










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
Print version ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.71 no.1-2 Mendoza Jan./June 2012
TRABAJOS CIENTÍFICOS
Diuraphis noxia (Hemiptera: Aphididae): Identificación de los biotipos presentes en poblaciones argentinas
Diuraphis noxia (Hemiptera: Aphididae): Identification of biotypes present in populations of Argentina
Ricci, Mónica *, Mehmet Cakir ** y Ana M. Castro *, ***
* Zoología Agrícola. Facultad de Ciencias Agrarias y Forestales. UNLP. 60 y 119 CC 31 (1900). La Plata. Bs. As. Argentina; e-mail: mricci@agro.unlp.edu.ar
** WA State Agricultural Biotechnology Centre, Faculty of Sustainability, Environmental and Life Sciences, Murdoch University, Murdoch, WA 6150, Australia
*** Instituto de Fisiología Vegetal (INFIVE)-CONICET CC 327, La Plata (1900), Buenos Aires, Argentina; e-mail: amcastro@isis.unlp.edu.ar
Recibido: 27-VII-2011
Aceptado: 22-IV-2012
RESUMEN. El objetivo del trabajo fue evaluar el comportamiento de líneas de trigo y cultivares diferenciales, portadores de genes de resistencia, a fin de determinar la composición biotípica del áfido Diuraphis noxia Kurdjumov, "Pulgón Ruso del Trigo" (PRT), en Argentina; y se identifican aquellos hospederos con resistencia genética al PRT. Los 18 cultivares diferenciales y líneas Novel, se sembraron en invernáculo, en un diseño aleatorio con 2 repeticiones. Las observaciones se realizaron a los 21 y 51 días de la infestación, se determinó el grado de clorosis y de enrollamiento foliar. Los resultados se analizaron con ANOVA y el Test de Tuckey (a=0,05). A los 21 días, las líneas 14 y 1 (Noveles) presentaron el mejor comportamiento, pero se desconocen sus genes de resistencia. Los cultivares portadores de Dn2 y Dn7 resultaron con un alto nivel de tolerancia, mientras que Dn1 y dn3 fueron moderadamente tolerantes y las portadoras de Dn4 y Dn5, muy dañados por las poblaciones Argentinas de PRT. A los 51 días, solo Dn7 y las líneas 14 y 1 mantuvieron un comportamiento de tolerancia al áfido. Se infiere que en el país se encontrarían los biotipos 1, 2* y 5*, estos dos últimos con características diferentes a los identificados en EE.UU. Se concluye que los cultivares mejorados en otros países, pueden comportarse como susceptibles frente a las poblaciones locales.
PALABRAS CLAVE. Resistencia genética; Trigo; Pulgón Ruso del Trigo; Cultivares diferenciales.
ABSTRACT. The aim of the current research was to assess the plant responses of wheat lines and differential cultivars carrying Diuraphis noxia Kurdjumov resistance genes when subjected to aphid infestation, in order to determine the biotypic composition of RWA populations in Argentina. Eighteen lines and differential cultivars were sown in a greenhouse in a randomized design with two replicates. Two recordings of the chlorotic damage and the level of leaf rolling were made after 21 and 51 days from the infestation onset. The results were analyzed with an ANOVA and Tuckey Test (a=0.05). After 21 days of infestation, two novel lines (14 and 1) showed the highest level of tolerance, with minor chlorotic damage. The differential cultivars carrying Dn2 and Dn7 resistance genes also were highly tolerant, Dn1 and dn3 resistance genes were moderately tolerant and those ones carrying Dn4 and Dn5 genes resulted seriously damaged. After 51 days of infestation, only the differential cultivar carrying Dn7 and the novel lines 14 and 1 maintained a high level of tolerance. Based on the current results, the RWA biotypes 1, 2* and 5* are present in Argentina, the last ones with different characteristics from biotypes 2 and 5 identified in EEUU. We conclude that improved cultivars in other countries can behave as susceptible against local populations.
KEY WORDS. Genetic resistance; Wheat; Russian Wheat Aphid; Differential cultivars.
