










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. vol.74 no.3-4 La Plata dic. 2015
TRABAJO CIENTÍFICO
Nuevas asociaciones entre Phasiinae (Diptera: Tachinidae) y Pentatomidae (Hemiptera: Heteroptera) fitófagos en la pampa ondulada (Argentina) y descripción del macho de Dallasimyia bosqi Blanchard
New associations between Phasiinae (Diptera: Tachinidae) and Pentatomidae (Hemiptera: Heteroptera) phytophagous insects in the rolling pampa (Argentina) and description of Dallasimyia bosqi Blanchard male
Liljesthröm, Gerardo G.1 & Delia S. Avalos2
1Centro de Estudios Parasitológicos y de Vectores (CEPAVE), Universidad Nacional de La Plata. Calle 2 N° 584, 1.900, La Plata, Argentina. E-mail: gerardo@cepave.edu.ar
2Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. Avda. Valparaíso s/n. Ciudad Universitaria. Córdoba, Argentina. E-mail: d-avalos@agro.unc.edu.ar
Recibido: 2-X-2014
Aceptado: 10-IX-2015
RESUMEN. Las relaciones entre parasitoides Phasiinae (Diptera: Tachinidae) y sus huéspedes Pentatomidae (Hemiptera: Heteroptera) son escasamente conocidas en la Argentina. Se presentan nuevas asociaciones hospedador-parasitoide en la pampa ondulada, y se describe por primera vez el macho adulto de Dallasimyia bosqi Blanchard. Se efectuaron muestreos quincenales de hemípteros adultos durante primavera-verano de 2005, 2006 y 2012 en áreas de vegetación espontánea y cultivos de trébol rojo y soja en campos de los partidos de La Plata, Magdalena y Chivilcoy (Buenos Aires). Los hemípteros capturados se criaron bajo condiciones controladas, los que tenían huevos de parasitoides pegados en el corion se mantuvieron hasta su muerte, y aquellos sin evidencia de parasitismo se mantuvieron hasta 25-30 días. Cada huésped muerto fue depositado en un recipiente con tierra húmeda y cubierto con una malla de nylon. Los parasitoides adultos emergidos fueron acondicionados para su identificación. Se registraron cinco nuevas asociaciones: E. meditabunda - Cilindromyia brasiliana y Homogenia sp., D. furcatus -Trichopoda argentinensis y Dallasimyia bosqi, y C. armigera - Trichopoda giacomellii, y la ya conocida N. viridula - Trichopoda giacomellii. Colecciones de referencia se encuentran depositadas en el Museo de La Plata, Argentina (MLP) y en el Museu de Zoologia da Universidade de São Paulo, Brasil (MZSP).
PALABRAS CLAVE: Pentatomidos; Soja; Trébol rojo; Parasitoides; Koinobiontes
ABSTRACT. The relationships between Phasiinae (Diptera:Tachinidae) parasitoid flies and their Pentatomidae (Hemiptera: Heteroptera) hosts are little known in Argentina. We present new host-parasitoid associations in the rolling pampa, and describe for the first time the adult male of Dallasimyia bosqi Blanchard. Adult Hemiptera were sampled biweekly during spring-summer 2005, 2006 and 2012 in areas of natural vegetation and crops of red clover and soybean, in fields in La Plata, Magdalena and Chivilcoy counties (Buenos Aires province). The Hemiptera were reared under controlled conditions; those with parasitoid macrotype eggs stuck on the corium were kept till death, and those with no signs of parasitism were kept for up to 25-30 days. Each dead host was placed in a container with damp soil and covered with a nylon mesh. The adult parasitoids emerged were conditioned for identification. Five new associations were recorded: E. meditabunda - Cilindromyia brasiliana (Townsend) and Homogenia sp., D. furcatus - Trichopoda argentinensis (Blanchard) and Dallasimyia bosqi, and C. armigera - Trichopoda giacomellii (Blanchard) as well as the already known N. viridula - Trichopoda giacomellii. Reference collections are deposited in the Museo de La Plata, Argentina (MCNLP) and in the Museu de Zoologia da Universidade de São Paulo, Brazil (MZSP).
