










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.71 no.3-4 Mendoza dic. 2012
TRABAJOS CIENTÍFICOS
Redescripción y consideraciones biogeográfcas de dos especies de Scotobius (Coleoptera: Tenebrionidae) de ambientes montanos del centro de Argentina y Chile
Redescription and biogeographic considerations of two species of Scotobius (Coleoptera: Tenebrionidae) from mountain environments of central Argentina and Chile
Silvestro, Violeta A.*, Rodolfo Carrara** y Gustavo E. Flores**
* Laboratorio de Entomología, Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina; e-mail: ayelensilvestro@hotmail.com
** CONICET, Laboratorio de Entomología, Instituto Argentino de Investigaciones de las Zonas Áridas (IADIZA, CCT CONICET Mendoza), Casilla de correo 507, 5500 Mendoza, Argentina; e-mail: rcarrara@mendoza-conicet.gov.ar; gfores@mendoza-conicet.gov.ar
Recibido: 8-IV-2012;
Aceptado: 12-VI-2012
RESUMEN. El género neotropical Scotobius Germar (Tenebrioninae: Scotobiini) comprende 61 especies distribuidas desde el centro de Perú y sur de Brasil hasta el sur de Argentina y Chile. En este trabajo, revisamos dos especies de Scotobius que habitan el oeste de las provincias de San Juan, Mendoza y Neuquén (Argentina) y en Chile central, a lo largo de la Cordillera de los Andes y las montañas extra-andinas en Argentina: S. punctatus Eschscholtz, 1831 y S. andrassyi Kaszab, 1969. Se proveen redescripciones utilizando nuevos caracteres morfológicos y se proporcionan fotografías de los adultos y de los pronotos. Se consigna la distribución geográfica, rangos altitudinales y las provincias biogeográficas en las que habitan estas especies. Se utiliza un modelo predictivo de distribución de especies para proponer hipótesis sobre los factores que influencian la distribución espacial y que explicarían la alopatría de ambas especies.
PALABRAS CLAVE. Scotobius; Tenebrionidae; Distribución; Biogeografía; Cordillera de los Andes.
ABSTRACT. The Neotropical genus Scotobius Germar (Tenebrioninae: Scotobiini) comprises 61 species distributed from central Peru and southern Brazil to southern Argentine and Chile. In this contribution two species of Scotobius: S. punctatus Eschscholtz, 1831 and S. andrassyi Kaszab, 1969, that inhabit in western San Juan, Mendoza, and Neuquén provinces (Argentina) and in central Chile, along the "Cordillera de los Andes" and extra-Andean mountains in Argentina, are revised. Redescriptions using new morphological data and photographs of habitus and pronota are provided. This article informs about the geographic distribution, altitudinal ranges and the biogeographic provinces that these species inhabit. A predictive model of species distribution is presented to propose hypothesis about the factors that influence the space distribution and the allopatry of these two species.
KEY WORDS. Scotobius; Tenebrionidae; Distribution; Biogeography; Cordillera de los Andes.
INTRODUCCIÓN
Scotobius Germar pertenece a Scotobiini, una tribu de Tenebrioninae creada por Solier (1838), cuyas especies son conspicuos habitantes de ambientes áridos y semiáridos de América del Sur (Doyen, 1993). Esto constituye una excepción respecto a especies de otras tribus dentro de la subfamilia Tenebrioninae que habitan ambientes mésicos y húmedos en todo el mundo (Watt, 1974). Las especies de Scotobiini presentan adaptaciones etológicas, morfológicas y fisiológicas, que les permiten regular el balance hídrico y así sobrevivir a la aridez. Las principales son: hábitos nocturnos, la pérdida del segundo par de alas, la unión íntima entre los élitros a lo largo de la sutura elitral y con los esternitos abdominales, y el desarrollo de una cavidad subelitral que ayuda a reducir la pérdida de agua por los espiráculos (Cloudsley-Thompson, 2001). De acuerdo a estas características y a la clasificación propuesta por Sánchez Piñero & Aalbu (2002), respecto a las adaptaciones a la aridez, los podemos considerar como tenebriónidos altamente adaptados para soportar las condiciones desérticas.
Dentro de Scotobiini, el género con mayor diversidad es Scotobius de las que se conocen 61 especies; todas ellas son ápteras, habitan ambientes áridos y semiáridos desde el sur de Perú y Brasil hasta el sur de Argentina y Chile y se encuentran desde el nivel del mar hasta los 4.000 m de altitud (Kulzer, 1955). La última revisión de Scotobius fue realizada por Kulzer (1955) y algunas de sus especies se describieron sobre la base de pocos caracteres, que a veces no son muy claros de apreciar. Eso ha llevado a errores en la identificación.
Como resultado de viajes de colecta y muestreos sistemáticos, desarrollados en los meses de diciembre y febrero entre los años 2004 y 2006, en la provincia de Mendoza (Argentina) por investigadores del Laboratorio de Entomología del Instituto Argentino de Investigaciones de las Zonas Áridas (IADIZA), se obtuvieron ejemplares de dos morfoespecies de Scotobius, que solo pudieron ser identificados al estudiar los tipos de S. andrassyi Kaszab, 1969 del HNHM y el material determinado por Hans Gebien y Hans Kulzer de S. punctatus Eschscholtz, 1831 del NHMB. Ambas especies habían sido descriptas para Chile (Eschscholtz, 1831; Kaszab, 1969).
Los objetivos del presente trabajo son: (1) realizar un estudio taxonómico de dos especies de Scotobius que han permanecido confundidas o mal identificadas en colecciones: S. punctatus y S. andrassyi, empleando nuevos caracteres que permiten una clara identificación; y (2) consignar su distribución geográfica con nuevas localidades y las provincias biogeográficas que habitan, conjuntamente con algunas consideraciones biogeográficas, aplicando modelos predictivos de distribución.
