaliensis (Lopes) (n=117), M. rusca (Hall) (n=32), M. penai (Lopes) (n=5) y M. aulacophyto Pape (n=96). Tanto M. antofagastensis, como M. quimaliensis fueron las únicas especies con colonización efectiva y representaron el 78% del total de adultos colectados. Microcerella antofagastensis fue la colonizadora primaria. Ambas especies respondieron de forma similar ante las condiciones microambientales y prefirieron el cadáver de cerdo. Se discute la importancia de M. antofagastensis y M. quimaliensis, como especies indicadoras y las posibles ventajas de la ovoviviparidad frente a otras especies ovíparas, en condiciones de extrema aridez.
PALABRAS CLAVE. Insectos necrófagos; Microcerella; Ovoviviparidad; Aridez; Entomología forense.
ABSTRACT. Despite their prevalence in human corpses during decomposition, Sarcophagidae are not frequently used in forensic studies due to the difficulty in their identification and to the lack of information on their biology. In this paper, we identified the species of Sarcophagidae associated to corpses and studied their relationship to decomposition stages. We analyzed preferences for different cadaveric substrates (pig and lama) and microenvironmental conditions (shade, sun) at a site located at 3600 m.a.s.l. during the spring. Two pig carcasses, one located in the shade and the other in the sun, and a third carcass of a lama located in the sun, were used. We collected 597 individuals belonging to five Sarcophagidae species: Microcerella antofagastensis Mulieri, Mariluis & Aballay (n=347), M. quimaliensis (Lopes) (n=117), M. rusca (Hall) (n=32), M. penai (Lopes) (n=5) and M. aulacophyto Pape (n=96). Both M. antofagastensis and M. quimaliensis were the effective colonizers and represented 78% of the total adults collected. Microcerella antofagastensis was the primary colonizer. Both species had a similar response against microenvironmental conditions and both preferred the pig carcass. We discuss the importance of these two species as forensic indicators and the advantages of ovoviviparity against other oviparous species under extreme arid conditions.
KEYWORDS. Necrophagous insects; Microcerella; Ovoviviparity; Arid conditions; Forensic entomology
INTRODUCCIÓN
La entomología forense es una disciplina científica dedicada al estudio de insectos y otros artrópodos relacionados con los cadáveres, como herramientas forenses para datar decesos y en muchos casos, estimar las causas y el lugar del evento (Magaña, 2001). El propósito principal de esta disciplina es la estimación del intervalo post mortem (IPM), para lo que se utilizan principalmente dos métodos: el ciclo de vida de las especies necrófagas dominantes (generalmente utilizado en los primeros estadios del proceso de descomposición), y los patrones de sucesión faunística (utilizado mayoritariamente en los estados avanzados de descomposición) (Catts, 1992; Tabor et al., 2004; Amendt et al., 2007). El uso de entomofauna cadavérica para estimar el IPM requiere de un conocimiento de las especies involucradas en el proceso de descomposición, el ciclo de vida, su relación con los distintos estados de la descomposición y con el hábitat en el cual son registrados, por lo que es necesario contar con información básica acerca de la entomofauna en diferentes lugares geográficos (Goff, 1993).
Los estudios realizados sobre descomposición cadavérica indican que las estaciones del año y factores ambientales como, por ejemplo, la exposición al sol, influyen en la composición específica y abundancia de la entomofauna carroñera (Payne, 1965; Carvalho & Linhares, 2001; Centeno et al., 2002, 2004; Battán Horenstein et al., 2005). Por otra parte, una variable ambiental como la altitud puede tener influencia sobre la composición de especies de las comunidades de insectos necrófagos, dado que, junto con la altitud, cambian todo un conjunto de variables climáticas (temperatura, humedad, presión atmosférica, etc.) (Baumgartner & Greenberg, 1985; Martínez et al., 2007).
Los estudios de descomposición cadavérica en sitios de altura son de gran utilidad legal, debido a que, en ocasiones, los cadáveres humanos son desechados en ambientes montanos alejados de zonas urbanas (F. Aballay obs. pers.). También, son de utilidad en estudios arqueológicos, ya que las especies asociadas a cadáveres en el sitio de entierro pueden brindar información valiosa sobre la historia de los mismos (Huchet & Greenberg, 2010). En el área de estudio, se recuperaron artrópodos asociados a entierros humanos, fardos funerarios y restos de animales (Urquiza, 2010), como el caso de la estructura funeraria agropastoril, donde los cuerpos estaban acompañados por camélidos (Babot et al., 2009). Por esta razón, los estudios de la entomofauna cadavérica permiten la comparación de ésta con aquella encontrada en contextos funerarios y en los restos animales provenientes de sitios arqueológicos (Urquiza, inéd.).
Las familias de dípteros de importancia forense más abundantes son Calliphoridae (generalmente las primeras colonizadoras), Sarcophagidae y Muscidae (Battán Horenstein et al., 2007). Debido a su gran abundancia y a que sus estadios inmaduros usan los cadáveres como fuente de alimento para su desarrollo, estas familias son consideradas los descomponedores más importantes (Battán Horenstein et al., 2010).
