










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680
Rev. Soc. Entomol. Argent. vol.71 no.3-4 Mendoza dic. 2012
TRABAJOS CIENTÍFICOS
Variación temporal de la riqueza, composición y densidad de acridios (Orthoptera: Acridoidea) en diferentes comunidades vegetales del Sur de la provincia de Buenos Aires
Temporal variation in grasshopper (Orthoptera: Acridoidea) richness, composition, and density in grasslands of the southern Buenos Aires province.
Mariottini, Yanina *; María L. De Wysiecki * y Carlos E. Lange *
* Centro de Estudios Parasitológicos y de Vectores (CEPAVE) (CCT La Plata - CONICET - UNLP - CICPBA), Calle 2 nº584, CP (B1902CHX), La Plata, Argentina. ymariottini@cepave.edu.ar; mlw@cepave.edu.ar; carlosl@cepave.edu.ar
Recibido: 31-VIII-2012;
Aceptado:31-X-2012
RESUMEN: Los acridios figuran entre los invertebrados herbívoros dominantes en la mayoría de los sistemas de pastizal. Si bien, en Argentina, la importancia económica de estos insectos ha sido reconocida desde el siglo XIX, aún se desconocen aspectos importantes de la biología y ecología de la mayoría de las especies. El objetivo principal de este estudio fue determinar cómo la riqueza de especies, la composición y la densidad de acridios variaron en las comunidades vegetales más características del Sur de la provincia de Buenos Aires, durante cinco temporadas sucesivas (2005-06 hasta 2009-10), un área históricamente afectada por estos insectos. Se colectó un total de 22especies. La riqueza de acridios fue similar entre las diferentes comunidades vegetales, observándose una disminución significativa de la misma durante la última temporada de muestreo. La densidad total de acridios aumentó significativamente en todas las comunidades vegetales durante el estudio, alcanzando niveles de explosión poblacional ("outbreak") en las últimas dos temporadas. En la de 2009-10, la densidad fue ocho veces mayor que en 2005-06. Las especies que más aportaron a este aumento en la densidad fueron Dichroplus maculipennis (Blanchard) y Borellia bruneri (Rehn). Dichroplus maculipennis resurgió como el acridio plaga más perjudicial en la región Pampeana, con el desarrollo de un "outbreak" de magnitud histórica, se registraron, en algunos sitios, densidades de hasta 75 ind/m2.
PALABRAS CLAVE: Borellia bruneri; Dichroplus maculipennis; Acridios; "Outbreaks"; Pastizales.
ABSTRACT: Grasshoppers are among the dominant invertebrate herbivores in most grassland habitats. The economic importance of grasshoppers has been recognized in Argentina since the XIX century, but many aspects of the biology and ecology of most species remain unknown. The main objective of this study was to determine how grasshopper species richness, composition, and density varied among characteristic plant communities during five successive seasons (2005-06 to 2009-10) in grasslands of the southern Buenos Aires province, an area historically affected by these insects. Twenty two species were collected. Grasshopper species richness was similar between different plant communities and showed a significant decrease in the final year of sampling. The overall density increased significantly in all plant communities during the study, reaching outbreak levels in the last two seasons. Density was over eight times higher in 2009-10 than in 2005-06. The greatest contribution to the increase in density observed was by Dichroplus maculipennis (Blanchard) and Borellia bruneri (Rhen). Dichroplus maculipennis re-emerged as the most serious grasshopper pest in the Pampas, developing an outbreak of historical magnitude that included high densities of up to75 ind/m2.
KEY- WORDS: Borellia bruneri; Dichroplus maculipennis; Grasshoppers; Outbreaks; Grasslands.
INTRODUCCIÓN
Los acridios (tucuras y langostas) figuran entre los insectos herbívoros dominantes en la mayoría de los sistemas de pastizal (Belovsky & Slade, 2000; Zhong-Wei et al., 2006; Branson, 2011). Como consumidores primarios, son importantes en el ciclado de nutrientes y energía en los ecosistemas (Belovsky, 2000). En años de explosiones poblacionales ("outbreaks"), no sólo compiten con el ganado y la fauna silvestre por el forraje, sino que suelen consumir y destruir diversos cultivos (Fielding & Brusven, 1995; Branson et al., 2006, Cigliano et al., en prensa).
Las comunidades de acridios, como otros grupos de insectos, suelen exhibir una gran variabilidad en la composición y abundancia de especies (Branson et al., 2006; Jonas & Joern, 2007), debido a que las poblaciones de las distintas especies tienden a responder en forma diferencial a la interacción de factores externos (condiciones climáticas, vegetación, suelo) e internos (interacciones bióticas) que varían espacial y temporalmente (Branson, 2008).