KEY WORDS: Pentatomids; Soybean; Red clover; Parasitoids; Koinobionts
INTRODUCCIÓN
Los Tachinidae conforman una de las más grandes familias de Diptera, y sus especies se agrupan en cuatro subfamilias: Phasiinae (incluyendo las tribus Trichopodini y Cylindromyiini), Dexiinae, Exoristinae y Tachininae (Wood, 1987). Se considera que representan un grupo monofilético (Wood, 1987; Pape, 1992; Cerretti et al., 2014) y sus integrantes se caracterizan por ser parasitoides de artrópodos, con fuerte preferencia por los insectos (Stireman et al., 2006; o’Hara, 2008). Dado que la mayoría de los huéspedes son fitófagos, la presencia de taquínidos podría influenciar la estructura de sus comunidades (Hawkins, 1994) limitando o regulando poblaciones plaga (Coombs, 2004; Liljesthröm & Rabinovich, 2004). A pesar de su abundancia, diversidad y larga historia en programas de control biológico (Clausen, 1962; De Bach, 1964; Huffaker, 1971), el conocimiento taxonómico, biológico y ecológico de esta familia es escaso (Stireman et al., 2006).
La subfamilia Phasiinae exhibe la menor riqueza específica y la mayor variabilidad morfológica (Crosskey, 1976). Es considerada una de las más primitivas por sus hábitos de ataque: depositar huevos macrotípicos no embrionados sobre o dentro del huésped (Wood, 1987). Se han especializado en parasitar a los Heteroptera (mayoritariamente Pentatomidae, Coreidae y Lygaeidae) (Townsend, 1934-1944; Blanchard, 1966; Crosskey, 1976; Arnaud, 1978; Guimarães, 1977). Las especies de Phasiinae se caracterizan por ser parasitoides koinobiontes, solitarios, generalmente sinovigénicos. La larva I penetra en el huésped y una vez completado el desarrollo larval lo abandona y se entierra para empupar. Unos pocos representantes poseen un ovipositor bien desarrollado con el que encastran sus huevos dentro del huésped (o’ Hara, 1985; Grenier & Liljesthröm, 1991; Stireman et al., 2006) y la fecundidad de las especies conocidas es de 100-300 huevos (Shahjahan, 1968; Coombs, 1997). El efecto del parasitismo de los Phasiinae sobre los Pentatomidae se traduce fundamentalmente en la muerte temprana del huésped y la reducción del número potencial de sus posturas (Liljesthröm & Rabinovich, 2004).
Los miembros de la tribu Trichopodini constituyen un grupo primariamente tropical y subtropical, muy bien representado en la región Neotropical (Guimarães, 1971; o’ Hara & Wood, 2004), aunque unos pocos miembros se hallan presentes en la región Etiópica, en las islas del sudeste asiático, Melanesia y Queensland (Crosskey, 1973), en la zona del Mediterráneo (Salerno et al., 2002) y también han sido exitosamente introducidos en Australia (Liljesthröm, 1994; Coombs & Sands, 2000; Coombs, 2004). La tribu Cylindromyiini es de amplia distribución en el mundo, algunos de sus representantes son comunes e importantes parasitoides de una gran variedad de Hemiptera, siendo los Pentatomidae sus huéspedes preferidos, y varias de sus especies se hallan citadas de la Argentina (Guimarães, 1976).
Los Pentatomidae numéricamente dominantes en cultivos de soja de la pampa ondulada (Argentina) son, en orden decreciente, Piezodorus guildinii (Westwood), Nezara viridula (Linnaeus), Dichelops furcatus (Fabricius), Edessa meditabunda (Fabricius), Chinavia armigera (Stål) y Edessa rufomarginata (De Geer). Todas estas especies son señaladas como las de mayor impacto en el cultivo de soja, alfalfa y trébol. tanto por los daños directos que ocasionan sobre la semilla como por los costos que insume su manejo (Trumper & Edelstein, 2008).