Los sarcofágidos son elementos muy importantes del componente necrófago, debido a que ciertas especies aparecen frecuentemente en cadáveres humanos (Goff, 1991; Anderson, 1995; Oliva 1997). Sin embargo, esta familia ha sido de poca utilidad en la estimación del IPM debido a la falta de trabajos taxonómicos y biológicos. Romera et al. (2003) sostienen que la imposibilidad de identificar las especies de esta familia y la falta de conocimiento de su biología hacen cada vez más necesario realizar estudios de este grupo. En la mayoría de los estudios en los que se los menciona, los ejemplares de Sarcophagidae han sido identificados solamente hasta familia y sólo en algunos casos hasta género (Bourel et al., 1999; Wolff et al., 2001; Grassberger & Frank, 2004; Battán Horenstein et al., 2005, Segura et al., 2009; Aballay et al., 2008; Battán Horenstein et al., 2010). Pero, en un estudio realizado en Finlandia en el que la determinación fue realizada hasta especie (e.g., Sarcophaga (Robineauella) caerulescens Zetterstedt; Pohjoismáki et al., 2010), ésta no pudo utilizarse en la estimación del IPM, debido a la falta de conocimiento de su biología.
Existen estudios para Argentina en los que se han registrado Sarcophagidae durante el proceso de descomposición de diferentes sustratos cadavéricos (en cadáveres humanos encerrados, Oliva, 1997; en pollos, Battán Horenstein et al., 2005; en cerdo, Battán Horenstein et al., 2010; y en cerdo, perro y zorro, Aballay et al., 2008). No obstante, debido a la dificultad en la determinación de adultos y a que, hasta el momento, es imposible determinar larvas a nivel de especie en esta familia (Oliva, 2007); el registro de estadios larvales de Sarcophagidae sobre cadáveres humanos no permite realizar peritaciones entomológico forense precisas. En cuanto a los adultos, sólo existe una publicación científica reciente que permite determinar los adultos de Sarcophagidae para la provincia de Buenos Aires (Mulieri et al., 2010), aunque aún existen pocos estudios acerca de las especies de Sarcophagidae en gran parte del país. En cuanto a la biología de esta familia, los datos también son muy escasos (Martínez-Sánchez et al., 2000) y sólo existe información de algunas especies del género Sarcophaga Meigen (Romera et al., 2003). Lamentablemente, estos estudios están frecuentemente restringidos a registros aislados, por lo que la biología de las especies es en gran medida desconocida.
En este trabajo, se realizó un experimento de descomposición cadavérica en un ambiente de altura en la provincia de Catamarca, durante la primavera. El objetivo de este trabajo fue determinar las especies de Sarcophagidae de importancia forense y conocer su biología. En particular, se pretendió profundizar en el conocimiento de estas especies en relación a los estados de descomposición cadavérica, y establecer sus preferencias por diferentes sustratos cadavéricos (cerdo y llama) en diferentes condiciones microambientales (sol y sombra).
MATERIAL Y MÉTODOS
El estudio se realizó en un sitio ubicado en la localidad de Punta de la Peña, departamento de Antofagasta de la Sierra, provincia de Catamarca (26º 01' S, 67º 20' W) a, aproximadamente, 3.600 msnm, en la Puna Austral Argentina; dominio andino patagónico de la provincia biogeográfica puneña (Cabrera & Willink, 1973). Se trata de un desierto de altura con clima árido y seco, intensa radiación solar, gran amplitud térmica (superior a 30ºC), baja presión atmosférica y porcentaje de humedad, alta evaporación y escasas precipitaciones (inferiores a los 200 mm anuales). La vegetación dominante es la estepa arbustiva formada por matas de Acantholippia sp., entre las cuales el suelo está desnudo la mayor parte del año (Fig. 1). Otras especies típicas son Adesmia sp., Festuca orthophilla y Stipa chrysophylla (Cabrera, 1976).

Fig. 1. Área de estudio con trampa Malaise modificada.
En el área de estudio, se encuentra un sitio arqueológico donde se encontró un entierro pastoril con restos humanos y de animales domésticos, como camélidos y roedores (Babot et al., 2009). En este lugar, fueron colocados dos cadáveres de cerdo doméstico, Sus scrofa L, uno dispuesto al sol (18 kg, 26º 01' 32.3'' S, 67º 20' 36.5'' W, 3.578 msnm) y otro a la sombra (17 kg, 26º 01' 38.2'' S, 67º 20' 31.6'' W, 3.595 msnm) entre un farallón ignimbrita, lo que permitía que el cadáver estuviera a la sombra durante todo el día. Además, se colocó un tercer cadáver de llama, Lama glama, (28 kg, 26º 01' 33.4'' S, 67º 20' 42.5'' W, 3.585 msnm), dispuesto al sol. Los cadáveres se encontraban separados por más de 100 m. Los tres animales fueron sacrificados en el sitio de estudio, utilizando el método de estocada en el corazón para cerdos (Centeno et al., 2002) y desmedulado y posterior degüello para la llama. Cada cadáver fue colocado en el interior de un armazón de madera recubierto con alambre tejido (120 X 100 X 150 cm.), para permitir el ingreso de la entomofauna cadavérica y evitar la acción de vertebrados carroñeros. Las jaulas con los cadáveres fueron colocadas sobre 5 cm de tierra suelta previamente mezclada, manteniendo las mismas condiciones del terreno bajo cada cadáver. En el interior de cada jaula y sobre cada cadáver, se incluyó una trampa Malaise modificada (Fig. 1). El tul se dispuso a 10 - 15 cm de la superficie para permitir el ingreso de la entomofauna cadavérica hacia el interior de la trampa.