En Argentina, dependiendo de la región considerada, la importancia económica de los acridios ha sido reconocida desde mediados y fines del siglo XIX (Cigliano et al., en prensa); siendo las explosiones demográficas de las distintas especies un fenómeno recurrente. En la región Pampeana, diferentes especies de acridios han sido una variable importante en la economía agrícola-ganadera, ya que causan considerables daños en pastizales naturales, pasturas implantadas y en diversos cultivos (maíz, soja, girasol, cebada, sorgo, entre otros) (Carbonell et al., 2006; Cigliano et al., 2002, en prensa). Dichroplus maculipennis (Blanchard), Dichroplus elongatus (Giglio-Tos) y Dichroplus pratensis (Bruner) se encuentran entre las especies características (y más importantes desde el punto de vista económico) de las comunidades de acridios de la región Pampeana. Estas suelen exhibir "outbreaks" de importante magnitud, generando significativas pérdidas económicas para los productores agropecuarios (COPR, 1982; Cigliano et al., 2002; Lange et al., 2005; Mariottini et al., 2011a, b; de Wysiecki et al., 2011).
A nivel mundial, existen numerosas contribuciones acerca de la biología, ecología e importancia económica de estos insectos (Capinera, 1987; Joern & Gaines, 1990; Kemp, 1992; Lockwood et al., 2000; Fielding, 2004; Olfert & Weiss, 2006; Branson, 2011, entre otros). Sin embargo en Argentina, la información cuantitativa sobre la estructura de sus comunidades es aún limitada. Salvo por algunos estudios relativamente recientes (Sánchez, 1980; Sánchez & De Wysiecki, 1993; Cigliano et al., 2000, 2002; Torrusio et al., 2002; Mariottini et al., 2011b, en prensa; de Wysiecki et al., 2000, 2004, 2011), el conocimiento acerca de las comunidades de acridios de nuestro país suele ser el resultado de observaciones aisladas y no el reflejo de estudios cuantitativos prolongados en espacio y tiempo.
Teniendo en cuenta lo expuesto, el objetivo principal de este trabajo fue estudiar la variación temporal de la riqueza, composición y densidad de acridios durante cinco temporadas (2005/06-2009/10), en las comunidades vegetales más características del Sur de la provincia de Buenos Aires, un área históricamente afectada por estos insectos (Liebermann, 1972; COPR, 1982).
MATERIALES Y MÉTODOS
Área de estudio:
El área de estudio fue el partido de Laprida (Fig.1), ubicado en el Sur de la región Pampeana (36º02´ S, 59º06´ O). La zona es una llanura que se extiende hasta los 210 m de altitud, la temperatura media es de 22ºC en verano y 6ºC en invierno. La precipitación media anual varía entre los 800 y 900 mm, es un régimen hídrico subhúmedo. Los pastizales constituyen la vegetación predominante (Batista et al., 1988; Chaneton, 2005). Las características de los suelos (baja infiltración, exceso de alcalinidad, napa freática elevada, escasa pendiente y manto de tosca a poca profundidad) han impedido o limitado el uso agrícola continuado de la mayor parte de las tierras, por lo tanto domina la actividad ganadera (Perelman et al., 2001; Batista et al., 2005). Aproximadamente el 45% de la superficie total del partido (345.498 Ha) es utilizada para la cría de ganado ovino (Torrusio & Otero, 2009).

Figura 1: Mapa de Argentina y países vecinos, indicando la región Pampeana (en gris) y el área de estudio (Partido de Laprida).
Muestreo de acridios:
Los sitios de muestreo fueron seleccionados en noviembre de 2005 y se mantuvieron constantes durante las cinco temporadas de estudio (2005-06 a 2009-10). Los mismos representan las comunidades vegetales más comunes de la zona (Batista et al., 1988; Fielding & Brusven, 1993; Torrusio et al., 2002, Mariottini et al., en prensa) y fueron clasificados en cuatro categorías: Pastizales nativos, con mayor cobertura de gramíneas nativas como Stipa formicarum, Panicum bergii, Stipa trichotoma, Festuca pampeana, Stipa caudata, entre otras; Pastizales halófilos, dominados por especies halófilas como Distichlis spicata; Pasturas implantadas, principalmente del género Agropyron y Pastizales disturbados, pastizales nativos sometidos a pastoreo de ganado, con mayor cobertura de dicotiledóneas introducidas como Cardus pycnocephalus, Centaurea calcitrapa y Ammi majus, entre otras (Mariottini et al., en prensa). De cada una de estas categorías se establecieron tres réplicas, lo que dio un total de 12 sitios. Los muestreos se realizaron desde mediados de primavera hasta fines del verano. En promedio, se realizaron 4 muestreos por año.
Los acridios fueron colectados mediante 200 golpes o barridos con redes entomológicas de 40 cm de diámetro y 75 cm de profundidad, a lo largo de cuatro transectas de aproximadamente 3 m de ancho y 50 m de longitud cada una, en los 12 sitios establecidos, de acuerdo a Evans (1984, 1988). Cada golpe consistió en un arco de 180ºa través de la vegetación. Esta metodología es de eficacia probada para la obtención de muestras representativas de una comunidad (Larson et al., 1999). Los acridios colectados se llevaron al laboratorio para su identificación de acuerdo a Carbonell et al. (2006) y Cigliano et al. (en prensa). Para cada fecha y sitio de muestreo, se estimó la riqueza específica, la composición de especies, la abundancia relativa de las mismas, la densidad total de las comunidades y la densidad específica. La densidad de acridios se estimó siguiendo el método desarrollado por Onsager & Henry (1977), que consiste en contar el número de individuos que saltan desde tres series de 10 aros cada una (0,1m2 cada uno), colocados a intervalos de 5 m y a lo largo de tres transectas.