En la Argentina, la información taxonómica publicada de los Phasiinae es escasa y se halla referida a la Tribu Trichopodini (Blanchard, 1944, 1966), los géneros Trichopoda, Trichopodopsis y Eutrichopodopsis (Blanchard, 1966; Mallea et al., 1977; Liljesthröm, 1992), y la descripción de una hembra de Dallasimyia bosqi Blanchard (Blanchard, 1944). La información sobre aspectos bioecológicos se restringe a Trichopoda giacomellii (Blanchard) (La Porta & Crouzel, 1985; Liljesthröm, 1991, 1992, 1993a,b, 1997) y a la interacción con el huésped N. viridula (Liljesthröm & Bernstein, 1990; Liljesthröm & Rabinovich, 2004). Respecto a la tribu Cylindromyiini, si bien varias especies son citadas de la Argentina (Guimarães, 1976), en nuestro país no se ha publicado sobre sus aspectos taxonómicos ni bioecológicos.
En este trabajo se dan a conocer cinco nuevas asociaciones entre parasitoides Trichopodini/Cylindromyiini y huéspedes Pentatomidae fitófagos, y la primera descripción del macho de D. bosqi. Además, complementando con información ya publicada, se presenta la matriz de interacciones entre los Tachinidae parasitoides y las especies de Pentatomidae mencionadas en la pampa ondulada (Argentina).
MATERIALES Y METODOS
Sitio de recolección
Los pentatómidos adultos fueron recolectados en vegetación espontánea y cultivos de trébol rojo y soja situados en cuatro sitios distribuidos en tres partidos (La Plata, Magdalena y Chivilcoy) de la provincia de Buenos Aires (Argentina): La Plata (Estación Experimental Agropecuaria de Los Hornos (FCA y FUNLP) (35° 02´ S, 58° 05´W) de 17 ha), Magdalena (una explotación comercial en la localidad de Bavio (35° 05´ S, 57° 44´ W) de 11 ha) y Chivilcoy (dos campos comerciales cultivados con soja de 50 ha. cada uno, 34°54´ S, 60° 02´ W). El trébol rojo se implantó en la Estación Experimental y no se realizaron aplicaciones de pesticidas ni cortes mecánicos. Los cultivos de soja se sembraron bajo labranza mínima, se efectuaron dos aplicaciones del herbicida glifosato al 48% (diferentes marcas comerciales) combinado con un insecticida (el más empleado fue Cipermetrina a 80 cc/ha), y cada tres años se hacían rotaciones con maíz.
Se realizaron muestreos quincenales utilizando el paño vertical (20 unidades muestrales distribuidas al azar) y red de arrastre (10 golpes de red en 10 transectas seleccionadas al azar) durante noviembre-abril de 2005 y 2006 y octubre-noviembre de 2012.
Cría en laboratorio
Todos los pentatómidos capturados (con y sin evidencias de parasitismo) se mantuvieron en recipientes y jaulas de cría individuales. Los individuos con evidencia de parasitismo (huevos pegados en el corion) se criaron bajo condiciones controladas: 25 °C ±1 °C, 70 % HR y fotoperíodo 14 hs luz. Se alimentaron con maíz (Zea mays L.), vainas de soja (Glycine max L.) y porotos (Phaseolus vulgaris L.) y fueron observadas hasta su muerte. En cambio, aquellos sin evidencia de parasitismo fueron mantenidos en cautiverio hasta 25-30 días.
Cada huésped adulto muerto fue depositado en un recipiente cilíndrico con tierra húmeda y cubierto con una malla de nylon de 2x2 mm para permitir la emergencia de las larvas del parasitoide. Fueron revisados diariamente hasta la emergencia de los adultos. Si al cabo de 40 días no se registraba emergencia del adulto, se disecaban los puparios para verificar la muerte del parasitoide. Los parasitoides fueron adecuadamente acondicionados para su posterior identificación. Junto con los respectivos puparios y la especie huésped fueron depositados en la colección de la División Entomología, Museo La Plata (MLP).
La identificación taxonómica de los huéspedes fue realizada por la Dra. M.C. Coscarón de la División Entomología (MLP).