MATERiAL Y MÉTODOS
Se estudiaron ejemplares de Scotobius depositados en las siguientes colecciones:
HNHM Hungarian Natural History Museum, Budapest, Hungría (Ottó Merkl),
IADIZA Instituto Argentino de Investigaciones de las Zonas Áridas, Mendoza, Argentina (Sergio Roig Juñent),
JEBC Colección privada Juan Enrique Barriga, Curicó, Chile (Juan E. Barriga),
MNNC Museo Nacional de Historia Natural, Santiago, Chile (Mario Elgueta),
NHMB Natural History Museum, Basilea, Suiza (Eva Sprecher).
Algunos tipos de las especies que describió Eschscholtz (1829, 1831) de Tenebrionidae de América del Sur Austral están depositados en el Museum fur Naturkunde der Humboldt Universitat, Berlín, Alemania (Flores, 2007). Sin embargo, al solicitar el tipo de Scotobius punctatus nos fue comunicado que no se encuentra allí (B. Jaeguer, com. pers.). Kulzer (1955) afirmó que no pudo examinar el tipo de S. punctatus, pero estudió ejemplares determinados por H. Gebien. Kaszab (1969) estableció pocas diferencias entre ambas especies, para lo cual debió disponer de ejemplares bien determinados de S. punctatus. En el presente estudio, se examinan ejemplares de S. punctatus de la ex colección H. Gebien en el NHMB -que también fueron estudiados por Kulzer (1955)-y el holotipo de S. andrassyi del HNHM.
La terminología para la morfología externa en la redescripción de las especies es la misma empleada por Kulzer (1955) y se añaden nuevos caracteres del pronoto y élitros que han sido usados recientemente para separar y reconocer especies (Flores, 2009). Se tomaron fotografías digitales de los especímenes con una cámara Canon S50 adaptada a un microscopio estereoscópico Leica MZ6. Las imágenes obtenidas (Figs. 1-4) fueron montadas con un programa de superposición combinada ZM (Hadley, 2006). La información del material tipo se cita en etiquetas separadas (entre corchetes).

Figs. 1-4. Adultos en vista dorsal y pronotos de Scotobius punctatus (1-2) y S. andrassyi (3-4). Escala = 2 mm.
Para la determinación y caracterización de las áreas biogeográficas se utilizaron los trabajos de Cabrera (1971), Gajardo (1994), Martínez Carretero (1995), Roig & Martínez Carretero (1998) y Roig-Juñent et al. (2003).
Área de Estudio
Abarca las provincias de San Juan, Mendoza y Neuquén en Argentina y las regiones de Valparaíso, Santiago, Libertador General Bernardo O'Higgins, Maule, Biobío y Araucanía en Chile (Fig. 5). La zona se caracteriza por la presencia de dos cordones montañosos paralelos con dirección N-S: la Cordillera de los Andes y, al este de esta, la Precordillera. La Cordillera se extiende por el margen pacífico de América del Sur, actúa como límite geográfico entre Argentina y Chile, y alcanza en estas regiones alturas que superan los 6.000 metros, decrece en la provincia de Neuquén y las regiones de Biobío y Araucanía. La Precordillera se extiende desde La Rioja hasta el sur del Río Mendoza en Argentina (desde los 28ºhasta los 33ºde latitud sur), ocupa unos 450 km al oeste de Argentina (Roig & Martínez Carretero, 1998); su ancho variable no supera los 100 km y alcanza altitudes máximas entre 3.000 m en Paramillos de Uspallata (Mendoza) hasta 4.900 m en Sierra de Punilla (La Rioja) (Roig Juñent et al., 2003).

Fig. 5. Distribución geográfica de Scotobius punctatus (círculos) y S. andrassyi (estrellas).
Biogeográficamente, el área de estudio pertenece al dominio Andino-Patagónico que incluye la Cordillera de los Andes y se ensancha al sur, abarcando la Patagonia. Su clima es frío y seco, con heladas casi todo el año y nevadas en invierno. La vegetación dominante es la estepa arbustiva o herbácea con formas extremas de adaptación al frío y a la sequía. El dominio se divide en tres provincias: Altoandina, Puneña y Patagónica, las que a su vez se dividen en distritos y estos en formaciones (Cabrera, 1971); se caracterizan aquí solo aquellos que abarca este trabajo.
Altoandina. Corresponde a las altas montañas de la Cordillera de los Andes, en la zona central de Chile y Argentina, entre los 2.200 y 4.500 m. El clima es frío y seco, con precipitaciones en forma de granizo o nieve y una temperatura media inferior a 8ºC. La vegetación es escasa, formada por estepas graminosas o de caméfitos en cojín; predominan especies de Stipa spp., Festuca spp. y Poa spp.; además de Adesmia pinnifolia, Ephedra andina, Berberis empetrifolia, Senecio uspallatensis, Adesmia subterranea, Menonvillea cuneata y Tropaeolum polyphyllum (Cabrera, 1971).
Puna. Ocupa la Precordillera de Mendoza y San Juan en Argentina, entre los 2.700 y 3.200 m (Roig & Martínez Carretero, 1998). La temperatura media oscila entre los 7,9 y 9,9ºC y presenta escasas precipitaciones (Martínez Carretero, 1995). El tipo de vegetación dominante son los pastizales de altura y estepas arbustivas áridas, arbustos áfilos o con hojas reducidas, plantas resinosas y en general formas altamente xerófilas (Cabrera, 1971). Presenta también amplias zonas de suelo desnudo que reciben irradiación solar directa. Las especies vegetales características son Plazia daphnoides, Lycium decipiens, L. fuscum, Ephedra multiflora, Verbena diversifolia, Baccharis thymifolia, B. polifolia y B. tola (Roig & Martínez Carretero, 1998).