Análisis de los datos:
Los resultados son expresados en valores promedio ± error estándar (ES). La variación en la riqueza de especies, la densidad total de acridios y la densidad específica de las diferentes comunidades vegetales por temporada fueron comparadas realizando un análisis de la varianza de medidas repetidas de una vía. Previo a los análisis, se utilizó la prueba de esfericidad de Mauchly y se realizó el ajuste de los grados de libertad por el método de Greenhouse Geisser, cuando no se cumplió con este supuesto (Scheiner & Gurevitch, 2001). Se utilizó la prueba de Fischer (LSD) para la comparación de medias a posteriori. El programa estadístico utilizado fue Statistica 7.0.
RESULTADOS
Riqueza de especies:
Considerando todo el periodo de estudio, se colectó un total de 22 especies de acridios pertenecientes a las familias Acrididae y Romaleidae (Tabla I). Se registraron 15 especies en los pastizales halófilos, 16 en las pasturas implantadas, 18 en los pastizales disturbados y 20 en los pastizales nativos. No hubo diferencias significativas en la riqueza de acridios de las diferentes comunidades vegetales (Tabla II). Existió una disminución significativa en la riqueza de acridios entre las temporadas de muestreo (factor años, p< 0,001) para todas las comunidades vegetales (Tabla II). En la última temporada, la riqueza de especies fue significativamente menor respecto de las tres primeras temporadas (LSD test p<0,05) (Fig. 2).
Tabla I: Especies de acridios colectadas y densidad de las mismas (promedio ± ES) durante 2005/06-2009/10 en las diferentes comunidades vege Laprida, Buenos Aires.
Tabla II: Reultado del ANOVA de medidas repetidas de una vía para evaluar la riqueza de especies de acridios en las diferentes comunidades vegetales durante 2005/06-2009/10, Laprida, Buenos Aires,


Figura 2: Promedio (± ES) de la riqueza de acridios (S) durante 2005/06-2009/10 en Laprida, Buenos Aires. Letras distintas indican diferencias significativas (LSD test p<0.05).
Densidad y composición de especies:
La densidad de acridios mostró diferencias altamente significativas entre los diferentes años de muestreo (p<0.0001) (Tabla III), observándose un incremento significativo de la misma a lo largo de las cinco temporadas (LSD p<0,001) (Fig. 3). Las cuatro comunidades vegetales estudiadas no mostraron diferencias significativas en la densidad de acridios (p=0,5) (Tabla III). La tabla IV muestra los valores de densidad promedio en las diferentes comunidades vegetales.
Tabla III: Resultado del ANOVA de medidas repetidas de una vía para evaluar la densidad total de acridios en las diferentes comunidades de plantas durante 2005/06-2009/10, Laprida, Buenos Aires, Argentina.
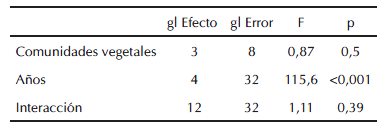

Figura 3: Promedio (± ES) de la densidad de acridios (ind/m2) durante 2005/06-2009/10 en Laprida, Buenos Aires.
Tabla IV: Densidad de acridios (promedio ± ES) en diferentes comunidades vegetales durante 2005/06-2009/10, Laprida, Buenos Aires, Argentina.

Con respecto a la composición y a la densidad específica, se observaron diferencias altamente significativas en todo el periodo de estudio en cada una de las comunidades vegetales (Tabla V).
Tabla V: Resultado del ANOVA de medidas repetidas de un vía para evaluar la densidad específica de acridios en diferentes comunidades vegetales durante 2005/06-2009/10, Laprida, Buenos Aires.

Pastizales Halófilos: En la primer temporada, Covasacris pallidinota (Bruner) fue la especie más abundante (3,83 ± 0,56 ind/m2) (Tabla I), con una densidad significativamente superior al resto de las especies (LSD p <0.05). En la dos temporadas siguientes, C. pallidinota y Borellia bruneri (Rehn) fueron las especies más abundantes (Tabla I) y sus densidades fueron significativamente superiores a las de las restantes (LSD p<0.05). En la cuarta temporada, B. bruneri fue la especie más abundante, con una densidad similar a C. pallidinota (Tabla I) (LSD p >0,05), pero significativamente superior a los valores de densidad registrados para las otras especies (LSD p<0,05). En la quinta temporada, D. maculipennis fue la especie numéricamentedominante con una densidad promedio de 37,74 ± 5,71 ind/m2 y diferencias altamente significativas respecto de las demás (LSD p<0,001).
Pasturas Implantadas: Dichroplus elongatus, B. bruneri, y C. pallidinota fueron las especies más abundantes durante las tres primeras temporadas de muestreo, con densidades similares y superiores a las de las restantes especies, colectadas en esas temporadas (LSD p<0,05). Borellia bruneri fue la más abundante en la cuarta temporada (Tabla I), mientras que, en la quinta temporada, lo fue D. maculipennis (22,12 ± 6,3 ind/m2), (Tabla I).