Se muestrearon adultos de dípteros diariamente durante 25 días seguidos, desde el 17/11/2009 hasta la finalización de la descomposición, utilizando una trampa Malaise modificada en cada cadáver. En aquellos muestreos en los que se registró presencia de larvas de dípteros, las mismas se muestrearon utilizando un sacabocado. La cantidad de larvas muestreadas con sacabocado es una pequeña parte de la masa larvaria presente en los cadáveres, por lo que se asume que esto no interfiere con el proceso de descomposición de los cadáveres, y que es una muestra representativa de las especies de larvas presentes en los cadáveres. La mitad de esta muestra larvaria se destinó a cría en recipientes de 500 cm3 con carne picada, envuelta en papel de aluminio, dispuesta en el fondo del recipiente y recubierta con aserrín de madera y una película fina de arena hacia la superficie. El extremo superior del recipiente estaba cubierto por una doble red de 300 μm de apertura de malla. La otra mitad de larvas fue muerta con agua caliente (80ºC a 90ºC), conservadas en alcohol al 70% y depositadas en la colección de entomología del IADIZA, CCT-Mendoza. Los dípteros adultos capturados en la trampa Malaise fueron sacrificados con acetona y posteriormente, dispuestos en camas entomológicas y recipientes herméticos.
En cada muestreo, se registró la temperatura y la humedad ambiente (máxima y mínima) con un termohigrómetro digital Tfa, que se dispuso en forma permanente a aproximadamente a 30 cm de cada cadáver. También, se registraron las temperaturas corporales internas (vía rectal) de los tres cadáveres con un termómetro digital de punción, diariamente y aproximadamente a la misma hora (alrededor del mediodía).
La determinación de las especies de Sarcophagidae se realizó mediante el estudio de la genitalia de los machos, utilizando la técnica de disección, consistente en estirar los segmentos genitales, mediante el uso de alfileres entomológicos, para exponer el phallus (Lopes, 1973; Dahlem & Naczi, 2006). Los especímenes así tratados fueron mantenidos en ambiente seco durante 2-3 días para mantener su terminalia en esa posición y ser estudiada. Las hembras fueron asociadas a los machos de acuerdo a su similitud morfológica y al estudio y comparación de sus esternitos genitales. La identificación de los estados de descomposición se basó en los descriptos por Payne (1965): fresco, hinchado, descompuesto activo, descompuesto avanzado, restos.
Se utilizó el método ANOVA de un factor para comparar las condiciones microambientales de los sitios donde estaban localizados los cadáveres al sol y a la sombra, y la temperatura interna de los tres cadáveres. Además, para comparar la duración de los estados del proceso de descomposición de los tres cadáveres, se utilizó una tabla de contingencia. Finalmente, para examinar el efecto de sustrato cadavérico (llama vs. cerdo) y microambiente (sol vs. sombra) sobre la abundancia de las especies, se realizaron ANOVAs de dos factores. Debido a la falta de normalidad en la distribución de los datos, estos fueron transformados como Log X + 1 para los análisis de abundancia. Para todos los análisis se utilizó el programa GenStat V. 13.3.0.5165.
RESULTADOS
Las condiciones de temperatura y humedad para los cadáveres situados al sol y a la sombra fueron similares, excepto para las temperaturas mínimas, las cuales fueron mayores para la sombra (Tmin sombra = 7,39 ºC + 0,42 ºC; Tmin sol = 3,33 ºC + 0,53 ºC; F1,48 = 35,84; P = 0.00). El promedio de temperaturas máximas fue de 41,90 ºC (+ 0,93 ºC), mientras que para la humedad promedio de la máxima fue del 33,44 % (+ 1,54%) y de la mínima fue del 12,46% (+ 0,47%). Probablemente, a causa de las condiciones extremas de humedad ambiente, los orificios naturales de los cadáveres se secaron rápidamente. A pesar de haberse registrado los cinco estadios descriptos por Payne (1965), recién en el estadio hinchado, se produjo la liberación de líquidos tisulares del cuerpo por los orificios naturales junto con la liberación de los gases. Aunque no se registraron grandes diferencias en las condiciones microambientales, la temperatura interna de los cadáveres fue mayor para aquellos situados al sol que para el cadáver de cerdo situado en la sombra
(Tcerdo sombra = 18,56 ºC + 1,35 ºC; Tcerdo sol = 32,36 ºC + 1,35 ºC; Tllama sol = 29,70 ºC + 1,53 ºC; F2,71 = 27,18; P = 0.00). A pesar de que no se registraron diferencias significativas en la duración de los estados de descomposición entre los diferentes sustratos cadavéricos (χ² = 1,2584; gl = 6; P = 0,973), existe una tendencia a una menor duración en los sustratos ubicados al sol (Tabla I).
Tabla I. Las especies Microcerella antofagastensis y M. quimaliensis (Sarcophagidae) registradas sobre cadáveres de cerdos (sol, sombra) y llama, en relación a los estados de descomposición (F= Fresco; Des.=Descomposición; A: adulto; L: adultos emergidos de los cultivos larvales.; M: macho; H: hembra).