INTRODUCCIÓN
Diuraphis noxia Kurdjumov (Aphidinae: Macrosiphini), conocido como "Pulgón Ruso del Trigo" (PRT), es uno de los áfidos con mayor capacidad de dispersión en el mundo en las últimas décadas. Es originario de las regiones paleárticas de Europa Oriental y Asia Central que rodean al Mar Negro, Mar Caspio y la Cuenca de los Cárpatos (Jones et al., 1989; Starý, 1996). Según Kazemi (2001), el primer registro del PRT data del año 1900 en el sur de Rusia (Ucrania) por Mordvilko, a partir de ese momento su dispersión fue permanente, hacia la mayoría de las áreas de producción de cereales.
En las zonas próximas al centro de origen, el áfido fue detectado en 1959 en Turquía, durante 1989 en Macedonia y Hungría, en 1990 en Serbia, en 1993 en Polonia, en 1996 en la República Checa, en 1997 en Alemania, en 2000 en Austria, en 2001 en Eslovaquia y en 2002 en Croacia (Starý et al., 2003). Ingresó a Sudáfrica en 1977 y en la región Neártica, fue detectado primero en México en 1980 y posteriormente en EE.UU. en 1986. En este último país, el áfido se dispersó desde Texas a todos los Estados productores de trigo del oeste, en un poco más de un año, lo que causó pérdidas estimadas en 100 millones de dólares, solo durante 1988 (Hughes, 1990; Norambuena & Gerding, 1991). En Sudamérica, se detectó en la zona central de Chile en 1987 (Zerené et al., 1987) y en la Argentina, el primer hallazgo fue realizado en 1991 en Malargüe, provincia de Mendoza (Ortego & Delfino, 1992).
El PRT está adaptado a regiones de climas secos, cálidos y con lluvias esporádicas; en estas condiciones las poblaciones tienen un rápido crecimiento (Jones, et al., 1989). De acuerdo a los registros tomados en EE.UU. y en Sudáfrica, las altas densidades de población del áfido en trigo y cebada, se asocian a climas secos, delimitados por las isohietas de 300 y 400 mm (Hughes & Maywald, 1990). Estos datos son concordantes con los obtenidos por Clúa et al., (2004), quienes hallaron la mayoría de las poblaciones de Argentina dentro del mismo régimen hídrico, a excepción de unas pocas que se localizaron en áreas con 2.000 mm de precipitaciones.
Desde el año 1978, en que al PRT se lo identificó como una plaga grave en el cultivo de trigo y cebada en Sudáfrica y posteriormente en los EE.UU., los mejoradores de plantas se abocaron a la búsqueda de fuentes de resistencia (Nkongolo et. al., 1990). Du Toit & Van Niekerk (1985) determinaron que las fuentes de resistencia a D. noxia existen en las especies ancestrales de trigo diploide Triticum monococcum, T. timopheevi, T. dicoccoides y T. tauschii, y en los trigos anfiploides de T. monococcum y T. durum. Otras fuentes se identificaron en centeno (Secale cereale) y triticale (Nkongolo et al, 1989; Webster, 1990).
Gracias a la búsqueda de fuentes de resistencia en trigos hexaploides, que pueden ser utilizados fácilmente en los programas de mejoramiento, se informó por primera vez la resistencia al PRT en dos líneas, la PI 137739 de Irán y PI 262660 de Bulgaria (Du Toit, 1987, 1988). Desde entonces, la resistencia ha sido registrada en varios cultivares de T. aestivum y en germoplasma no mejorado en todo el suroeste y el centro de Asia y la región del Medio-Oriente (Nkongolo et al., 1989; Zemetra et al., 1990; Quick et al., 1991; Harvey & Martin, 1990; Smith et al., 1991). Hasta el presente, se han identificado 11 genes que otorgan resistencia al PRT, designados como Dn1-Dn2, dn3, Dn4-Dn9, Dnx y Dny (Liu et al., 2001; Smith, 2004, Shufran & Payton, 2009; Beyer et al., 2011).
Los estudios sobre los mecanismos de resistencia implicados en las líneas de trigo, han revelado que la antibiosis, antixenosis y/o la tolerancia están involucradas. La resistencia identificada en las primeras fuentes se atribuyó principalmente a la antibiosis y antixenosis (Du Toit, 1987, 1989; Smith et al., 1992), aunque luego se identificaron algunos casos con tolerancia (Du Toit, 1989; Nkongolo et al., 1989).