Pastizales Disturbados: En la primera y segunda temporadas, D. elongatus y B. bruneri fueron las especies más abundantes (Tabla I), mientras que, en la tercera y cuarta temporadas, fueron B. bruneri, D. elongatus, C. pallidinota, y D. maculipennis, con densidades similares y superiores a las de las demás (LSD p<0,05). En la quinta, D. maculipennis fue la especie más abundante (16,71 ± 4,93 ind/m2), con una densidad similar a la registrada para B. bruneri (LSD p>0,05).
Pastizales Nativos: En la primera y segunda temporadas, todas las especies presentaron densidades similares (Tabla I). En la tercera y cuarta, B. bruneri fue la especie más abundante (4,20 ± 0,96 ind/m2 y 10,73 ± 2,36 ind/m2, respectivamente). Durante la quinta temporada, como en las demás comunidades vegetales, D. maculipennis fue la especie más abundante (36.01 ± 2.07 ind/m2).
Teniendo en cuenta las especies más características (B. bruneri, D. maculipennis, C. pallidinota, D. elongatus) de las diferentes comunidades vegetales, se observó que B. bruneri y D. maculipennis mostraron un progresivo y significativo incremento en su densidad a lo largo de los años (LSD p<0,05) (Fig. 4). Contrariamente, C. pallidinota y D. elongatus tuvieron, en algunas comunidades vegetales, una disminución significativa en sus densidades. En particular, en los pastizales halófilos, se registró una disminución significativa en la densidad de C. pallidinota durante la quinta temporada respecto de las anteriores (LSD p <0,05). Dichroplus elongatus disminuyó significativamente su densidad en las pasturas implantadas y los pastizales nativos en la quinta temporada (LSD p <0,05) (Fig. 4). Otras especies menos abundantes que incrementaron su densidad en algunos momentos de muestreo fueron: Dichroplus conspersus (Bruner), Borellia pallida (Rehn), Baeacris pseudopunctulactus (Ronderos) y D. pratensis Bruner (LSD p<0,05). En los pastizales nativos, la densidad de Scotussa lemniscata (Stål), en la cuarta y quinta temporadas, fue menor que durante la primera (LSD p<0,05).

Figura 4: Promedio (± ES) de la densidad (ind/m2) de las especies de acridios más abundantes (Borellia bruneri, Covasacris pallidinota, Dichroplus elongatus y Dichroplus maculipennis) durante 2005/06-2009/10 en Laprida, Buenos Aires.
DISCUSIÓN
En la mayoría de los ecosistemas de pastizales, una riqueza superior de especies de insectos se asocia de manera positiva con una mayor riqueza de plantas y una mayor heterogeneidad en la estructura de la vegetación (Joern, 2005). Considerando las comunidades vegetales del presente estudio, los P. Nativos y los P. Disturbados tienen una mayor riqueza de plantas y también, más heterogeneidad espacial que los P. halófilos y las P. Implantadas (Torrusio et al., 2002, Zhong-Wei et al., 2006, Mariottini et al., en prensa). Ahora bien, los resultados obtenidos en este estudio no coinciden con el patrón antes descripto, ya que el número de especies de acridios colectadas en las diferentes comunidades vegetales fue similar. Al respecto, Kemp et al. (2002) sugieren que la riqueza de plantas no es la variable más adecuada a la hora de diferenciar las comunidades vegetales; ya que no considera la composición específica de las mismas, lo que en definitiva determina la oferta de diferentes recursos alimenticios para los acridios. En este sentido, Schaffers et al. (2008) indican que la composición específica de las plantas en un sitio es una de las variables que mejor predice la riqueza y la abundancia de los grupos de insectos presentes en una determinada comunidad. Recientemente, Branson (2011) señaló que la relación entre la riqueza de plantas y la diversidad y abundancia de acridios en los diferentes sistemas de pastizales aún no es clara.
Teniendo en cuenta la variación temporal de la riqueza de especies, se constató una disminución significativa en la riqueza de acridios durante la última temporada de estudio en todas las comunidades vegetales. Esta situación podría ser consecuencia de que la temporada 2008-09 fue un periodo de una sequía extrema, en la que se registraron las menores precipitaciones de los últimos 47 años (Servicio Meteorológico Nacional 2009). En este sentido, Kemp & Cigliano (1994), en pastizales de Montana (EEUU), colectaron un número de especies significativamente menor en las temporadas siguientes a años de extrema sequía. Es importante destacar que las precipitaciones representan una de las variables climáticas que más afecta la dinámica de las diferentes poblaciones de acridios, modificando la estructura del hábitat y la disponibilidad y calidad nutricional de las plantas de las que se alimentan (Branson & Sword, 2010; de Wysiecki et al., 2011).