Se colectó un total de 597 individuos adultos de Sarcophagidae durante todo el proceso de descomposición en los tres cadáveres, pertenecientes a 5 especies: Microcerella antofagastensis Mulieri, Mariluis & Aballay (n=347), Microcerella quimaliensis (Lopes) (n=117), Microcerella rusca (Hall) (n=32), Microcerella penai (Lopes) (n=5) y Microcerella aulacophyto Pape (n=96). De las larvas utilizadas para el experimento de cría, solo M. antofagastensis y M. quimaliensis desarrollaron sus estadios inmaduros (nM.antofa. = 92; nM.quimal. = 37), esto indica que son colonizadoras efectivas. Estas dos especies representaron el 78% del total de adultos Sarcophagidae colectados en las trampas Malaise. Las hembras adultas de M. antofagastensis fueron las colonizadoras primarias en los cadáveres de cerdo, tanto al sol como a la sombra (Tabla I). Sin embargo, en el cadáver dispuesto a la sombra aparecieron en el estado fresco, mientras que en el cadáver dispuesto al sol aparecieron en el estado hinchado (Tabla I). Por otro lado, Microcerella quimaliensis apareció en días posteriores a M. antofagastensis, en el estado hinchado en los dos cadáveres de cerdos, comportándose como especie secundaria. En el cadáver de llama, las dos especies colonizadoras fueron registradas en el mismo día durante el estadio hinchado. También se colectaron machos de M. antofagastensis, los cuales fueron colectados en días posteriores a la aparición de las hembras de esta especie. En el caso de M. quimaliensis, no se registraron machos ni se obtuvieron adultos a partir de larvas criadas en la llama (Tabla I).
La variación de la abundancia de ambas especies de moscas en los tres cadáveres se muestra en la Tabla I. Ninguna de las dos especies presentó abundancias diarias mayores a dos individuos en el cadáver de llama. Para los cadáveres de cerdo, M. antofagastensis presentó la máxima abundancia de adultos en los dos cadáveres en el estadio hinchado (cerdosombra: día 5, n = 44; cerdosol: día 7, n =69). En el caso de las larvas criadas, la máxima abundancia de adultos emergentes se registró al final del estadio hinchado y comienzos del estadio de descomposición activa, en el día 7. Para los adultos de M. quimaliensis, la máxima abundancia en el cadáver dispuesto al sol se registró más tarde, a fines del estadio hinchado y comienzos del estado de descomposición activa, entre los días 7 y 10. Para el cadáver de cerdo dispuesto a la sombra, la máxima abundancia se dio más temprano, durante el estadio hinchado (día 5). Los adultos que emergieron de larvas criadas no presentaron gran abundancia en el cerdo dispuesto al sol, mientras que la mayor abundancia en el cerdo dispuesto a la sombra ocurrió en el día 15, al final del estadio de descomposición activa.
La abundancia diaria de adultos de M. antofagastensis fue casi el doble de la de M. quimaliensis (mediaM.antofa.= 0,299, mediaM. quimal.=0,159; F1, 96 =9,630; P = 0,010), pero no se registraron diferencias en abundancia entre los cadáveres de cerdo ubicados al sol y a la sombra (media cerdo sol=0.236, media cerdo sombra=0.222; F1,196 =0,070; P = 0,790) (Fig. 2). Tanto M. antofagastensis como M. quimaliensis desarrollaron sus estadios inmaduros en los cerdos ubicados en ambos microambientes (sombra y sol). La abundancia de larvas no fue diferente entre microambientes (media cerdo sol=0.059, media cerdo sombra=0.108; F1,196=2,060; P =0,152), ni entre especies (mediaM.antofa.= 0.110, mediaM. quimal.=0.057; F196,1,0.5=2,060; P=0,125); aunque M. antofagastensis presentó una tendencia a una abundancia mayor, similar a lo que ocurrió con los adultos.
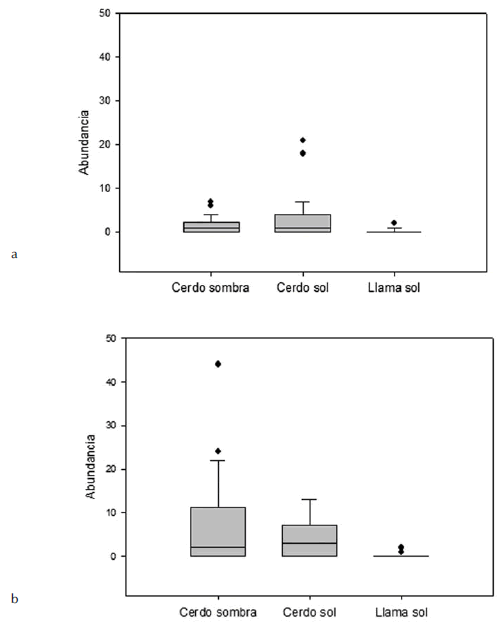
Fig. 2. Abundancia diaria de (a) Microcerella antofagastensis y (b) M. quimaliensis en los diferentes sustratos cadavéricos
Al examinar las abundancias de las dos especies de Sarcophagidae en diferentes sustratos (llama y cerdo) ubicados al sol, se observó un mayor número de adultos en el cerdo que en la llama (media cerdo sol = 0,236, mediallama = 0,028; F1,196 = 26,590; P = 0,001) (Fig. 2), pero no existieron diferencias en las abundancias de las dos especies de moscas (mediaM.antofa. = 0.161, mediaM.quimal. = 0.103; F1,196 = 2,080; P = 0,151). Por otro lado, la abundancia larvaria sobre el cadáver de cerdo fue mayor que en la llama (media cerdo sol=0.059, media llama=0.009; F1,196 = 4,280; P = 0,005), M. antofagastensis fue más abundante que M. quimaliensis (mediaM.antofa. = 0,076, mediaM.quimal. = 0,009; F1,196 = 8,050; P = 0,005).