La Universidad de Colorado (USA) ha desarrollado para el mercado variedades de trigo resistentes al PRT como Halt, Prairie Red, Prowers 99 y Yuma, entre otras (Kiplagat, 2005). Todas ellas con el gen de resistencia Dn4, que demostró un buen grado de tolerancia, ya que el PRT puede sobrevivir en densidades poblacionales similares a las observadas en las variedades susceptibles, pero no produce ni enrollamiento ni clorosis foliar en forma de estrías (Kiplagat, 2005). En la actualidad, en los EE.UU. y Sudáfrica, son sembradas anualmente grandes extensiones con variedades de trigo resistentes al PRT, sin embargo, los cultivares resistentes portadores del gen Dn4 demostraron ser susceptibles a un nuevo biotipo de PRT, designado como biotipo 2 (Voothuluru et al., 2006).
Dada la inminente aparición de biotipos del PRT en distintos países del mundo y a la permanente expansión del áfido, el objetivo del presente trabajo fue evaluar el comportamiento de distintas líneas de trigo y cultivares diferenciales portadores de genes de resistencia a D. noxia, con el propósito de establecer la composición biotípica del áfido y de identificar aquellos hospederos con resistencia genética al PRT.
MATERIAL Y MÉTODOS
Población de Diuraphis noxia
Se utilizó una mezcla de seis poblaciones del PRT provenientes de distintas regiones del país, colectadas sobre hospederos cultivados y silvestres, como puede observarse en la Tabla I. Las poblaciones se multiplicaron y mantuvieron aisladas en trigo (variedad susceptible Buck Poncho), sobre un sustrato compuesto por 3 partes de tierra y 1 de arena. En cada recipiente se colocó una mezcla de pulgones adultos, provenientes de las distintas localidades. El material vegetal se renovó cuando el 50% de las plantas mostraron síntomas de clorosis, y se aislaron con un envase plástico transparente, con malla de red fina (voile) para la ventilación (cobertor). Esta protección permitió, además, evitar las contaminaciones con otros áfidos que afectan a las gramíneas y el efecto de los parasitoides que pudieran diezmar las poblaciones del PRT. Las poblaciones fueron mantenidas en el insectario de la Cátedra de Terapéutica Vegetal, Fac. Cs. Agrs. y Forestales, bajo condiciones ambientales controladas (20 °C ± 2 y 16:8 hs de fotoperíodo).
Tabla I. Colecciones de poblaciones de Argentina de D. noxia, utilizadas para evaluar la composición biotípica del áfido.

Material vegetal
Se utilizaron cultivares diferenciales de trigo, caracterizados por ser portadores de los distintos genes de la serie "Dn", que otorgan resistencia a los biotipos designados en EE.UU. y de fuentes de resistencia de las que se desconocen dichos genes, denominadas "Líneas Novel" (Tabla II). Dichos materiales, provistos por el Dr. Lage del CIMMYT (Centro Internacional de Mejora de Maíz y Trigo de México), se sembraron el 15 de Julio de 2007 en un invernáculo de la Cátedra de Genética (Facultad de Ciencias Agrarias y Forestales, UNLP), bajo condiciones ambientales no controladas. A fin de evitar contaminaciones con otras especies de áfidos y de proteger el material de los enemigos naturales, se cubrió la totalidad del ensayo con una malla de red fina (organza), tanto en los laterales como el techo de la superficie cultivada.
Tabla II: Diferenciales de trigo utilizados para la determinación de la composición biotípica de D. noxia en Argentina. Letras iguales en la misma columna no difieren estadísticamente (a=0,05).

La siembra se realizó siguiendo un diseño completamente aleatorio con 2 repeticiones, incluyendo 18 cultivares y fuentes de resistencia. Cada línea (diferenciales y noveles) se sembró a chorrillo, en una parcela de 30 x 30 cm, con aproximadamente 70 plantas. Se empleó como testigo susceptible el cultivar Seri y como testigos resistentes a Synthetic y CItr2401.
Al mes de realizada la siembra, cuando las plantas de trigo tenían dos a tres hojas verdaderas, se infestaron con 10 pulgones adultos ápteros por planta. Se realizaron observaciones semanales, agregando insectos en caso de ser necesario, en aquellas parcelas en las que se observó una baja densidad poblacional.