A lo largo del período de estudio, se observó un aumento progresivo y significativo de la densidad de acridios en todas las comunidades vegetales; se registró en la cuarta y quinta temporada una explosión demográfica ("outbreak") de gran magnitud; siendo en la última temporada, la densidad total de acridios 8 veces mayor que la registrada al inicio del estudio. Los procesos subyacentes a la aparición de explosiones poblacionales de acridios siguen siendo poco conocidos (Joern, 2000; Branson et al., 2006; Branson, 2008). Sin embargo, Lockwood (1997) proporcionó una definición ecológica sobre los "outbreaks" de acridios en pastizales de América del Norte para abordar problemas tanto científicos como de gestión. Combinando las variables de espacio, tiempo y densidad, dicho autor define tentativamente un "outbreak" como un incremento mayor a 6 ind/m2, durante un período de 1 a 2 años, que ocurre sobre más de un tipo de hábitat. El criterio considerado en la definición se basa en la densidad a partir de la cual las poblaciones exceden la capacidad de carga del recurso. Lamentablemente, en Argentina, la información sobre la densidad de acridios y la capacidad de carga de los diferentes ambientes es extremadamente limitada. Teniendo en cuenta este enfoque, Cigliano et al. (2002) reportaron un "outbreak" de acridios en el partido de Benito Juárez (sudeste de la provincia de Buenos Aires), con una densidad promedio de 27,4 ind/ m2 (7 veces superior a la registrada en los años anteriores) y determinaron períodos de "no outbreak", años de transición y periodos de "outbreak". Los resultados del presente trabajo muestran una dinámica similar, ya que corresponde el periodo 2005-08 a los años de transición y el periodo a partir de fines del año 2008, al "outbreak".
Con respecto a la composición y abundancia de las especies de acridios, se observó que relativamente pocas especies representaron la mayor proporción de la comunidad, independientemente de la densidad total de acridios. Esta característica ha sido observada previamente por otros autores en otras zonas de la región Pampeana, así como también en pastizales de América del Norte (Joern, 1982; Joern & Pruess, 1986; Cigliano et al., 2002; de Wysiecki et al., 2004). La situación observada en este trabajo fue similar a la que describen Joern & Pruess (1986), quienes plantean que, a pesar de las fluctuaciones poblacionales, el rango de especies de una comunidad es relativamente predecible y las especies típicas suelen ser habituales año tras año. De igual manera, Cigliano et al. (2002) registraron que las especies más comunes fueron las mismas tanto en años de "outbreak" como de "no outbreak".
Las especies que más contribuyeron al aumento de la densidad de la comunidad en su conjunto fueron D. maculipennis y B. bruneri. El predominio de estas dos especies constituye la diferencia principal con el "outbreak" registrado por Cigliano et al. (2002), donde las dos especies más abundantes fueron D. elongatus y S. lemniscata. Esta diferencia podría deberse a las condiciones inusualmente secas de la temporada 2008-09, las cuales podrían no ser favorables para el desarrollo de D. elongatus y S.lemniscata, ya que ambas especies prefieren ambientes más húmedos con vegetación densa (Cigliano & Ronderos, 1994; Carbonell et al., 2006).
La explosión poblacional registrada en este estudio puede considerarse como un "outbreak" de dos especies, D. maculipennis y B. bruneri. La importancia de este "outbreak" radica en que representa el resurgimiento de D. maculipennis como acridio perjudicial en la región y también el primer registro de altas densidades de B. bruneri para la misma. Esta última especie es considerada como uno de los acridios plaga más importante de las pampas en Uruguay (COPR, 1982) y fue abundante en zonas de la provincia de Neuquén (Lange & Cigliano, 2010). Históricamente, D. maculipennis ha sido una de las especies de acridios más perjudiciales de Argentina, tanto en la región Pampeana como en zonas del Oeste de la Patagonia (COPR, 1982; Carbonell et al., 2006; Cigliano et al., en prensa). En la primera mitad de los años 80, D. maculipennis representaba aproximadamente un 70% de las comunidades de acridios de la región Pampeana, numéricamente, es la especie más importante en varios partidos del Sur de la provincia de Buenos Aires (Ronderos, 1986). Sin embargo, a partir de la década del 90, distintos estudios realizados en la región advirtieron una importante disminución en su abundancia (Cigliano et al., 2000; De Wysiecki et al., 2000, 2004; Torrusio et al., 2002, Mariottini et al., 2011b). En 1994, Cigliano et al. (1995) encontraron en las comunidades de acridios de los alfalfares del sudeste de Buenos Aires, una abundancia relativa de D. maculipennis menor al 5% y de Wysiecki et al., (2004) en el partido de Benito Juárez, la muestran como una especie de distribución rara, que se encontró solo en el 6% del total de los sitios muestreados. Contrariamente a lo que ocurrió en la región Pampeana, en varias temporadas de la década del 90, ocurrieron fuertes explosiones demográficas de D. maculipennis en valles fértiles del Oeste de las provincias de Neuquén y Chubut, las que ocasionaron importantes pérdidas de forraje (Lange et al., 2005). Cigliano et al. (1995) sugirieron posibles factores responsables de la aparente disminución de D. maculipennis en la región Pampeana, tales como: cambios climáticos, campañas de fumigación masivas, cambios en los patrones de uso de la tierra, e incluso características intrínsecas de la biología de esta especie. Sin embargo, a partir de los muestreos periódicos realizados en este trabajo se pudo constatar que la dinámica poblacional de D. maculipennis experimentó un cambio respecto a su abundancia. En la actualidad, representa la especie dominante en la mayoría de las comunidades de acridios estudiadas. El "outbreak" de D. maculipennis, que comenzó a fines de 2008, fue de una dimensión histórica, con densidades que en algunos sitios alcanzaron los 75 ind/m2, además, se observaron los denominados "vuelos en masa" característicos de la dispersión de los adultos de esta especie, los que pueden alcanzar hasta 40 Km (COPR, 1982), lo que causa substanciales pérdidas económicas para los productores agrícolas y ganaderos de la zona. Aquí, es necesario resaltar la importancia de desarrollar estudios de campo adicionales y experiencias en laboratorio, que permitan identificar los factores que mayor influencia tienen en la variación espacial y temporal de la abundancia de las diferentes especies de acridios.