Los ejemplares de Tachinidae fueron previamente identificados por los autores de este trabajo utilizando descripciones originales, claves taxonómicas (Townsend, 1908; Blanchard, 1944; Guimarães, 1976) y realizando comparaciones con ejemplares de las colecciones del Instituto Nacional de Tecnología Agropecuaria (INTA) Castelar, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (Buenos Aires) y Museo de La Plata. Dichos ejemplares fueron enviados posteriormente al Dr. Silvio Nihei, del Museu de Zoologia da Universidade de São Paulo, Brasil (MZSP) para su corroboración y se encuentran depositados en este museo.
Dos especímenes identificados por los autores: uno como Homogenia sp. (1 ♂ ex Edessa meditabunda de soja, (Chivilcoy), 14.XII.2009, G. Liljesthröm), y otro como Cylindromyia brasiliana (TT 1927) (1 ♂ ex Dichelops furcatus de soja (Chivilcoy), 14.XII.2006, G. Liljesthröm), fueron enviados al Dr. Silvio Nihei para su corroboración y se encuentran depositados en la colección del MZSP. A excepción de estos dos ejemplares, una muestra de T. giacomellii, los restantes adultos y el pupario de Homogenia sp. fueron depositados en el Museo de La Plata (MLP).
Para confeccionar la matriz de interacciones, se trabajó con datos obtenidos a partir de la cría de parasitoides provenientes de los huéspedes aquí colectados y de la literatura referida a las citas de parasitoides de las especies de pentatómidos mencionadas.
RESULTADOS
El número total de pentatómidos adultos colectados y aquellos parasitados se distribuyeron en las siguientes especies: Piezodorus guildinii (n=279, parasitados=0), Nezara viridula (n=179, parasitados=90), Dichelops furcatus (n=75, parasitados=14), Edessa meditabunda (n=71, parasitados=11) y Chinavia armigera (n=11, parasitados=3).
De N. viridula emergieron n=60 (67%) ejemplares de Trichopoda giacomelli: 33 ♂ y 27 ♀ (Chivilcoy y La Plata, sobre soja, G. Liljesthröm, MLP).
El número y porcentaje de parasitoides adultos emergidos de D. furcatus fue n=9 (64%): 7 ejemplares pertenecientes a Trichopoda argentinensis: 3 ♂ y 4 ♀ (Chivilcoy y La Plata, los huéspedes pentatómidos fueron colectados sobre soja y trébol rojo, G. Liljesthröm, MLP), y 2 ♂ fueron identificados como Cylindromyia brasiliana (2 huéspedes ♂ colectados sobre soja, (Chivilcoy), 14.XII.2006, G. Liljesthröm: un ejemplar en el MZSP y otro en MLP).
Se obtuvieron n=5 (45.5%) parasitoides adultos del huésped E. meditabunda: un individuo perteneciente a Homogenia sp. (MZSP), y los 4 restantes identificados como Dallasimyia bosqi: 1 ♂ y 3 ♀ (Chivilcoy y La Plata, los huéspedes fueron colectados sobre soja, G. Liljesthröm, MLP).
Emergieron n=3 (100%) parasitoides adultos de C. armigera: todos pertenecientes a Tr ichopoda giacomellii: 1 ♂ y 2 ♀ (La Plata, los huéspedes fueron colectados sobre plantas de tomate, G. Liljesthröm, MLP).
En la Tabla I se presentan las cinco nuevas asociaciones entre Trichopodini/Cylindromyiini y chinches Pentatomidae fitófagos registradas en este trabajo, más la obtenida de bibliografía publicada en la Argentina. En la Tabla II se presentan características del ciclo vital de los parasitoides.
Tabla I. Asociaciones entre huéspedes Pentatomidae y parasitoides Trichopodini /Cylindromyiini en la pampa ondulada (Argentina). El doble asterisco representa una nueva asociación huésped-parasitoide.
Tabla II. Algunas características del ciclo vital de los parasitoides.