La primera observación se realizó a los 21 días de la infestación (Tabla II), se evaluó el nivel de daño y se asignó un puntaje en grados de 0 a 5, donde 0 correspondió a planta sana y 5 a planta muerta. En la evaluación del daño, se consideró el nivel de estriado, el vigor y la altura de la planta. Además, se evaluó el enrollamiento foliar, considerando si la hoja se encontraba enrollada (E) o plana (P). En todos los casos, se evaluó planta a planta y se obtuvo un promedio por parcela. La segunda evaluación se realizó a los 51 días de iniciada la infestación y se registraron los mismos parámetros que en la primera (Tabla II).
Análisis estadístico
Los resultados obtenidos fueron analizados mediante ANOVA con el PROC GLM (SAS 1998) y el Test de Tukey, para determinar las diferencias entre medias (a=0,05). Los supuestos del ANOVA se verificaron a través de la prueba de normalidad de Shapiro Wilks modificado, y el supuesto de homocedasticidad por la prueba de Levene, con el software estadístico InfoStat (Di Rienzo et al., 2008).
RESULTADOS
Primera lectura
De la totalidad de los cultivares y fuentes de resistencia evaluadas, a los 21 días de iniciada la infestación, se observaron diferencias altamente significativas en el nivel de daño producido por el PRT (F: 549,16; gl: 17; 162; p< 0,0001) (Tabla II).
Los dos testigos resistentes (CItr2401 y Synthetic) tuvieron un buen comportamiento en la primera lectura, con un grado de daño foliar menor a 1,5 y hoja plana, sin embargo, varias líneas presentaron un mayor grado de tolerancia frente a la injuria de D. noxia (Tabla II). Dentro de estas últimas, la línea portadora del gen Dn2 (que presentó hoja enrollada, Tabla II) y la mayoría de las líneas Novel del CIMMYT (14, 2, 1, 15, 17 y 19) registraron un nivel de daño menor a 1,5 y hoja plana en la primera observación (Tabla II).
Entre los cultivares que presentaron un comportamiento de tolerancia intermedia (con un daño entre 1,5 y 3,5), se ubicaron el cultivar portador del gen Dn7 (Diferencial 7), junto a la línea CIMMYT 20, seguida por el testigo susceptible Seri, la línea Diferencial 3 (portadora del gen dn3), Diferencial 1 (con el gen Dn1) y Diferencial 5 (con el gen Dn5) (Tabla II). En estas tres últimas líneas, se observó enrollamiento foliar. Por otro lado, dentro de los materiales más afectados, se ubicaron la línea CIMMYT 33 y por último la línea Diferencial 4 (gen Dn4); este cultivar fue el más dañado a los 21 días de iniciada la infestación (Tabla II).
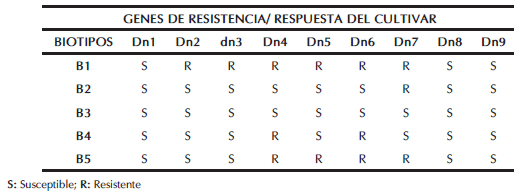
Al comparar el nivel de daño de los cultivares diferenciales estudiados, la línea Diferencial 2 (gen Dn2) fue la que presentó mejor comportamiento, seguida por la Diferencial 7 (gen Dn7) (daño foliar de 1 y 2, respectivamente). En base a estos resultados, podría inferirse que, dado que Dn2 solo es resistente al Biotipo 1 (B1) y que Dn7 también es resistente a B2 y B5 (Tabla III), los biotipos presentes en las poblaciones Argentinas serían el B1, B2* y B5*. Al B2 lo denominamos B2* porque la línea Diferencial 2 (Dn2) no mostró susceptibilidad a ese biotipo local (Tabla II). De igual modo, al B5 se lo identifica como B5*, puesto que el gen Dn2 no otorga resistencia a ese biotipo (Tabla III), sin embargo bajo infestación con las poblaciones Argentinas, dicho gen mantuvo resistencia (Tabla II). Esto vuelve a corroborarse al observar que el cultivar diferencial con peor comportamiento fue el portador del gen Dn4 (susceptible al B2, resistente al B5; Tabla III). Por tal motivo, en las poblaciones Argentinas del PRT, existiría el Biotipo 2* y el Biotipo 5*. Estos tendrían características diferentes al B2 y B5, encontrados en EE.UU., dada la reacción de los cultivares diferenciales portadores de Dn1 (susceptible a todos los biotipos de EE.UU., que frente a las poblaciones locales de PRT mantuvo mayor resistencia que Dn4 y Dn5), Dn2, Dn4 y Dn7 (Tabla III). Por otro lado, el testigo Seri, a pesar de su condición de susceptible a todos los biotipos de EE.UU., presentó un nivel de daño intermedio, lo que confirma las diferentes características de los biotipos B2* y B5* de las poblaciones locales.