Finalmente, la proporción de sexos de los individuos adultos colectados en trampas Malaise estaba sesgada hacia las hembras para ambas especies de Microcerella, mientras que, en los adultos obtenidos a partir de las larvas colectadas en los cadáveres, la proporción de sexo no estaba sesgada hacia ningún sexo (Fig.3).

Fig. 3. Abundancia total de (a) Microcerella antofagastensis y (b) M. quimaliensis por sexo en individuos colectados como adultos y en adultos que emergieron de larvas criadas en los diferentes sustratos cadavéricos.
DISCUSIÓN Y CONCLUSIONES
En este estudio sobre Sarcophagidae asociados a distintos sustratos cadavéricos, registramos la presencia de cinco especies, todas ellas pertenecientes al género Microcerella. Tres de estas especies: M. rusca, M. penai y M. aulacophyto han sido registradas en ambientes áridos como la estepa patagónica y la puna jujeña (Mariluis, 2002). Sin embargo, M. antofagastensis y M. quimaliensis no habían sido registradas previamente sobre cadáveres en descomposición en la Argentina, y constituyen nuevos registros para la provincia de Catamarca.
El género Microcerella Macquart presenta mayor diversidad en los ambientes secos y de altura en Sudamérica (Pape, 1990; Mulieri & Mariluis, 2009). En la provincia de Córdoba, Battán Horenstein et al. (2010) registraron una especie no determinada de Microcerella sobre cadáveres de cerdo. Previamente, dos especies, M. austrohartigia Pape y M. muehni (Blanchard), fueron criadas artificialmente a partir de carne en descomposición (Oliva, 1997; Mulieri et al., 2010) o colectados sus adultos con cebos (Mariluis et al., 2007; Mulieri et al., 2008) en la provincia de Buenos Aires. Esto sugiere que el género Microcerella podría contener más especies que formen parte de la fauna cadavérica. Además, pueden existir diferencias en la composición de especies asociadas a los cadáveres en los diferentes ambientes (Centeno et al., 2004, Battán Horenstein et al., 2005). En estudios llevados a cabo en Colombia, se encontraron diferencias en la composición de la fauna cadavéricas en función de la altura. Martínez et al. (2007) las encontraron entre un ambiente de altura localizado a 3.500 msnm y la fauna registrada a 1.450 msnm por Wolff et al. (2001) y Pérez et al. (2005), en el Páramo colombiano.
Tanto M. antofagastensis como M. quimaliensis fueron las únicas especies con colonización efectiva en los cadáveres, M. antofagastensis fue la más abundante. Ambas especies respondieron de forma similar ante las condiciones microambientales y tipos de sustratos cadavéricos, prefiriendo el cerdo frente a la llama (las dos especies fueron más abundantes en los cadáveres de cerdo tanto al sol como a la sombra). La falta de respuesta ante las condiciones microambientales, probablemente, se deba a que estas condiciones no fueron tan diferentes. El cadáver de cerdo en la sombra estaba ubicado junto a un farallón de ignimbrita, quizá este farallón actuó como amortiguador en la variación de temperatura a lo largo del día, e hizo que las condiciones de temperatura ambiente fueran muy similares para los dos cadáveres de cerdo. Sin embargo, sí existieron diferencias en las temperaturas internas de los sustratos cadavéricos dispuestos al sol, lo que podría ser responsable de la tendencia a la menor duración de los estados de descomposición en estos sustratos (Tabla I).
Las especies M. antofagastensis y M. quimaliensis presentaron un sesgo hacia las hembras en la proporción de sexos de adultos, mientras que en la proporción de sexos de larvas criadas hasta adultos no existe tal sesgo. Esto reflejaría la atracción de las hembras por los cadáveres como sitio de cría.
Los cadáveres de cerdo son el modelo animal más utilizado en los estudios forenses debido a que el proceso de descomposición es muy similar al del humano (Payne, 1965; Goff, 1993). Este hecho, junto a la preferencia por el sustrato cerdo encontrada en nuestro estudio, señalan las especies de Sarcophagidae, M. antofagastensis y M. quimiliensis, como potenciales indicadoras forenses en ambientes áridos, durante la primavera.