Pastizales de altura. En la vertiente oriental de la Precordillera, entre los 2.300 hasta los 2.700 m aparecen los pastizales con vegetación propia de las Sierras, como Adesmia uspallatensis, A. horrida, Mulinum spinosum, Verbena asparagoides, Senecio uspallatensis, Ephedra andina, Chuquiraga ruscifolia, Stipa paramilloensis y coironales de Stipa tenuissima. Este ambiente posee una marcada estacionalidad, que condiciona en muchos casos los procesos de migración, por lo que su fauna está estrechamente relacionada con la de los dos ambientes que la rodean: la Puna y el Monte (Roig Juñent et al., 2003).
Payunia. Pertenece a la provincia biogeográfica de la Patagonia (Cabrera, 1971), abarca desde el centro de la provincia de Mendoza hasta el centro de Neuquén en Argentina, con precipitaciones entre 200 y 400 mm anuales (Paruelo et al., 1998) y una temperatura media de 13,4ºC (Cabrera, 1971). En ella dominan las estepas de Nassauvia axillaris, Haplopappus pectinatus, Maihuenia patagonica y Stillingia patagonica y se encuentran muchas plantas endémicas como Prosopis castellanosii, P. ruiz leali, Cassia arnottiana, C. kurtzii, Condalia megacarpa y Berberis comberi (Roig, 1998).
Estepa patagónica con Araucaria. Es una formación vegetal que caracteriza la cuenca del curso superior del río Biobío en la región de Araucanía, Chile. Con clima frío de altura y uno a dos meses de período seco (Mardones et al., 2002). Predomina la estepa de Festuca scabriuscula, Poa obvalata, Quinchamalium chilense y Adesmia emarginata con bosques de Araucaria araucana en los lugares más altos de la vertiente oriental de los Andes (Gajardo, 1994). Sobre los 1.500 metros sobre el nivel del mar, las precipitaciones suelen ser sólidas y las temperaturas bajo 0°C (Mardones et al., 2002).
Modelos predictivos de distribución de especies
Los modelos de distribución de especies son usados, en general, para predecir una distribución potencial, relacionando las localidades conocidas de las especies con una serie de variables ambientales (Guisan & Thuillier, 2005). Estos modelos generan predicciones espaciales de hábitats apropiados que indican dónde podrían estar distribuidas las especies analizadas. En este estudio, se aplica la metodología descripta en Roig-Juñent et al. (2008) para realizar los modelos predictivos de distribución. Las localidades conocidas de Scotobius punctatus y S. andrassyi fueron georreferenciadas, mapeadas y usadas para modelar su distribución potencial. El modelo de distribución de especies se corrió usando datos climáticos actuales. Se utilizó el programa MaxEnt (Phillips et al., 2006), combinado con 19 variables climáticas obtenidas de la base de datos WorldClim (Hijmans et al., 2005) para modelar la distribución de S. punctatus y S. andrassyi. La resolución de las capas ambientales usadas fue de 4,6 x 4,6 km, aproximadamente.
Finalmente, para representar las distribuciones potenciales de ambas especies se traducen los resultados predichos por MaxEnt, a un mapa de presencias y ausencias de acuerdo al criterio de mínima presencia. Este criterio define como umbral de corte la igualdad del valor mínimo de predicción del modelo, a un dato de presencia observada (i.e., valores por debajo del valor mínimo de predicción fueron transformados a cero) lo que permite un modelo final sin omisiones (Thomé et al., 2010).
RESULTADOS
Scotobius punctatus Eschscholtz, 1831 (Figs. 1-2)
Scotobius punctatus Eschscholtz, 1831: 11, lámina 18, Fig. 5; Solier, 1838: 65 [como sinónimo de Scotobius armentarius Lacordaire, 1830: 283]; Laporte, 1840: 198; Lacordaire, 1859: 129 (restaurado); Gemminger & Harold, 1870: 1857 (cat.); Philippi, 1887: 725 (cat.); Gebien, 1910: 185 (cat); Gebien 1937: 800; Blackwelder, 1945: 520; Kulzer, 1955: 429; Peña 1966: 422; Vidal & Guerrero, 2007: 294 (caracterización y fotografía como S. andrassyi).
Diagnosis. Pronoto con puntos iguales o más grandes que los del élitro (Fig. 2), muy densos, distribuidos de manera uniforme, separados por una distancia igual o menor al diámetro de un punto; ángulos anteriores agudos, proyectados hacia adelante y ángulos posteriores rectos.
Redescripción. Largo 13-17 mm (Fig. 1). Coloración negra a rojiza. Cabeza. Labro con borde anterior cóncavo y pelos dorados; clípeo y frente con abundantes puntos grandes, de igual tamaño, con una pequeña seta en cada alveolo, distribuidas regularmente. Antenas cortas, llegan a la mitad del pronoto; segundo segmento más corto que los restantes, tercer segmento más largo que los restantes, del cuarto al séptimo más largos que anchos y del octavo al onceavo más anchos que largos. Tórax. Pronoto cóncavo, más ancho que largo (Fig. 2), con puntos iguales o más grandes que los del élitro, muy densos, distribuidas uniformemente en todo el pronoto, separados por una distancia igual o menor al diámetro de un punto; ángulos anteriores agudos, proyectados hacia adelante (Fig. 2), ángulos posteriores rectos; cuartos laterales y bordes laterales planos y dirigidos hacia abajo, mitad anterior del margen lateral más baja que el disco, cuarto posterior del margen lateral del mismo nivel que el disco. élitros (Fig. 1) con nueve hileras longitudinales de grandes puntos, las dos laterales externas en el pseudopleuron; entre las hileras de puntos hay costillas insinuadas anchas, con pequeños puntos dispersos; las costillas derivan posteriormente en protuberancias y convergen postero-lateralmente; sutura elitral no elevada, epipleuron con reborde irregular y del mismo ancho en toda su longitud. Abdomen con puntos dispersos. Patas. Fémures y tibias con puntos; tibias posteriores rectas en la base.