Tabla III: Respuesta de los distintos genes de resistencia a los 5 biotipos identificados en EE.UU. según Puterka et al., 2007.
Segunda lectura
A los 51 días de la infestación, los genotipos evaluados presentaron diferencias altamente significativas (F: 1452,01; gl: 17; 162; p<0,0001). Los menores niveles de daño (< 1,5) se observaron en las líneas Novel de CIMMYT 14 y 1, que mantuvieron un comportamiento similar a la primera lectura (Tabla II).
Con un comportamiento intermedio en el nivel de daño (entre 1,5 y 3,5), se ubicaron CItr2401, Synthetic y la línea portadora de Dn7 y CIMMYT 2; todas con hoja plana (Tabla II). Finalmente, dentro de las líneas que presentaron mayores niveles de injuria producida por el PRT, se encontraron los cultivares diferenciales portadores de los genes de resistencia Dn1, Dn3, Dn4 y Dn5 que resultaron muertas y dentro de las líneas Novel, las CIMMYT 15 y 33 (Tabla II). Dentro de los diferenciales, se destaca la línea portadora del gen Dn2. La misma en la primera lectura tuvo un buen comportamiento (a pesar de tener las hojas enrolladas), en la segunda lectura presentó un daño elevado (de 4,2 Tabla II). Como puede observarse en la Tabla II, el resto de las líneas estudiadas resultaron muertas.
El único cultivar diferencial que en la segunda lectura evidenció un comportamiento de tolerancia intermedia fue la línea portadora del gen Dn7 (que otorga resistencia a B2 y a B5). Por tal motivo, podría inferirse que en nuestras poblaciones de PRT estarían presentes el B2* y el B5* (biotipos locales). Los únicos materiales que mantuvieron resistencia a las poblaciones Argentinas del PRT fueron los controles resistentes (CItr2401 y Synthetic) y las líneas Noveles de CIMMYT 14, 1 y 2.
Estos datos indican la presencia de nuevos biotipos en nuestras poblaciones, dado que ninguna de las otras fuentes Dn mantuvo un comportamiento de resistencia estable.
DISCUSIÓN
Los estudios realizados en EE.UU. por Weiland et. al. (2008), quienes evaluaron la virulencia de cinco biotipos del PRT (B1, B2, B6, B7 y B8), encontraron que los diferenciales de trigo portadores del gen Dn7 y el cultivar CITr2401 fueron los que demostraron poseer resistencia a todos los biotipos evaluados. A estas conclusiones arribaron a través del grado de enrollamiento y de clorosis foliar a las tres/cuatro semanas de la infestación. Dado el comportamiento que evidenciaron en este estudio dichos diferenciales, que presentaron una tolerancia media, se estaría en condiciones de confirmar la presencia de, por lo menos, dos biotipos diferentes a los hallados en el país del norte.
A nivel mundial, Smith et al. (2004) determinaron la presencia del B2 en la República Checa, Chile y Etiopía, cuya aparición fue simultánea a la de los EE.UU. Esta aparición global del B2 no podría explicarse fácilmente (Shufran & Payton, 2009). El hecho que ocurriera una nueva introducción de PRT, como se intentó justificar la aparición del nuevo biotipo en EE.UU., no es muy veraz. Se planteó que el B2 ingresó junto a la población original a comienzos de 1980, y por su baja frecuencia en la población no fue detectado. A partir del año 1996, momento en el cual comenzó a cultivarse el trigo "Halt" (portador del gen Dn4), la presión de selección de los nuevos trigos mejorados, cultivados en gran escala, permitió el incremento poblacional de ese biotipo y su detección en el año 2003. En Argentina, el único intento privado de incluir el gen Dn4 a trigos comerciales no llegó a la producción, dado que las líneas portadoras experimentales resultaron susceptibles (Castro, obs. pers.).