Microcerella antofagastensis fue la especie colonizadora primaria, ocurrió el primer día en el estadio fresco del cerdo localizado a la sombra (Tabla I). Estos resultados no coinciden con otros estudios en los que las especies de Microcerella aparecieron durante el estadio hinchado, descomposición avanzada, y se comportaron como colonizadoras terciarias (Battán Horenstein et al., 2010). Las especies de Sarcophagidae poseen la característica de depositar larvas de primer estadio sobre los sustratos de cría (Barros et al., 2008; Pape & Dahlem, 2010). Esta característica les confiere una ventaja adaptativa en comparación con las especies ovíparas, en especial cuando se trata de sustratos efímeros, como son los casos de pequeños cadáveres o cadáveres en condiciones de rápida desecación como los de este experimento. Probablemente, esta característica fue la que permitió a las especies de esta familia tener una colonización efectiva sobre el cadáver, inmediatamente después del deceso, en comparación con la fauna cadavérica de dípteros ovíparos (como los Calliphoridae). A pesar de que, inmediatamente después del deceso, se registraron adultos de Calliphoridae en las trampas Malaise y se observaron huevos en los orificios naturales de los cadáveres, a las pocas horas los mismos estaban secos (F. Aballay obs. pers.) y no se detectaron larvas de Calliphoridae en los primeros días de la descomposición; recién se registraron larvas de esta familia al final del estado hinchado, cuando el cadáver libera líquidos tisulares en forma permanente (F. Aballay en preparación). Las condiciones climáticas extremas del área de estudio (humedad máxima del 33%, amplitud térmica de más 30ºC y fuertes vientos), desde el primer día de la descomposición, provocaron que se secaran los orificios naturales de los cadáveres, probablemente, dificultando la colonización efectiva por dípteros ovíparos en los primeros días de la descomposición. Sin embargo, las larvas de Sarcophagidae que se desplazaron hacia las partes más profundas de las cavidades, pudieron colonizar los cadáveres en forma efectiva inmediatamente producido el deceso.
Barros et al. (2008) consideran que la ventaja que la ovoviviparidad les confiere a los Sarcophagidae, hace que esta familia sea de principal importancia en los estudios forenses. Esto sería particularmente importante en ambientes áridos, donde las condiciones ambientales de humedad y la amplitud térmica extremas les confieren una ventaja, dado que, luego de la puesta, las larvas ya pueden desplazarse a sitios adecuados para su desarrollo dentro del cadáver.
Lamentablemente, la escasez de claves para la identificación de especies junto con el hecho de que las claves están desarrolladas para la identificación de machos, hace que no siempre puedan utilizarse los Sarcophagidae en estudios forenses, con lo que se pierde una valiosa herramienta. De allí la necesidad de estudios que se focalicen en esta familia, tanto en la sistemática de formas adultas e inmaduras como en su biología.
Éste es el primer trabajo donde se identifican especies de Sarcophagidae de importancia forense en los diferentes estados de descomposición, en los ambientes de altura en la puna Argentina. El mismo señala a M. antofagastensis y M. quimaliensis como potenciales indicadoras de IPM. Además, al actuar como colonizadoras primarias, serían de gran utilidad para estimar el tiempo de deceso en forma precisa, durante los estados tempranos de la descomposición. Para lograr esto último, es necesario realizar experimentos de cría de ambas especies a diferentes temperaturas y que incluya la descripción de los estadios inmaduros. Las especies de Sarcophagidae son principalmente abundantes en las estaciones cálidas (primavera, verano) (Battan Horenstein et al., 2010). Esto probablemente sea más acentuado en ambientes con condiciones extremas, como lo son los ambientes de altura. Es por eso que sería necesario complementar el presente trabajo con estudios realizados en las otras estaciones del año, en particular durante el verano.
AGRADECIMIENTOS
Durante el curso de este estudio F. H. Aballay recibió el apoyo de un programa de becas de postgrado del CONICET. Este trabajo es parte de su proyecto de doctorado, bajo la supervisión del Dr. G. E. Flores y Dr. N. Centeno. Esta investigación se desarrolló en el marco del proyecto FONCyT-PICT 38127, dirigido por C. Aschero. Agradecemos al Dr. G. E. Flores, por la lectura y las sugerencias del manuscrito, así como por su colaboración en el experimento de campo. Agradecemos a la familia Morales y especialmente a Jacoba, por su buena predisposición para llevar a cabo este experimento.