AGRADECIMIENTOS
Este estudio fue parcialmente financiado por la Agencia Nacional de Promoción Científica y Tecnológica (PICT 2006-679) y la Comisión de Investigaciones Científicas de la provincia de Buenos Aires (CICPBA). C. Lange es Investigador de la CICPBA.
BIBLIOGRAFÍA
1. BATISTA, W. B., R.J.C. LEÓN & S. B. PERELMAN. 1988. Las comunidades vegetales de un pastizal natural de la región de Laprida, Prov. de Buenos Aires, Argentina. Phytoeconología. 16 (4): 519-534. [ Links ]
2. BATISTA, W. B., M. A. TABOADA, R. S. LAVADO, S. B. PERELMAN & R. J. C. LEON. 2005. Asociación entre comunidades vegetales y suelos en el pastizal de la Pampa Deprimida. En: Oesterheld, M., M. R. Aguiar, C. M. Ghersa & J. M. Paruelo (eds.), La heterogeneidad de la vegetación de los agroecosistemas. Un homenaje a Rolando León. Editorial Facultad de Agronomía, Buenos Aires, pp. 113-129. [ Links ]
3. BELOVSKY, G. E. 2000. Do grasshoppers diminish grassland productivity? A new perspective for control based on conservation. En: Lockwood, J. A., A. V. Latchininsky & G. Sergeev (eds.), Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks without Risking Environmental Disaster. Kluwer Academic, Boston, pp. 7-29. [ Links ]
4. BELOVSKY, G. E. & J. B. SLADE. 2000. Insect herbivory accelerates nutrient cycling and increases plant production. Proc. Natl. Acad. Sci. 97 (26): 14412-14417. [ Links ]
5. BRANSON, D. H. 2008. Influence of a large late Summer Precipitation Event on Food Limitation and Grasshopper Population Dynamics in a Northen Great Plains Grassland. Environ. Entomol. 37 (3): 686-695. [ Links ]
6. BRANSON, D. H. 2011. Relationships between Plant Diversity and Grasshoppers Diversity and Abundance in the Little Missouri National Grassland. Psyche. Special issue: 1-7. [ Links ]
7. BRANSON, D. H. & G. A. SWORD. 2010. An experimental analysis of grasshopper community responses to fire and livestock grazing in a northern mixed-grass prairie. Environ. Entomol. 39 (5): 1441-1446. [ Links ]
8. BRANSON, D. H., A. JOERN & G. A. SWORD. 2006. Sustainable Management of Insect Herbivores in Grassland Ecosystems: New Perspectives in Grasshopper Control. Bioscience. 56 (9): 743-755. [ Links ]
9. CAPINERA, J. L. 1987. Population ecology of rangeland grasshoppers. En: Capinera J.L. (ed.), Integrated PestManagement on Rangeland: A shor-tgrass Prairie perspective. Westview, Boulder, Colorado, pp. 162-182. [ Links ]
10. CARBONELL, C., M. M. CIGLIANO & C. E. LANGE. 2006. Especies de Acridomorfos (Orthoptera) de Argentina y Uruguay. CD-ROM. Published by the "Orthopterists Society" and The Museo de la Plata, Argentina. [ Links ]
11. CHANETON, E. J. 2005. Factores que determinan la heterogeneidad de la comunidad vegetal en diferentes escalas espaciales. La heterogeneidad de la vegetación de los agroecosistemas. En: Oesterheld M, M. R. Aguiar, C. M. Ghersa & J. M. Paruelo (eds), La heterogeneidad de la vegetación de los agroecosistemas. Un homenaje a Rolando León. Editorial Facultad de Agronomía, Buenos Aires, pp. 19-42. [ Links ]
12. CIGLIANO, M. M. & R. A. RONDEROS. 1994. Revision of the South American grasshopper genera Leiotettix Bruner and Scotussa Giglio Tos (Orthoptera, Acrididae, Melanoplinae). Trans. Am. Entomol. Soc. 120 (2): 145-180. [ Links ]
13. CIGLIANO, M M., M. L. DE WYSIECKI & C. E. LANGE 1995. Disminución de la abundancia de Dichroplus maculipennis en comunidades del sudoeste de la provincia de Buenos Aires. Rev. Soc. Entomol. Argent. 54 (1-4): 41-42. [ Links ]
14. CIGLIANO, M M., M. L. DE WYSIECKI & C. E. LANGE. 2000. Grasshopper (Orthoptera, Acrididae) species diversity in the pampas, Argentina. Divers. Distrib 6: 81-91. [ Links ]
15. CIGLIANO, M M., S. TORRUSIO & M. L. DE WYSIECKI. 2002. Grasshopper (Orthoptera: Acrididoidea) community composition and temporal variation in The Pampas, Argentina. J. Orthoptera. Res. 11(2): 215-221. [ Links ]