Se encuentra ampliamente aceptado que la presión de selección de los cultivares resistentes, constituye un desencadenante en la aparición de los biotipos (Burd et al., 2006). Esto es posible en países como EE.UU. y Sudáfrica, en donde los materiales de trigo mejorados con genes de resistencia, se encuentran ampliamente difundidos, sembrándose anualmente millones de hectáreas. No es el caso de Argentina, donde se cultivan variedades de trigo en los que, en su proceso de mejora, no se han incorporado los genes de la serie Dn. Por tal motivo, deben existir otros factores desencadenantes de la aparición de biotipos.
Porter et al. (1997) realizaron una cronología entre la aparición de los biotipos y el comienzo de la utilización de cultivares comerciales resistentes al áfido, y encontraron que no existiría correlación entre ambos. Estudios más recientes afirman que las razas adaptadas al huésped, probablementeevolucionaron sobre gramíneas silvestres y luego invadieron los cereales cultivados. Por lo tanto, los hospederos no cultivados juegan un rol importante en el origen y mantenimiento de la diversidad de los biotipos (Anstead et al., 2003).
Si se lo compara con la evolución de los biotipos en el "pulgón verde de los cereales" Schizaphis graminum Rond., Burd & Porter (2006) realizaron un relevamiento de los biotipos del áfido en los EE.UU., y encontraron cinco de los nueve biotipos iniciales y 13 biotipos nuevos. Sorprendentemente, los más virulentos fueron los colectados sobre gramíneas silvestres. Los autores sugieren que para el desarrollo de cultivares resistentes, en nuevos planes de mejora, no se tenga en cuenta el biotipo dominante sino los biotipos locales.
Estos conceptos son aplicables a las poblaciones empleadas en este trabajo, principalmente aquellas colectadas en Chos Malal, Junín de los Andes (Neuquén) y Junín (Mendoza), que no corresponden a zonas productoras de cereales y en donde el PRT se encuentra sobre hospederos silvestres todo el año, con climas sumamente rigurosos. Si bien, la población Chos Malal fue colectada en cebada, la misma se obtuvo en una pequeña parcela de no más de una ha, en una región que dista mucho de ser la zona típica productora de trigo o cebada de la Pampa húmeda argentina, donde se siembran cientos de miles de hectáreas de cereales.
CONCLUSIONES
A partir de los resultados obtenidos, se concluye que la interacción del ambiente con el áfido origina poblaciones del PRT que son características de cada región. Por tal motivo, los cultivares mejorados en otros países, que utilizan la resistencia genética como estrategia de manejo del áfido, pueden comportarse como susceptibles ante las poblaciones argentinas. Si bien, algunas de la líneas Novel desarrolladas por el CIMMYT podrían ser incorporadas en planes de mejora del cultivo de trigo en Argentina, los cultivares deberían ser desarrollados en nuestras condiciones de cultivo y con las poblaciones nativas de D. noxia.
BIBLIOGRAFÍA CITADA
1. ANSTEAD, J. A., J. D. BURD & K. A. SHUFRAN. 2003. Mitochondrial DNA sequence divergence among Schizaphis graminum (Hemiptera: Aphididae) clones from cultivated and non-cultivated hosts: haplotype and host associations. Bull. Entomol. Res. 92: 17-24. [ Links ]
2. BEYER, B. M., S. D. HALEY, N. L. V. LAPITAN, J. H. PENG & F. B. PEAIRS. 2011. Inheritance of Russian wheat aphid resistance from tetraploid wheat accessions during transfer to hexaploid wheat. Euphytica. 179: 247-255. [ Links ]