Variación intraespecífica. En el carácter ancho máximo del pronoto encontramos variaciones fenotípicas: en ejemplares de Chile, se observa en el medio del pronoto y, en ejemplares de Argentina (Mendoza y Neuquén), en el tercio posterior (Fig. 2).
Material examinado: ARGENTINA: Mendoza: Dto. Tupungato: Pampa Guaitallari, 23-X-1998, S. Roig, 1 IADIZA; Dto. San Carlos: Reserva Laguna Del Diamante, Alvarado, 2328 m, I-2006, S. Roig-G. Debandi, 9 IADIZA, 2352 m, XII-2004, G. Debandi-A. Marvaldi, 6 IADIZA, 2539 m, 18-II-2005, G. Debandi, 1 IADIZA; Dto. San Rafael: Volcán Diamante, 2233 m, 26-XII-2005 al 10-I-2006, S. Roig-G. Debandi, 10 IADIZA; 19 km E dique Agua del Toro, 18-XII-1998, G. Flores-S. Roig, 1 IADIZA; 20 km N dique Agua del Toro, 18-XII-1998, G. Flores-S. Roig, 1 IADIZA; Dto Malargüe: Puesto Las Gateadas, Arroyo el Rosario, 12-X-1974, A. Roig, 10 IADIZA; 7 km S seccional guardaparques La Payunia, 1.728 m, 11-II-2007, G. Flores, 1 IADIZA; 58 km N Las Loicas, 1.847 m, 3-II-2007, G. Flores, 1 IADIZA; 43 km N Las Loicas, 1.742 m, 3-II-2007, G. Flores, 1 IADIZA; 32 km SW Las Loicas, 2.257 m, 5-II-2007, G. Flores, 1 IADIZA; Sierra de Palauco, 2.659 m, 6-II-2007, G. Flores, 1 IADIZA; Sierra de Palauco, Laguna el Piojo, 2.329 m, 30-I-2010, R. Carrara, F. Campón, E. Scheibler, 1 IADIZA; El Nevado, 2.344m, II-2005, G. Debandi-E. Ruiz, 9 IADIZA, 2.592 m, I-2006, S. Roig-G. Debandi, 17 IADIZA, 2.909 m, XII-2004, G. Debandi-A. Marvaldi, 3 IADIZA, 3.114 m, XII-2004, G. Debandi-A. Marvaldi, 3 IADIZA. Neuquén: Dto. Norquin: Chenque-Pehuén, 1.850 m, 9-I-1967, M. Gentili, 3 IADIZA; Copahue, 12-II-1951, P. R. San Martín, 1 IADIZA; Dto. Picunches: Primeros Pinos, 1.615 m, 11-II-1971, M. Gentili, 4 IADIZA. CHILE: sin datos [Museo Nacional Santiago] 2 MNNC, 1 NHMB, Lonquimay [Museo Nacional Santiago] 1 MNNC (todos determinados por H. Kulzer en 1955).
Distribución (Fig. 5): En Argentina: sur y oeste de Mendoza y oeste de Neuquén, y en Chile al este de la Región de la Araucanía. En los distritos y formaciones Altoandina central, Pastizal de altura, Payunia y Estepa patagónica con Araucaria; desde los 1.600 m en la estepa patagónica hasta los 2.300 m en la Cordillera de los Andes y en los sistemas extra-andinos El Nevado y Sierra de Palauco, hasta los 3.150 y 2.700 m respectivamente.
Scotobius andrassyi Kaszab, 1969 (Figs. 3-4)
Scotobius andrassyi Kaszab, 1969: 314; Elgueta, 1988: 129 (biog.).
Diagnosis. Puntos del pronoto más pequeños que los puntos del élitro (Fig. 4), dispersos en el disco, separados por una distancia igual o mayor al diámetro de dos puntos y más densos en los cuartos laterales; ángulos anteriores rectos u obtusos, no proyectados hacia adelante y ángulos posteriores obtusos.
Redescripción. Largo 12-19 mm (Fig. 3). Coloración negra a rojiza, opaca. Cabeza. Labro con borde anterior cóncavo y pelos dorados; clípeo y frente con abundantes puntos grandes de igual tamaño, con una pequeña seta en cada alveolo, distribuidas regularmente. Antenas cortas que llegan a la mitad del pronoto; segundo segmento más corto que los restantes, tercer segmento más largo que los restantes, del cuarto al séptimo más largos que anchos y del octavo al onceavo más anchos que largos. Tórax. Pronoto más ancho que largo (Fig. 4); ancho máximo en el medio; puntos del pronoto más pequeños que los puntos del élitro, dispersos en el disco, separados en el disco por una distancia igual o mayor al diámetro de dos puntos, en los cuartos laterales más densos, separados por una distancia igual o menor al diámetro de un punto; ángulos anteriores rectos u obtusos, no proyectados hacia adelante, ángulos posteriores obtusos (Fig. 4); cuartos laterales y bordes laterales aplanados y dirigidos hacia abajo, muy por debajo del nivel del disco, mitad anterior del margen lateral de menor nivel que el disco; cuarto posterior del margen lateral del mismo nivel que el disco. élitros (Fig. 3) con nueve hileras longitudinales de grandes puntos, las dos laterales externas en el pseudopleuron; entre las hileras de puntos hay costillas insinuadas anchas con pequeños puntos dispersos o sin ellos; las costillas derivan posteriormente en protuberancias y convergen postero-lateralmente; sutura elitral no elevada; epipleuron con reborde irregular y del mismo ancho en toda su longitud. Abdomen con puntos dispersos. Patas. Fémures y tibias con puntos; tibias posteriores rectos en la base.