Descripción del macho de Dallasimyia bosqi
Largo total del cuerpo: 9 mm. Cutícula cefálica ocre, placa ocelar y lúnula negruzcas, al igual que la frontalia. Parafrontalias con pruina dorada y parafacialias con pruina más leve, plateada. Facialias, lámina facial y epistomio con pruina tenue, plateada. Bordes externos de las facialias negruzcos. Altura cefálica 3.0 mm, ancho 4.0 mm, eje antenal 1.84 mm, y eje vibrisal 1.44 mm. Perfil oral 1.56 mm, perfil frontal 1.91 mm, y perfil facial 1.51 mm. Frontalia 0.80 mm ancho menor y 0.89 mm ancho mayor. Cerdas ocelares presentes, pero su tamaño y proyección se desconocen por estar cortadas. Cerdas frontales (FRL) 8 pares que se extienden desde más abajo del nivel del ocelo anterior hasta el borde superior de la lúnula. Parafrontalias 0.18 mm de ancho menor y 0.40 mm de ancho mayor, calvas. Lámina facial de 1.11 mm de alto x 1.20 mm de ancho. Facialias de 0.27 mm de ancho medio y con 6-7 sétulas negras en el nivel vibrisal. Vibrisas 0.53 mm. Parafacialias de 0.37 mm de ancho medio. Ancho máximo del epistomio: 1.60 mm. Genas: 0.22 mm, o sea 0.09 del eje máximo ocular: 2.60 mm. ojos calvos: 2.62 mm de alto máximo y 1.33 mm de ancho máximo. Antenas negruzcas, I artejo muy corto, con las bases separadas, II artejo de perfil subtriangular y 0.31 mm de largo, con finos pelos negros; III artejo de 0.67 mm de largo por 0.31 mm de ancho. El III artejo es 2.16 veces más largo que el II. Arista se desconoce pues falta en el ejemplar.
Cutícula torácica ocre, mesonoto con 3 fajas negras, la mediana se extiende desde el foramen occipital hasta el medio de la postsutura; las laterales desde la línea posthumeral hasta antes del callo post alar, ligeramente separadas de la mediana en la presutura y contiguas con ésta en la postsutura. Las tres fajas se interrumpen apenas en la presutura. Mesonoto 2.88 mm: presutura 1.24 mm y postsutura 1.64 mm. Callo humeral con 2 cerdas humerales (HM) y varios finos pelos negros, 2 notopleurales (NP), 1 presupralar (PRSA), 2 presuturales (PRS), 1 postsupraalar (PSA), 1 post intralar (PI), 2 postsuturales con la anterior corta (PS), y 1 postacrostical (PA), y 1 propleural (PPLR). Mesopleura con finos pelos negros en la mitad superior y 6-7 cerdas mesopleurales (MSPLR), 1 pteropleura (PTPLR) fuerte y 1 fina y varios finos pelos negros. Esternopleura con sternopleurales (STPL) 2 (+1). El escutelo es ocre parduzco, subtriangular: 1.2 mm de largo y 1.96 mm ancho máximo, con 1 lateral escutelar (LS), y 1 apical escutelar (APS).
Alas negruzcas en casi toda la superficie, excepto el borde inferior subhialino, algo irregular de 0.33 mm (10.7% del ancho alar máximo). Largo alar máximo 8.17 mm y ancho máximo 3.07 mm, el largo es 2.66 veces el ancho. Pre-estigma 2.13 mm, estigma 1.32 mm, postestigma 2.27 mm, y radiocosta 0.85 mm. La vena R5: 2.22 mm (1ra sección) y 3.54 mm (2da sección), Vena M2: 3.35 mm (1ra sección) y 2.22 mm (2da sección). La vena M1: 1.56 mm, Vena C1: 3.17 mm (1ra sección) y 0.47 mm (2da sección), y la vena anal de 1.94 mm. La celda 5R cerrada antes del extremo alar, con un corto pecíolo (Fig. 1).
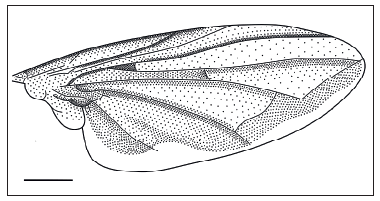
Fig. 1. Dallasimyia bosqi ♂: vista dorsal del ala. Escala 1 mm.