16. CIGLIANO, M M., M. E. POCCO & C. E. LANGE. En prensa. Acridoideos (Orthoptera) de importancia agroeconómica. En: Roig-Juñent S, L. E. Claps & J. J. Morrone (eds.), Biodiversidad de Artrópodos Argentinos, V. 3. Sociedad Entomológica Argentina, Mendoza, pp: 1-26. [ Links ]
17. COPR (Centre for Overseas Pest Research). 1982. The locust and grasshopper agricultural manual, COPR, London, p. 690. [ Links ]
18. DE WYSIECKI, M. L., N. SÁNCHEZ & S. RICCI. 2000. Grassland and shrubland grasshopper community composition in northern La Pampa province, Argentina. J. Orthoptera. Res. 9: 211-221 [ Links ]
19. DE WYSIECKI, M. L., S. TORRUSIO & M. M. CIGLIANO. 2004. Caracterización de las comunidades de acridios del partido de Benito Juárez, sudeste de la provincia de Bs. As, Argentina. Rev. Soc. Entomol. Argent. 63: 87-96. [ Links ]
20. DE WYSIECKI, M. L., M. ARTURI., S. TORRUSIO & M. M. CIGLIANO. 2011. Influence of weather variables and plant communities on grasshopper density in the Southern Pampas, Argentina. J. Insect Sci. Disponible en: http//insectscience.org/11.109 [ Links ]
21. EVANS E. W. 1984. Fire as a natural disturbance to grasshopper assemblages of tallgrass prairie. Oikos. 43: 9-16. [ Links ]
22. EVANS E. W. 1988. Grasshopper (Insecta: Orthoptera: Acrididae) assemblages of tallgrass prairie: influences of fire frequency, topography, and vegetation. Can.J. Zool. 66: 1495-1501. [ Links ]
23. FIELDING, D. J. 2004. Developmental time of Melanoplus sanguinipes (Orthoptera: Acrididae) at high latitudes. Environ. Entomol. 33 (6): 1513-1522. [ Links ]
24. FIELDING, D. J. & M. A. BRUSVEN. 1993. Grasshoppers (Orthoptera: Acrididae) Community composition an ecological disturbance on Southern Idaho rangeland. Environ. Entomol. 22 (1): 71-81. [ Links ]
25. FIELDING, D. J. & M. A. BRUSVEN. 1995. Ecological correlates between rangeland grasshopper (Orthoptera: Acrididae) and plant communities of southern Idaho. Environ. Entomol. 24 (6): 1432-1441. [ Links ]
26. JOERN, A. 1982. Distributions, Densities and Relative Abundances of Grasshoppers (Orthoptera: Acrididae) in a Nebraska Sandhills Prairie. Prairie. Nat. 14: 37-45. [ Links ]
27. JOERN, A. 2000. What are the consequences of non-linear ecological interactions for grasshopper control strategies?. En: J. A. Lockwood, A. V. Latchininsky & M.Sergeev (eds.), Grasshoppers and Grassland Health. Kluwer Academic Publishers. Dordrecht, Netherlands, pp. 131-144. [ Links ]
28. JOERN, A. 2005. Disturbance by Fire and Bison Grazing Modulate Grasshopper Assemblages in Tallgrass Prairie. Ecology. 86: 861-873. [ Links ]
29. JOERN, A. & K. P. PRUESS. 1986. Temporal constancy in grasshopper assemblies (Orthoptera: Acrididae). Ecol. Entomol. 11: 379-385. [ Links ]
30. JOERN, A. & S. B. GAINES. 1990. Population dynamics and regulation in grasshoppers. En: Chapman, R. F. & A. Joern (eds.), Biology of Grasshoppers. John Wiley and Sons, Inc. New York, pp. 415-483. [ Links ]
31. JONAS, J. L. & A. JOERN. 2007. Grasshopper (Orthoptera: Acrididae) communities respond to fire, bison grazing and weather in North American tallgrass prairie: a longterm study. Oecologia. 153: 699-711. [ Links ]
32. KEMP, W. P. 1992. Temporal variation in rangeland grasshopper (Orthoptera: Acrididae) communities in the steppe region of Montana, USA. Can.Entomol. 124: 437-450 [ Links ]
33. KEMP, W.P. & M. M. CIGLIANO. 1994. Drought and rangeland grasshopper species diversity. Can. Entomol. 126: 1075-1092. [ Links ]
34. KEMP, W.P., K. M. O'NEILL., M. M. CIGLIANO & S. TORRUSIO. 2002. Field-scale Variations in Plant and grasshopper Communities: A GIS-based Assessment. Trans. in GIS. 6: 115-133. [ Links ]
35. LANGE, C. E. & M. M. CIGLIANO. 2010. Prevalence and infection intensity of the biocontrol agent Paranosema locustae (Microsporidia) in field-collected, newly-associated hosts (Orthoptera: Acrididae: Melanoplinae). Biocontrol. Sci. Technol. 20: 19-24. [ Links ]
36. LANGE, C. E., M. M. CIGLIANO & M. L. DE WYSIECKI. 2005. Los acridoideos de importancia económica en la Argentina. En: Barrientos Lozano, L. & P. Almaguer. (eds), Manejo integrado de la langosta centroamericana y acridoideos plaga en América Latina. Instituto Tecnológico de Ciudad Victoria, Tamaulipas, México, p. 93-135 [ Links ]