3. BURD, J. D., D. R. PORTER, G. J. PUTERKA, S. D. HALEY & F. B. PEAIRS. 2006. Biotypic variation among North American Russian wheat aphid (Homoptera: Aphididae) populations. J. Econ. Entomol. 99: 1862-1866. [ Links ]
4. CLÚA, A. A., A. M. CASTRO, S. RAMOS, D. O. GIMÉNEZ, A. VASICEK, H. O. CHIDICHIMO & A. F. G. DIXON. 2004. The biological characteristcs and distribution of the greenbug Schizaphis graminum and Russian wheat aphid Diuraphis noxia (Hem: Aphididae) in Argentina and Chile. Eur. J. Entomol. 101: 193-198. [ Links ]
5. DI RIENZO, J. A., F. CASANOVES, M. G. BALZARINI, L. GONZALEZ, M. TABLADA & C. W. ROBLEDO. 2008. InfoStat, versión 2008, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
6. DU TOIT, F. & H. A VAN NIEKERK. 1985. Resistance in Triticum species to the Russian wheat aphid, Diuraphis noxia (Mordvilko) (Hemiptera: Aphididae). Cereal Res. Commun. 13 : 371-378. [ Links ]
7. DU TOIT, F. 1987. Resistance in wheat (Triticum aestivum) to Diuraphis noxia (Hemiptera: Aphididae). Cereal Res. Commun. 15: 175-179. [ Links ]
8. DU TOIT, F. 1988. Another source of Russian wheat aphid (Diuraphis noxia) resistance in Triticum aestivum. Cereal Res. Commun.16: 105-106. [ Links ]
9. DU TOIT, F. 1989. Inheritance of resistance in two Triticum aestivum lines to Russian wheat aphid (Homoptera: Aphididae). J. Econ. Entomol. 82: 1779-1781. [ Links ]
10. HARVEY, T. L. & T. J. MARTIN. 1990. Resistance to Russian wheat aphid, Diuraphis noxia, in wheat, (Triticum aestivum). Cereal Res. Commun. 18: 127-129. [ Links ]
11. HUGHES, R. D. & G. F. MAYWALD. 1990. Forecasting the favourableness of the Australian environment for the Russian wheat aphid, Diuraphis noxia (Homoptera: Aphididae), and its potential impact on Australian wheat yields. Bull. Entomol. Res. 80: 165-175. [ Links ]
12. HUGHES, R. D. 1990. Russian wheat aphid: Will Australia be next?. In: "Aphid plant interactions: Population to Molecules", Proceedings sponsored by the Division of Agriculture, Oklahoma State University and Agricultural Research Service, USDA. Stillwater, Oklahoma, USA. August 12-17, 1990. pp. 329. [ Links ]
13. JONES, J., W.J. R. BYERS, R. A. BUTTS & J. L. HARRIS. 1989. A new pest in Canada: Russian Wheat Aphids Diuraphis noxia (Mordvilko) (Homoptera: Aphididae). Can. Entomol. 121 (7): 623-624. [ Links ]
14. KAZEMI, H. M., P. TALEBI-CHAICHI, M. R. SHAKIBA & M. MASHHADI JAFARLOO. 2001. Biological responses of Russian wheat aphid, Diuraphis noxia (Mordvilko) (Homoptera: Aphididae) to different wheat varieties. J. Agr. Sci. Tech. 3: 249-255. [ Links ]
15. KIPLAGAT, O. K. 2005. The Russian wheat aphid (Diuraphis noxia Mord.): Damage on Kenyan wheat (Triticum aestivum L.) varieties and possible control through resistance breeding. Thesis Wageningen University. Available in: http://edepot.wur.nl/121633, 147 pp. [ Links ]
16. LIÚ, X. M., C. M. SMITH, B. S. GILL & V. TOLMAY. 2001. Microsatellite markers linked to six Russian wheat aphid resistance genes in wheat. TAG. 102: 504-510. [ Links ]
17. NKONGOLO, K. K., J. S. QUICK, A. E. LIMIN, D. B. FOWLER, F. B. PEAIRS & W. L. MEYER. 1990. Russian Wheat Aphid (Diuraphis noxia) resistance in wheat and related species. Can. Plant Sci. 70 (3): 691-698. [ Links ]
18. NKONGOLO, K. K., J. S. QUICK, W. L. MAYER & F. B. PEAIRS. 1989. Russian wheat aphid resistance of wheat, rye, and triticale in greenhouse tests. Cereal Res. Commun. 17: 227-232. [ Links ]
19. NORAMBUENA, M. H. & P. GERDING. 1991. Actual distribución en Chile del áfido ruso del trigo Diuraphis noxia Mordvilko (Homoptera: Aphididae). Agricultura Técnica (Chile) 51 (1): 65-68. [ Links ]