Patas de color ocre anaranjado. Tibias posteriores con una serie apretada de aproximadamente 17-18 setas negras, achatadas, plumiformes. Ancho máximo de tibia posterior 0.47 mm y el largo de la misma es de 3.40 mm, y las setas plumiformes ocupan 2.60 mm (76% del largo tibial). El largo de las setas plumiformes es de 0.30 mm, luego, las setas plumiformes representan 64% del ancho máximo tibial (Fig. 2).

Fig. 2. Dallasimyia bosqi ♂: tibia posterior. Escala 1 mm.
Abdomen anaranjado claro, con los últimos dos segmentos algo más oscurecidos y con una leve pruina plateada. Largo y ancho del abdomen: 6.0 mm y 2.59 mm, respectivamente. Basisegmento visible. Segmentos I, II, III y IV sub-iguales. Segmentos II a IV con 1 cerda mediano marginal (MM) apenas diferenciadas. Segmentos III, IV y V sin mediano marginales diferenciales y sin marginales laterales (ML).
Genitalia del macho. Ventral. Las forcípulas internas de los forceps anales están fusionadas en una sola estructura ancha con la proyección mediana inferior, subcilíndrica y dirigida en sentido postero-superior y es 2.10 veces la longitud de las proyecciones laterales (0.17 mm), que son ligeramente curvadas hacia adentro y dirigidas en sentido posterolateral. En la Fig. 3 se representan los fórceps anales.

Fig. 3. Dallasimyia bosqi ♂: representación esquemática de forceps anales. Escala 1 mm.
Pupario. Mide 8 mm de largo y 3 mm de ancho.
Material examinado. 1 ♂ (Paratipo), Los Hornos (La Plata), 25.X.2005, G. Liljesthröm.
DISCUSIÓN
A pesar de los esfuerzos en la recolección y cría de P. guildinii durante el desarrollo de este trabajo no se obtuvieron parasitoides de esta especie. En Florida (USA) se ha registrado a Trichopoda pennipes (Fabricius) parasitando a P. guildinii (Panizzi & Smith, 1976), y los parasitoides adultos emergidos resultaron de una dimensión menor a la habitual, probablemente debido al pequeño tamaño del huésped. En Brasil se cita a éste pentatómido parasitado por el taquínido Eutrichopodopsis nitens Blanchard (sinonimizado por Liljesthröm (1992) con Trichopoda giacomellii). Sin embargo, estas asociaciones parecen representar sólo sucesos aislados (Panizzi & Slansky, 1985) y corresponderían probablemente a eventos en los que los parasitoides se encuentran en situación "time limited", por lo que no serían interacciones huésped-parasitoide persistentes.
El huésped D. furcatus fue exitosamente parasitado por C. brasiliana. Este parasitoide ha sido señalado atacando a Euchistus heros (Fabricius) en Brasil (Correa-Ferreira, 1984). otro parasitoide obtenido de este huésped fue T. argentinensis, constituyendo la primera cita sobre un pentatómido. Hasta el momento esta especie de Tachinidae sólo se había registrado como parasitoide del Coreidae Athaumastus haematicus (Stål) (Blanchard, 1966).
Edessa meditabunda fue parasitada por D. bosqi y Homogenia sp. La primera era sólo conocida, hasta el momento, por la descripción de un ejemplar hembra encontrado en la provincia de Buenos Aires (Blanchard, 1944), sin referencia alguna a una especie huésped. Respecto del género Homogenia Wulp, la literatura revisada señala a Homogenia dysderci (Townsend) parasitando a Dysdercus sp., Dysdercus peruvianus (Guerin-Méneville) y D. ruficollis (Linnaeus) (Hemiptera: Phyrrocoridae) (Mendes, 1938; Townsend, 1937, 1938). En Uruguay Neobrachelia edessa (Townsend) fue registrada como parasitoide de E. meditabunda (Guimarães, 1977) y en Argentina, Liljesthröm (1980) reportó a T. giacomellii como parasitoide circunstancial de éste hemíptero, sin alcanzar a completar su desarrollo. Se encuentra ampliamente documentado que este parasitoide selecciona con preferencia adultos y ninfas grandes de Nezara viridula (La Porta, 1987; Liljesthröm, 1991;1992), aunque se señalan otros huéspedes eventuales (Liljesthröm, 1980, 1992). Sands & Coombs (1999) mencionan que en Australia T . giacomellii fue liberada para controlar a N. viridula y también parasitó a otras especies aunque sin completar en ellas su desarrollo.