Variación intraespecífica. En individuos de Paramillos de Uspallata, se encontraron variaciones fenotípicas: puntos del disco del pronoto densos, distribuidos uniformemente, separados por una distancia mayor al diámetro de un punto y menor al diámetro de dos puntos, bordes laterales dirigidos hacia arriba, muy poco por debajo del nivel del disco.
Material tipo. Holotipo: [Hungarian Soil-Zool. Exp./ Chile: Prov. Santiago/ Baños de Morales/ 30-X-1965] [Nr. P-B 74/ leg. Andrássy, Balogh/ Loksa et Mahunka] [Holotypus 1969/ Scotobius/ andrassyi/ Kaszab] (HNHM). Un paratipo: [Anden b. Santiago/ Chile, leg. Franz] [Paratypus 1969/ Scotobius/ andrassyi/ Kaszab] (HNHM).
Otro material examinado.
ARGENTINA: Mendoza: Dto. Las Heras: Punta de Vacas, 2.442 m, I-2006, S. Claver-A. Scollo, 1 IADIZA; Los Penitentes, 2.700 m, 12-III-1995, G. Flores, 4 IADIZA; 1 km E Puente del Inca, 2.650 m, 15-XII-2001, G. Flores, 13 IADIZA; Puente del Inca, 2.700 m, 18-I-1976, A. Roig, 2 IADIZA, 16-II-1977, A. Roig, 2 IADIZA, 28-XII-1981, A. Roig, 2 IADIZA, 20-I-1984, A. Roig, 4 IADIZA, 15-XII-2001, G. Flores, 3 IADIZA, 2-XI-2003, G. Flores, 6 IADIZA; Parque Provincial Aconcagua, 2.907 m, II-2006, S. Claver-A. Scollo, 2 IADIZA, Laguna Horcones, 2.900 m, 28-XII-1981, A. Roig, 1 IADIZA, 16-XII-2001, G. Flores, 1 IADIZA; entre Puente del Inca y Las Cuevas, 3.100 m, 28-XII-1981, A. Roig, 1 IADIZA; Las Cuevas, 3.200 m, 28-XII-1981, A. Roig, 3 IADIZA, 16-XII-2001, G. Flores, 2 IADIZA; 14 km NW Estancia Tambillos, Uspallata, 2.700 m, 13-I-1999, G. Flores, 1 IADIZA; Paramillos de Uspallata, 2.850 m, II-2006, S. Claver-A. Scollo, 9 IADIZA, 26-I-2008, G. San Blas, F. Aballay, E. Ruiz, 3 IADIZA; 3 km E Paramillos de Uspallata, 2.830 m, 1-XI-2001, S. Roig-G. Flores, 3 IADIZA; Pampa de Canota, 2.890 m, I-2006, S. Claver-A. Scollo, 3 IADIZA. San Juan: Dto. Ullúm: Reserva Don Carmelo, 3.090 m, XI-2005, N. Borruel, 1 IADIZA. CHiLE: Región de Valparaíso: Río Blanco, 11-I-1918, 1 MNNC; Piscicultura, 1.500 m, 11-XII-1958, L.E. Peña, 1 NHMB; entre Juncal y Portillo, 2.550 m, 20-I-1984, A. Roig, 4 IADIZA. Región Metropolitana de Santiago: Baños de Colina, 18-IV-1947, L.E. Peña, 1 NHMB, 2-IV-1959, L.E. Peña, 3 NHMB (todos determinados por H. Kulzer en 1955 y 1959 como S. punctatus), 14-III-1983, M. Elgueta, 3 MNNC; Baños de Morales 2.000 m, III-1984, J.E. Barriga, 1 JEBC; Farellones, 3.200 m, IV-1988, J.E. Barriga, 1 JEBC; Lo Valdés, 12-III-1948, L.E. Peña, 1 NHMB, 13-II-1955, 1 MNNC, Lo Valdés, 3.000 m, 16-XII-1986, J.E. Barriga, 1 JEBC; Lagunillas, 10-X-1969, J. Moroni, 1 MNNC, 10-XII-1982, G. Arriagada, 1 MNNC, 30-XI-1983, G. Arriagada , 1 MNNC. Región del Libertador General Bernardo O'Higgins: Caletones, 16-XII-1982, M. Elgueta, 1 MNNC.
Distribución: En Argentina: noroeste de Mendoza y centro-suroeste de San Juan, en Chile: este de las regiones de Valparaíso, Santiago y Libertador General Bernardo O'Higgins. En los distritos Altoandina central y Puna, en la Cordillera de los Andes, entre los 1.500 y los 3.200 m en el lado chileno de la Cordillera; entre los 2.440 y los 3.200 m en el lado argentino de la Cordillera y en el sistema extra-andino Precordillera, entre los 2.800 y los 3.100 m.