20. ORTEGO, J. & M. A. DELFINO.1992. Presencia de Diuraphis noxia (Mordvilko) (Homoptera: Aphididae) en la Argentina. En: VIII Jornadas Fitosanitarias Argentinas, Paraná, Septiembre 1992. pp. 56. [ Links ]
21. PORTER, D. R., J. D. BURD, K. A. SHUFRAN, J. A. WEBSTER & G. L. TEETES. 1997. Greenbug (Homoptera: Aphididae) biotypes: selected by resistant cultivars or preadapted opportunists? J. Econ. Entomol. 90: 1055-1065. [ Links ]
22. PORTER, D. R., J. A. WEBSTER & C. A. BAKER. 1993. Detection of resistance to Russian wheat aphid in hexaploid wheat. Plant Breed. 110: 157-160. [ Links ]
23. PUTERKA, G. J., J. D. BURD, D. PORTER, K. SHUFRAN, C. BAKER, B. BOWLING & C. PATRICK. 2007. Distribution and diversity of Russian Wheat Aphid (Hemiptera: Aphididae) biotypes in North America. J. Econ. Entomol. 100 (5):1679-1684. [ Links ]
24. QUICK, J. S., K. K. NKONGOLO, W. L. MEYER, F. B. PEARS & B. WEAVER. 1991. Russian Wheat Aphid Reaction and Agronomic and Quality Traits of a Resistant Wheat. Crop. Sci. 31:50-53. [ Links ]
25. SAS, Institute. 1987. SAS/STAT guide for personal computers, version 6.03 Carry NC. [ Links ]
26. SHUFRAN, K. & L. PAYTON. 2009. Limited genetic variation within and between Russian Wheat Aphid (Hemiptera: Aphididae) Biotypes in the United States J. Econ. Entomol. 102 (1): 440-445. [ Links ]
27. SMITH, C. M., D. J. SCHOTZKO, R. S. ZEMETRA & E. J. SOUZA. 1992. Categories of Resistance in Wheat Plant Introductions Resistant to the Russian Wheat Aphid (Homoptera: Aphididae) Resistance in Wheat. J. Encon.Entomol. 85: 1480-1484. [ Links ]
28. SMITH, C. M., D. J. SCHOTZKO, R. S. ZEMETRA, E. J. SOUZA & S. SCHROEDER-TEETER. 1991. Identification of Russian Wheat Aphid (Homoptera: Aphididae) Resistance in Wheat. J. Encon. Entomol. 84: 328-332. [ Links ]
29. SMITH, C. M., T. BELAY, C. STAUFFER, P. STARY, I. KUBECKOVA & S. STARKEY. 2004. Identification of Russian wheat aphid (Homoptera: Aphididae) biotypes virulent to the Dn4 resistance gene. J. Econ. Entomol. 97: 1112-1117. [ Links ]
30. STARÝ, P. 1996. The expansive Russian wheat aphid, Diuraphis noxia (Mord.) detected in the Czech Republic. Anz. Schädlingskd. Pfl. Umwelt. 69: 19-20. [ Links ]
31. STARÝ, P., Z. BASKY, L. K. TANIGOSHI & Z. TOMANOVICĆ. 2003. Distribution and history of Russian wheat aphid, Diuraphis noxia (Kurdj.) in the Carpatian Basin (Hom, Aphididae). J. Pest Sci. 76: 17-21. [ Links ]
32. VOOTHULURU, P., J. MENG, C. KHAJURIA, J. L. LIECENG ZHU, S. STARKEY, J. E. WILDE, C. BAKER & M. SMITH. 2006. Categories and Inheritance of Resistance to Russian Wheat Aphid (Homoptera: Aphididae) Biotype 2 in a Selection from Wheat Cereal Introduction 2401. J. Econ. Entomol. 99 (5): 1854-1861. [ Links ]
33. WEBSTER, J. A., C. A. BAKER & D. R. PORTER. 1991. Detection and mechanisms of Russian Wheat Aphid (Homoptera: Aphididae) resistance in barley. J. Econ. Entomol. 84 (2): 669-673. [ Links ]
34. WEILAND, A., F. PEAIRS, T. RANDOLPH, J. RUDOLPH, S. HALEY & G. PUTERKA. 2008. Biotypic Diversity in Colorado Russian Wheat Aphid (Hemiptera: Aphididae) Populations. J. Econ. Entomol. 101 (2): 569-574. [ Links ]
35. ZEMETRA, R. S., D. SCHOTZKO, C. M. SMITH & E. J. SOUZA. 1990. Seedling resistance to the Russian wheat aphid in wheat germoplasm. Cereal Res. Commun. 18: 223-227. [ Links ]
36. ZERENE, M. Z., M. D. GAGLEVIC & I. A. RAMÍREZ. 1987. Un nuevo áfido de los cereales detectado en Chile. Agric. Téc. 48: 60-61. [ Links ]