Cuando el cultivo de soja comenzó su expansión en la región pampeana, en la década del 70, N. viridula incrementó marcadamente su abundancia adquiriendo el "status" de plaga principal en soja (La Porta & Crouzel, 1985). En esa etapa N. viridula, una especie exótica establecida, podría haber sido atacada exitosamente por T. giacomellii, una especie Neotropical, y se habría establecido desde entonces una interacción persistente. El pentatómido C. armigera, de distribución Neotropical, podría ser uno de los huéspedes habituales de T. giacomellii.
BIBLIOGRAFÍA CITADA
1. ARNAUD, P. H. 1978. A host parasite catalog of North American Tachinidae (Diptera). U.S. Dept. of Agriculture Miscellaneous Publication, Washington D.C. [ Links ]
2. BLANCHARD, E. E. 1944. Dallasimyia bosqi, nuevo género y especie de triquiopodino (Dipt. Gymnosomatidae). Revista Argentina de Entomología 2: 7-11. [ Links ]
3. BLANCHARD, E. E. 1966. Nuevos triquiopodinos argentinos, parásitos de hemípteros nocivos (Diptera, Gymnosomatidae). Revista de Investigaciones Agropecuarias 3: 59-95. [ Links ]
4. CLAUSEN, C. P. 1962. Entomophagous insects. Hafner Publishing Company, New Cork. [ Links ]
5. COOMBS, M. 1997. Influence of adult food deprivation and body size on fecundity and longevity of Trichopoda giacomellii: A South American parasitoid of Nezara viridula. Biological Control 8: 119-123. [ Links ]
6. COOMBS, M. 2004. Overwintering survival, starvation resistance, and postdiapause reproductive performance of Nezara viridula (L.) (Hemiptera: Pentatomidae) and its parasitoid Trichopoda giacomelli Blanchard (Diptera: Tachinidae). Biological Control 30: 141-148. [ Links ]
7. COOMBS, M. & D. A. SANDS. 2000. Establishment in Australia of Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae), a biological control agent for Nezara viridula (L.) (Hemiptera: Pentatomidae). Australian Journal of Entomology 39: 219-222. [ Links ]
8. CORREA-FERREIRA, B. 1984. Incidência do parasitóide Eutrichopodopsis nitens Blanchard, 1966 em populações do percevejo verde Nezara viridula (Linnaeus, 1758). Annais da Sociedade Entomológica do Brasil 13: 321-330. [ Links ]
9. CROSSKEY, R. W. 1973. A conspectus of the Tachinidae (Diptera) of Australia, including keys to the supraspecific taxa and taxonomic and host catalogues. Bulletin of the British Museum of Natural History (Entomology), Supp. 21: 1-221. [ Links ]
10. CROSSKEY, R. W. 1976. A taxonomic conspectus of the Tachinidae (Diptera) of the oriental Region. Bulletin of the British Museum of Natural History (Entomology), Supp. 26: 1-357. [ Links ]
11. DE BACH, P. 1964. Biological control of insect pests and weeds. De Bach, Chapman and Hill Ltd., London. [ Links ]
12. CERRETTI, P., J. E. O’HARA, D.M. WOOD, H. SHIMA, D. J. INCLÁN & J. O. STIREMAN III. 2014. Signal through the noise? Phylogeny of the Tachinidae (Diptera) as inferred from morphological evidence. Systematic Entomology 39: 335-353.