Modelos predictivos de distribución de especies
Teniendo en cuenta los resultados de los modelos predictivos de distribución (Figs. 6 a y b), se puede observar que S. punctatus y S. andrassyi comparten áreas con características climáticas viables para su subsistencia, por lo que eventualmente podrían ser simpátricos (i.e., en el centro de nuestra área de estudio, a lo largo de la Cordillera de los Andes). Este hallazgo contrasta con nuestras observaciones, ya que estas especies nunca han sido colectadas en simpatría en la misma localidad; más aún, considerando que la zona límite de distribución entre ambas especies representa una de las áreas mejor prospectadas en los viajes de colecta y muestreos sistemáticos.
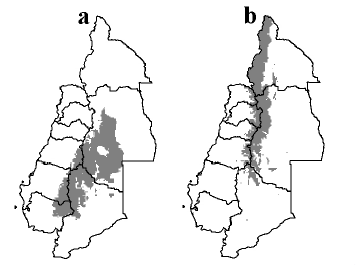
Fig. 6. Distribución predictiva de: a, Scotobius punctatus; b, S. andrassyi. De acuerdo al criterio de mínima presencia el umbral de corte fue de 41% para S. andrassyi y de 45% para S. punctatus.
DISCUSIÓN
La redescripción de Scotobius andrassyi y S. punctatus, con el empleo de nuevos caracteres realizado en este trabajo, contribuye sustancialmente a clarificar la identidad de estas dos especies. Esta clase de estudios taxonómicos constituye una verdadera herramienta para evitar confusiones cuando las especies son similares en su morfología externa, y sirve de aporte para otras disciplinas como la Ecología, donde conocer la identidad de las especies es la base para el desarrollo de las investigaciones.
Considerando el área de distribución de Scotobius punctatus y S. andrassyi, ambas habitan a ambos lados de la Cordillera de los Andes. Esto resulta llamativo, porque ha sido reportado que para otras especies del mismo género la Cordillera de los Andes actuó como una barrera, originando endemismos de especies de Scotobius a ambos lados; en el centro de Chile (Coquimbo a Biobío) (e.g., S. bullatus Curtis, S. kirbyi Solier, S. gayi Solier, S. asperatus Erichson, S. rugosulus Guérin-Méneville y S. emarginicollis Kulzer) y en el centro-oeste de Argentina (San Juan a Neuquén) (e.g., S. casicus Lacordaire, S. wittmeri Kulzer, S. alaticollis Kulzer y S. caraboides Fairmaire) (Kulzer, 1955). Teniendo en cuenta estas evidencias, se puede afirmar que el levantamiento de los Andes no ha sido una barrera que afectó la distribución de S. punctatus y S. andrassyi, como sí lo fue para otros tenebriónidos ápteros (Flores & Pizarro-Araya, 2006). Esto sugiere que para estas dos especies, al poder habitar ambientes de altura, la Cordillera de los Andes no funcionó como barrera sino como corredor.
La distribución observada de S. punctatus y S. andrassyi nos muestra que estas especies no son simpátricas. Específicamente S. punctatus vive en la estepa patagónica y ambientes montanos, tanto en hábitats andinos como extra-andinos. En la región central y austral de su distribución se encuentra en los Andes del lado chileno y argentino, en cambio, al norte de su distribución solo se encuentra en ambientes al pie de los Andes del lado argentino. Los ambientes extra-andinos que habita se encuentran en la Argentina (e.g. El Nevado, Sierra de Palauco) y pertenecen a formaciones volcánicas de la Payunia. La distribución de S. punctatus coincide con lo reportado por Flores & Carrara (2006) para otras especies de tenebriónidos, lo que concuerda con el patrón observado: que los ambientes de altura de la Payunia son viables para especies que habitan la estepa patagónica (Roig-Juñent et al., 2008).
En cambio, la distribución de S. andrassyi aparece restringida solo a ambientes montanos, no se distribuye en la estepa patagónica. Esta especie está asociada a los hábitats de altura de la Cordillera de los Andes, tanto en Chile como en Argentina a una misma latitud; y a los del sistema extra-andino de la Precordillera en Argentina. Este patrón de distribución contrasta con lo hallado por Roig-Juñent et al. (2003), quienes encontraron una escasa relación entre los géneros de artrópodos epigeos que habitan la Precordillera y los que habitan la Cordillera de los Andes; y con lo reportado por Flores & Gómez (2005) quienes encontraron que distintas especies de un mismo género de Tenebrionidae habitan la Precordillera y la Cordillera de los Andes.
Considerando los patrones de distribución de estas especies, surge la pregunta: ¿cuál o cuáles factores pueden ser los responsables de esta alopatría? Se puede hipotetizar acerca de las causas por las cuales S. andrassyi y S. punctatus ven restringida su distribución, se considera que estas no corresponden a un efecto de competencia interespecífica, ya que diversas especies del género Scotobius viven en simpatría (Kulzer, 1955), pero sí que estas podrían deberse a accidentes geográficos que limitan su dispersión en el área. Es probable que una de las causas esté relacionada con la acción erosiva de diversos glaciares (a lo largo del paralelo 32°), que durante el último máximo glacial han ocasionado profundos valles en forma de "U" de más de 100 km de longitud (Kuhle, 2004) y que pueden detener la dispersión de especies ápteras como es el caso de S. andrassyi y S. punctatus (e.g., el Río Mendoza presenta paredes de aproximadamente 30 metros de alto). Apoyando esta última idea, ha sido reportado por Flores & Gómez (2005) que el Río Mendoza, en la misma área donde habita S. andrassyi, ha interrumpido el flujo génico en poblaciones de una especie áptera de tenebriónido a ambos lados del río, y que esta interrupción ha llevado a diferenciaciones en distintos caracteres corporales.