BIBLIOGRAFÍA CITADA
1. ABALLAY, F. H., A. F. MURÚA, J. C. ACOSTA & N. D. CENTENO. 2008. Primer registro de artropodofauna cadavérica en sustratos humanos y animales en San Juan, Argentina. Rev. Soc. Entomol. Argent.67 (3-4): 157-163. [ Links ]
2. AMENDT, J., C. P. CAMPOBASSO, E. GAUDRY, C. REITER, H. N. LEBLANC & M. J. R. HALL. 2007. Best practice in forensic entomology standards and guidelines. Int. J. Legal Med. 121: 90-104. [ Links ]
3. ANDERSON, G. S. 1995. The use of insects in death investigations: an analysis of forensic entomology cases in British Columbia over a five year period. Can.Soc. Forensic Sci. J. 28 (4):277-292. [ Links ]
4. ANDERSON, G. S. 2001. Insect succession on carrion and its relationship to determining time since death. En: Byrd J. H. & J. L. Castner (eds.), Forensic entomology: the utility of arthropods in legal investigations, CRC Press, Boca Raton London New York Washington, pp.143-175. [ Links ]
5. BABOT, M. P., L. G. GONZÁLEZ BARONI, S. V. URQUIZA, M. G. AGUIRRE, M. G. COLANERI, S. HOCSMAN & M. C. HAROS. 2009. Dinámicas de formación y transformación de un entierro en el desierto puneño (Antofagasta de la Sierra, Puna Meridional Argentina). Intersecciones Antropol. 10: 183-201. [ Links ]
6. BARROS, R M. de., C. A. de MELLO-PATIU, J. R. PUJOL-LUZ. 2008. Sarcophagidae (Insecta, Diptera) associados à decomposição de carcaças de Sus scrofa Linnaeus (Suidae) em área de Cerrado do Distrito Federal, Brasil. Rev. Bras. Ent. 52: 606-609. [ Links ]
7. BATTÁN HORENSTEIN, M., M. ARNALDOS, B. ROSSO & D. GARCÍA. 2005. Estudio preliminar de la comunidad sarcosaprófaga en Córdoba (Argentina): aplicación a la entomología forense. Anal. Biol. 27: 191-201. [ Links ]
8. BATTÁN HORENSTEIN, M., A. X. LINHARES, B. ROSSO & D. GARCÍA. 2007. Species composition and seasonal succession of saprophagous calliphorids in a rural area of Córdoba, Argentina. Biol. Res. 40: 163-171. [ Links ]
9. BATTÁN HORENSTEIN, M., A. X. LINHARES, B. ROSSO DE FERRADAS & D. GARCÍA. 2010. Decomposition and dipteran succession on pig carrion in central en Argentina: Ecological aspects and their importance to forensic science. Med. Vet. Entomol. 24: 16-25. [ Links ]
10. BAUMGARTNER, D. L. & B. GREENBERG.1985. Distribution and medical ecology of blow flies (Diptera: Calliphoridae) of Peru. Ann. Entomol. Soc. Am. 78: 565-587. [ Links ]
11. BOUREL, B., L. M. BOUYER, V. HEDOUIN, J. C. CAILLIEZ, D. DEROUT & D. GOSSET. 1999. Necrophilous insect succession on rabbit carrion in sand dune habitats in northern France. J. Med. Entomol. 36: 420-425. [ Links ]
12. CABRERA, A. L. 1976. Regiones fitogeográficas argentinas. En: Kugler, W. F. (ed.): Enciclopedia Argentina de Agricultura y Jardinería. 2º ed. Tomo II. Fasc. I. Acme, Bs. As. [ Links ]
13. CABRERA, A. & A. WILLINK. 1973. Biogeografía de América Latina. Monografía 13. Serie Biología OEA. Washington, DC: Secretaria General de la OEA. [ Links ]
14. CARVALHO, L. M. L. & A. X. LINHARES. 2001. Seasonality of insect succession and pig carcass decomposition in a natural forest area in southeastern Brazil. J. Foren. Sci. 46: 604-608. [ Links ]
15. CATTS, E. P. 1992. Problems in estimating the PMI in death investigations. J. Agric. Entomol. 9: 245-255. [ Links ]
16. CENTENO, N. D., M. MALDONADO & A. OLIVA. 2002. Seasonal patterns of arthropods occurring on sheltered and unsheltered pig carcasses in Buenos Aires province (Argentina). Forensic Sci. Int.126: 63-70. [ Links ]
17. CENTENO, N. D., D. ALMORZA & C. ARNILLAS. 2004. Diversity of Calliphoridae (Insecta: Diptera) in Hudson, Argentina. Neotrop. Entomol. 33: 387-390. [ Links ]
18. DAHLEM, G. A. & R. F. C. NACZI. 2006. Flesh flies (Diptera: Sarcophagidae) associated with North American pitcher plants (Sarraceniaceae), with descriptions of three news species. Ann. Entomol. Soc. Am. 99: 218-240. [ Links ]
19. GOFF, M. L. 1991. Comparison of insect species associated with decomposing remains recovered inside dwellings and outdoors on the Island of Oahu, Hawaii. J. Forensic Sci. 36 (3): 748-753. [ Links ]
20. GOFF, M. L. 1993. Estimation of postmortem interval using arthropod development and successional patterns. Forensic Sci. Review 5: 81-94. [ Links ]
21. GRASSBERGER, M. & C. FRANK. 2004. Initial Study of Arthropod Succession on Pig Carrion in a Central European Urban Habitat. J. Med. Entomol. 41(3): 511-523. [ Links ]
22. HUCHET, J. B. & B. GREENBERG. 2010. Flies, Mochicas and burial practices: a case study from Huaca de la Luna, Peru. J. Archaeol. Sci. 37: 2846-2856. [ Links ]