37. LARSON, D. P., K. M. O´NEILL & W. P. KEMP. 1999. Evaluation of the accuracy of sweep sampling in determining grasshopper (Orthoptera: Acridoidea) community composition. J. Agron. Urban. Entomol. 16 (3): 207-214. [ Links ]
38. LIEBERMANN, J.1972. The current state of the locust and grasshopper problem in Argentina. En: Hemming, C.F, & T.H.C. (eds.), Proc. Int. Study Conf. on the Current and Future Problems of Acridology, London, pp.191-198. [ Links ]
39. LOCKWOOD, J. A. 1997. Rangeland Grasshopper Ecology. En: Gangwere, S.K, M. C. Muralirangan & M. Muralirangan (eds.), The Bionomics of Grasshoppers. Katydids and Their Kin. CAB International. Wallingford, UK, pp. 83-101. [ Links ]
40. LOCKWOOD, J. A., A. V. LATCHINISKY & M. G. SERGEEV. 2000. Grasshoppers and Grassland Health. Managing grasshopper Outbreaks without risking environmental disaster. Kluver Academic Publishers. NATO Sciences series. 2. Environmental Security. [ Links ]
41. MARIOTTINI, Y., M. L. DE WYSIECKI & C. E. LANGE. 2011a. Postembryonic development and consumption of the melanoplines Dichroplus elongatus Giglio-Tos and Dichroplus maculipennis (Blanchard) (Orhtoptera: Acrididae: Melanoplinae) under laboratory conditions. Neotrop. Entomol. 40: 190-196. [ Links ]
42. MARIOTTINI, Y., M. L. de WYSIECKI & C. E. LANGE. 2011b. Seasonal occurrence of life stages of Grasshopper (Orthoptera: Acridoidea) in the Southern Pampas, Argentina. Zool. Stud 50 (6): 737-744. [ Links ]
43. MARIOTTINI, Y., M. L. de WYSIECKI & C. E. LANGE. En prensa. Diversidad y distribución de acridios (Orthoptera: Acridoidea) en pastizales del sur de la región pampeana, Argentina. Rev. Biol. Trop. [ Links ]
44. OLFERT , O. O. & R. M. WEISS. 2006. Bio-climatic model of Melanoplus sanguinipes (Fabricius) (Orthoptera: Acrididae) populations in Canada and the potential impacts of climate change. J. Orthoptera. Res. 15: 65-77. [ Links ]
45. ONSAGER, J. A. & J. E. HENRY. 1977. A method for estimating the density of rangeland grasshoppers (Orthoptera, Acrididae) in experimental plots. Acrida. 6: 231-237. [ Links ]
46. PERELMAN, S. B., R. J. C. LEON & M. OESTERHELD. 2001. Cross-scale vegetation patterns of Flooding Pampa Grasslands. J. Ecol. 89: 562-577. [ Links ]
47. RONDEROS, R. A. 1986. "Stability and diversity of grasshoppers species due to spatial heterogeneity" En: Proc. 4th Trienn. Meeting, Pan Amer. Acridol. Soc. 1985, USA, pp. 121-124. [ Links ]
48. SÁNCHEZ, N. E. 1980. Variaciones estacionales de la fauna de Acrídidos en un pastizal sucesional de la Provincia de Buenos Aires (Argentina). Rev. Soc. Entomol. Argent. 39 (3 4): 227-234. [ Links ]
49. SÁNCHEZ, N. E. & M. L. DE WYSIECKI. 1993. Abundancia y diversidad de acridios (Orthoptera: Acrididae) en pasturas de la Provincia de La Pampa, Argentina. RIA. 24 (1): 29-39. [ Links ]
50. SCHAFFERS, A. P., I. P. RAEMAKERS., K. V. SÝKORA & C. J. F. TER BRAAK. 2008. Arthropod Assemblages Are Best Predicted by Plant Species Composition. Ecology. 8: 782-794. [ Links ]
51. SCHEINER, S. M. & J. GUREVITCH. 2001. Design and analysis of ecological experiments, 2nd ed. Oxford University, New York. [ Links ]
52. STAT SOFT, Inc. 2004. STATISTICA (data analysis software system), version 7. www.statsoft.com. [ Links ]
53. SERVICIO METEOROLOGICO NACIONAL. 2009. Informe Sobre Sequía. Disponible en: http://www.smn.gov.ar/?mod=clima&id=75 [ Links ]
54. TORRUSIO, S. A., M. M. CIGLIANO & M. L DE WYSIECKI. 2002. Grasshopper (Orthoptera: Acrididae) and plant community relationships in the Argentine pampas. J. Biogeogr. 29: 221-229. [ Links ]
55. TORRUSIO, S. A. & J. OTERO. 2009. Monitoreo de tucuras, análisis de imágines Landsat 5 TM realizado en la CONAE. Programa Nacional de monitoreo de tucuras, [ Links ]
56. ZHONG-WEI, G., L. HONG-CHANG & G. YA-LING. 2006. Grasshoppers (Orthoptera: Acrididae) biodiversity and grassland ecosystems. Insect. Science. 13: 221-227. [ Links ]