AGRADECIMIENTOS
A los curadores de las colecciones examinadas (Eva Sprecher, Ottó Merkl, Mario Elgueta D. y Sergio Roig-Juñent), por el préstamo del material estudiado; a Remedios Marín, por su ayuda en la confección de las láminas; a la familia Zapata, por permitirnos colectar en la Estancia Tambillos (Uspallata); a la Dirección de Recursos Naturales Renovables de Mendoza, por el permiso para colectar en la provincia de Mendoza y en sus áreas protegidas; y a la Reserva Villavicencio, por el permiso de colecta; a los revisores, por sus comentarios que ayudaron a mejorar el manuscrito. Este trabajo fue financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, Argentina, PIP 112-200801-00162 y PIP 112-201101-00987) y por el Fondo para la Investigación Científica y Tecnológica de Argentina (FONCYT, PICT 01-11120 "Diversidad de artrópodos en ambientes montanos del centro-oeste argentino"). RC y GEF son miembros de la Carrera del Investigador Científico y Tecnológico (CONICET).
BIBLIOGRAFÍA CITADA
1. BLACKWELDER, R. E. 1945. Checklist of the coleopterous insects of Mexico, Central America, the West Indies, and South America. Part 3. Bulletin of the United States National Museum 185: 343-550. [ Links ]
2. CABRERA, A. L. 1971. Fitogeografía de la República Argentina. Boletín de la Sociedad Argentina de Botánica XIV (1-2): 1-42. [ Links ]
3. CLOUDSLEY-THOMPSON, J. 2001. Thermal and water relations of desert beetles. Naturwissenschaften 88: 447-460. [ Links ]
4. DOYEN, J. T. 1993. Cladistic relationship among Pimeliinae Tenebrionidae (Coleoptera) Journal of the New York Entomological Society 101 (4): 443-514. [ Links ]
5. ELGUETA, M. 1988. Insectos epigeos de ambientes altomontanos en Chile central: algunas consideraciones biogeográficas con especial referencia a Tenebrionidae y Curculionidae (Coleoptera). Boletín del Museo Nacional de Historia Natural de Chile 41: 125-144. [ Links ]
6. ESCHSCHOLTZ, F. 1829. Zoologischer Atlas, enthaltend Abbildungen und Beschreibungen neuer Thierarten, während des Flottcapitains von Kotzebue zweiter Reise um die Welt, auf der Russisch- Kaiserlichen Kriegsschlupp Predpriaetië in den Jahren 1823-1826. Drittes Heft. G. Reimer, Berlin. 18 pp. [ Links ]
7. ESCHSCHOLTZ, F. 1831. Zoologischer Atlas, enthaltend Abbildungen und Beschreibungen neuer Thierarten, während des Flottcapitains von Kotzebue zweiter Reise um die Welt, auf der Russisch- Kaiserlichen Kriegsschlupp Predpriaetië in den Jahren 1823-1826. Viertes Heft. G. Reimer, Berlin. 19 pp. [ Links ]
8. FLORES, G. E. 2007. New synonymies and lectotype designations in the Neotropical tribes Nycteliini, Praocini, and Scotobiini (Coleoptera: Tenebrionidae). Annales Zoologici 57 (3): 409-426. [ Links ]
9. FLORES, G. E. 2009. Revision of some types of the South American tribes Nycteliini, Praocini, and Scotobiini (Coleoptera: Tenebrionidae), with new synonymies. Zootaxa 1985: 21-33. [ Links ]
10. FLORES, G. E. & R. CARRARA. 2006. Two new species of Nyctelia Latreille (Coleoptera: Tenebrionidae) from Western Argentina, with zoogeographical and ecological remarks on the high mountain habitat. Annales Zoologici 56 (3): 487-495. [ Links ]
11. FLORES, G. E. & R. S. GóMEZ. 2005. Taxonomía y biogeografía de cuatro especies de Psectrascelis (Coleoptera: Tenebrionidae) de la Precordillera y Cordillera de los Andes en Mendoza, Argentina. Revista de la Sociedad Entomológica Argentina 64 (3): 93-106. [ Links ]
12. FLORES, G. E. & J. PIZARRO-ARAYA. 2006. The Andes mountain range uplift as a vicariant event in the Pimeliinae (Coleoptera: Tenebrionidae) in southern South America. Cahiers scientifiques 10: 95-102. [ Links ]
13. GAJARDO, R. 1994. La vegetación natural de Chile. Clasificación y distribución geográfica. Editorial Universitaria, Santiago, 165 pp. [ Links ]
14. GEBIEN, H. 1910. Tenebrionidae II. En: Junk, W. & S. Schenkling (eds) Coleopterorum Catalogus XVII. Pars 22. W. Junk, Berlin, 167-354. [ Links ]
15. GEBIEN, H. 1937. Katalog der Tenebrioniden (Col. Heteromera). Teil I. Publicazioni del Museo Entomologico Pietro Rossi 2: 505-883. [ Links ]
16. GEMMINGER, M. & E. HAROLD. 1870. Catalogus Coleopterorum hujusque descriptorum synonymiscus et sistematicus. S. Gummi, Monachii, Vol. 7, pp. 1801-2180. [ Links ]
17. GUISAN, A. & W. THUILLIER. 2005. Predicting species distribution: offering more than simple habitat models. Ecology Letters 8: 993-1009. [ Links ]
18. HADLEY, A. 2006. CombineZM software de procesamiento de imagen de dominio público. Disponible en: http://www.hadleyweb.pwp.blueyonder.co.uk/CZM/ combinezm.htm (descargado el 11/Junio/2009). [ Links ]