23. LOPES, H. S. 1973. Collecting and rearing sarcophagid flies (Diptera) in Brazil during forty years. An. Acad. Bras. Cienc. 45: 279-291. [ Links ]
24. MAGAÑA, C. 2001. La entomología forense y su aplicación a la medicina legal. Data de muerte. Bol. Soc. Entomol. Arag. 28:49-57. [ Links ]
25. MARILUIS, J. C. 2002. A new species and new records of Microcerella Macquart (Diptera: Sarcophagidae) from Argentinean Patagonia. Proc. Entomol. Soc.Wash.104:91-96. [ Links ]
26. MARILUIS, J. C., J. C. SCHNACK, P. R. MULIERI & J. P. TORRETTA. 2007. The Sarcophagidae (Diptera) of the coastline of Buenos Aires city, Argentina. J. Kansas Entomol. Soc.. 80: 243-251. [ Links ]
27. MARTÍNEZ, E., P. DUQUE & M. WOLFF. 2007. Succession pattern of carrion-feeding insects in Páramo, Colombia. Forensic Sci. Int.166: 182-189. [ Links ]
28. MARTÍNEZ-SÁNCHEZ, A., S. ROJO & M. GARCÍA. 2000. Sarcofágidos necrófagos y coprófagos asociados a un agrosistema de dehesa (Diptera, Sarcophagidae). Bol. Asoc. Esp. Entomol. 24: 171-185. [ Links ]
29. MULIERI, P. R., & J. C. MARILUIS. 2009. New species and new records of Microcerella Macquart (Diptera: Sarcophagidae) belonging to the M. spinigena species-group. Neotrop. Entomol. 38: 847-849. [ Links ]
30. MULIERI, P. R, J. C. MARILUIS & L. D. PATITUCCI. 2010. Review of the Sarcophaginae (Diptera: Sarcophagidae) of Buenos Aires province (Argentina), with a key and description of a new species. Zootaxa 2575: 1-37. [ Links ]
31. MULIERI, P. R, J. C. SCHNACK, J. C. MARILUIS & J. P. TORRETTA. 2008. Flesh flies (Diptera: Sarcophagidae) from a grassland and a woodland in a Nature Reserve of Buenos Aires, Argentina. Rev. Biol. Trop. 56: 1287-1292. [ Links ]
32. OLIVA, A. 1997. Insectos de interés forense de Buenos Aires (Argentina). Primera lista ilustrada y datos bionómicos. Rev. Mus. Argnt. Cien. Nat. Bernardino Rivadavia Inst. Nac. Inv. Cien. Nat. 7: 13-59. [ Links ]
33. OLIVA, A. 2007. Frecuencia y distribución temporal de moscas cadavéricas (Diptera) en la ciudad de Buenos Aires. Rev Mus Argnt Cien Nat. (n.s.) 9: 5-14. [ Links ]
34. PAPE, T. 1990. Revisionary notes on American Sarcophaginae (Diptera: Sarcophagidae). Tijdschr. Entomol. 133: 43-74. [ Links ]
35. PAPE, T. & G. A. DAHLEM. 2010. Sarcophagidae (Flesh flies). In: B. V Brown, A. Borkent, J. M. Cumming, D. M. Wood, N. E. Woodley & M. A. Zumbado [Eds], Manual of Central American Diptera Vol. 2. NRC Research Press, Ottawa, Canada. pp. 1297-1335. [ Links ]
36. PAYNE, J. A. 1965. A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology 46: 592-602. [ Links ]
37. PÉREZ, S., P. DUQUE & M. WOLFF. 2005. Successional behavior and occurrence matrix of carrion-associated arthropods in the urban area of Medellín, Colombia. J. Forensic Sci. 50: 1-7. [ Links ]
38. POHJOISMÁKI, J. L. O., P. J. KARHUNEN, S. GOEBELER, P. SAUKKO & I. E. SÄÄKSJÄRVI. 2010. Indoors forensic entomology: colonization of human remains in closed environments by specific species of sarcosaprophagous flies. Forensic Sci. Int.199: 38-42. [ Links ]
39. ROMERA, E., M. I. ARNALDOS, M. D. GARCÍA & D. GONZÁLEZ-MORA. 2003. Los Sarcophagidae (Insecta, Diptera) de un ecosistema cadavérico en el sureste de la Península Ibérica. Anal. Biol. 25: 49-63. [ Links ]
40. SEGURA, N. A., W. USAQUÉN & M. C. SÁNCHEZ. 2009. Succession pattern of cadaverous entomofauna in a semi-rural area of Bogotá, Colombia. Forensic Sci. Int. 187: 66-72. [ Links ]
41. TABOR, K., C. BREWSTER & R. FELL. 2004. Analysis of the successional patterns of insects on carrion in southwest Virginia. J. Med. Entomol. 41: 785-795. [ Links ]
42. URQUIZA, S. V. Inéd. Arqueofaunas del Alero Punta de la Peña 4: Implicaciones para el Manejo del Recurso Camelidae en Antofagasta de la Sierra, Puna Meridional, Catamarca. Tesis Doctoral, Facultad de Bioquímica, Química y Farmacia, Universidad Nacional de Tucumán, 2009. [ Links ]
43. URQUIZA, S. V. 2010. Entomología y Química en el Contexto Arqueológico: Nuevos Elementos para el Entendimiento de la Historia Tafonómica del Registro Arqueofaunístico en Antofagasta de la Sierra, Catamarca. En: De Nigris, M., P. Fernández, M. Giardina, A. Gil, M. Gutiérrez, A. Izeta, G. Neme & H. Yacobaccio (eds.) Zooarqueología a principios del siglo XXI: aportes teóricos, metodológicos y casos de estudio, Malargue, pp. 429-437. [ Links ]
44. WOLFF, M., A. URIBE, A. ORTIZ & P. DUQUE. 2001. A preliminary study of forensic entomology in Medellín, Colombia. Forensic Sci. Int.120: 53-59. [ Links ]