19. HIJMANS, R. J., S. E. CAMERON, J. L. PARRA, P. G. JONES & A. JARVIS. 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25:1965-1978. [ Links ]
20. KASZAB, Z. 1969. The scientific results of the Hungarian soil Zoological Expeditions to South America. 17. Tenebrioniden aus Chile (Coleoptera). Opuscula Zoologica Budapest 9 (2): 291-337. [ Links ]
21. KUHLE, M. 2004. The Last Glacial Maximum (LGM) glacier cover of the Aconcagua group and adjacent massifs in the Mendoza Andes (South America). En: Ehlers, J. & P. L. Gibbard (eds.), Quaternary Glaciation- Extent and Chronology. Part III: South America, Asia, Africa, Australia, Antarctica, Development in Quaternary Science, vol. 2c. Elsevier B.V., Amsterdam, pp. 75-81. [ Links ]
22. KULZER, H. 1955. Monographie der Scotobiini (Zehnter Beitrag zur Kenntnis der Tenebrioniden). Entomologische Arbeiten aus dem Museum Georg Frey 6: 383-478. [ Links ]
23. LACORDAIRE, J. T. 1830. Mémoire sur les habitudes des insectes coléoptères de l'Amérique méridionale (Hétéromères). Annales de Sciences Naturelles 20: 274-291. [ Links ]
24. LACORDAIRE, J. T. 1859. Histoire naturelle des insectes. Généra des coléoptères ou exposé méthodique et critique de tous les genres proposés jusqu'ici dans cet ordre d'insectes. Tome cinquième. Première partie contenant les familles des Ténébrionides, Cistélides, Nilionides, Pythides, Mélandryides, Lagriides, Pédilides, Anthicides, Pyrochroïdes, Mordellides, Rhipiphorides, Stylopides, Meloïdes et Oedémérides. Librairie Encyclopédique de Roret, Paris. 750pp. [ Links ]
25. LAPORTE, F. L. N. C. (Comte de Castelnau). 1840. Historie naturelle des insectes, vol. 2. P. Duménil, Paris, 564 pp. [ Links ]
26. MARDONES, F., M. MELLADO, M. A. HIDALGO & T. CONNELLY. 2002. Reporte regional, Región de la Araucanía. En: Feliú Silva, X. & J. C. Utreras Díaz (eds.) Informe Regional, Biblioteca del Congreso Nacional, Valparaíso, Chile. [ Links ]
27. MARTíNEZ CARRETERO, E. 1995. La Puna Argentina: Delimitación general y división en distritos florísticos. Boletín de la Sociedad Argentina de Botánica 31 (1-2): 27-40. [ Links ]
28. PEñA L. E. 1966. Catálogo de los Tenebrionidae (Coleoptera) de Chile. Entomologische Arbeiten aus dem Museum Georg Frey 17: 397-453. [ Links ]
29. PHILIPPI, F. H. 1887. Catálogo de los Coleópteros de Chile. Anales de la Universidad de Chile 71: 619-806. [ Links ]
30. PHILLIPS, S. J., R. P. ANDERSON & R. E. SCHAPIRE. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling 190:231-259. [ Links ]
31. PARUELO, J. M., A. BELTRÁN, E. JOBBAGY, O. SALA & R. A. GOLLUSCIO. 1998: The climate of Patagonia: general patterns and controls on biotic processes. Ecología Austral 8: 85-101. [ Links ]
32. ROIG, F. A. 1998. La vegetación de la Patagonia. En: Correa, M.N. (ed.), Flora Patagónica, INTA Colección Científica Tomo VIII, vol. I: 48-174. [ Links ]
33. ROIG, F. A. & E. MARTíNEZ CARRETERO. 1998. La vegetación puneña en la provincia de Mendoza, Argentina. Phytocoenologia 28 (4): 565-608. [ Links ]
34. ROIG-JUñENT, S., G. E. FLORES & C. MATTONI. 2003. Consideraciones biogeográficas de la Precordillera (Argentina), con base en artrópodos epígeos. En: Morrone, J. J. & J. Llorente Bousquets (eds.), Una perspectiva latinoamericana de la Biogeografía, Las Prensas de Ciencias, Facultad de Ciencias, UNAM, México, D.F., pp. 275-288. [ Links ]
35. ROIG-JUñENT, S. A., F. AGRAIN, R. CARRARA, E. RUIZ-MANZANOS & M. F. TOGNELLI. 2008. Description and phylogenetic relationship of two new species of Baripus (Coleoptera: Carabidae: Broscini) and considerations regarding patterns of speciation. Annals of Carnegie Museum 77 (1): 211-227. [ Links ]
36. SÁNCHEZ PIñERO, F. & R. L. AALBU. 2002. The desert specialists: Tenebrionid beetles. En: Case, T. J., M. L. Cody & E. Ezcurra (eds.), A new Island Biogeography of the Sea of Cortes, Chapter 6. Oxford University Press, New York, pp. 129-153. [ Links ]
37. SOLIER, A. J. J. 1838. Essai sur les Collaptèrides. Tribu Scotobites. Annales de la Société Entomologique de France 7: 5-73. [ Links ]
38. THOMÉ, M. T. C., K. R. ZAMUDIO, J. G. R. GIOVANELLI, C. F. B. HADDAD, F. A. BALDISSERRA Jr. & J. ALEXANDRINO. 2010. Phylogeography of endemic toads and post-Pliocen persistente of the Brazilian Atlantic Forest. Molecular Phylogenetics and Evolution 55: 1018-1031. [ Links ]
39. VIDAL, P. & M. GUERRERO. 2007. Los tenebriónidos de Chile. Ediciones Universidad Católica de Chile. Santiago, Chile, 478 pp. [ Links ]
40. WATT, J. C. 1974. A revised subfamily classification of Tenebrionidae (Coleoptera). New Zealand Journal of Zoology 1 (4): 381-452 [ Links ]