13. GRENIER, S. & G. G. LILJESTHRÖM. 1991. Préférences parasitaires et particularites biologiques des Tachinaires (Diptera: Tachinidae). Bulletin Mensuel de la Société Linéenne de Lyon 60(4): 128-141. [ Links ]
14. GUIMARÃES, J. H. 1971. A Catalogue of the Diptera of the Americas South of the United States. Family Tachinidae (Larvaevoridae). Museu de Zoologia, Universidade de São Paulo, São Paulo. [ Links ]
15. GUIMARÃES, J. H. 1976. A revision of the genus Cylindromyia Meigen in the Americas south of the United States (Diptera, Tachinidae). Arquivos de Zoologia 27(1): 1-150. [ Links ]
16. GUIMARÃES, J. H. 1977. Host-Parasite and Parasite-Host catalogue of South American Tachinidae (Diptera). Arquivos de Zoologia 28(3): 1-131. [ Links ]
17. HAWKINS, B. A. 1994. Patterns and process in host-parasitoid interactions. Cambridge University Press, Cambridge Mass. [ Links ]
18. HUFFAKER, C. B. 1971. Biological Control. Plnum Press. New York, London. [ Links ]
19. LA PORTA, N. C. 1987. Aspectos biológicos de Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae) parasitoide de Nezara viridula (L.) 1758. Hem. Pentatomidae. Revista de la Sociedad Entomológica Argentina 44: 433-439. [ Links ]
20. LA PORTA, N. C. & I. S. de CROUZEL. 1985. Estudios básicos para el control biológico de Nezara viridula (L.1758) (Hemiptera: Pentatomidae) en la Argentina. Revista de la Sociedad Entomológica Argentina 43: 119-166. [ Links ]
21. LILJESTHRÖM, G. G. 1980. Nota sobre Trichopoda giacomellii (Blanchard, 1966) (Diptera: Tachinidae). Neotropica 26: 233-236. [ Links ]
22. LILJESTHRÖM, G. G. 1991. Selectividad del parasitoide Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae) hacia individuos de Nezara viridula (L.) (Hemiptera: Pentatomidae) que difieren en el estado de desarrollo, sexo, edad y patrones de coloración. Ecología Austral 1: 41-49. [ Links ]
23. LILJESTHRÖM, G. G. 1992. Revisión de las especies de los géneros Trichopoda Berthold, Trichopodopsis Townsend y Eutrichopodopsis Blanchard descriptas para la República Argentina. Revista de la Sociedad Entomológica Argentina 50: 51-71. [ Links ]
24. LILJESTHRÖM, G. G. 1993a. Superparasitismo y competencia intraespecífica entre larvas del parasitoide Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae). Ecología Austral 3: 43-48. [ Links ]
25. LILJESTHRÖM, G. G. 1993b. Efectos del parasitismo de Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae) sobre una población de Nezara viridula L. (Hemiptera: Pentatomidae). Revista de la Sociedad Entomológica Argentina 52: 21-28. [ Links ]
26. LILJESTHRÖM, G. G. 1994. Primer envío de Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae) a Australia, para el control de Nezara viridula (L.) (Hemiptera: Pentatomidae). Neotropica 40: 89-90. [ Links ]
27. LILJESTHRÖM, G. G. 1997. Persistencia de Trichopoda giacomellii (Diptera: Tachinidae) durante el período de inactividad invernal del hospedador Nezara viridula (Hemiptera: Pentatomidae) en el nordeste de la provincia de Buenos Aires. Revista de la Sociedad Entomológica Argentina 56: 133-136. [ Links ]
28. LILJESTHRÖM, G. G. & C. BERNSTEIN. 1990. Density dependence and regulation in the system Nezara viridula (Hemiptera: Pentatomidae), host and Trichopoda giacomellii (Blanchard) (Diptera: Tachinidae), parasitoid. Oecologia 84: 45-52. [ Links ]
29. LILJESTHRÖM, G. G. & J. RABINOVICH. 2004. Modeling biological control: the population regulation of Nezara viridula and Trichopoda giacomellii. Ecological Applications 14: 254-267. [ Links ]
30. MALLEA, A., G. MACOLA, J. GARCÍA, L. BAHAMONDES, J. SUAREZ & S. LANATI. 1977. Trichopodopsis gustavoi n. sp. (Diptera- Tachinidae- Gymnosomatidae- Trichopodini) parásito de Nezara viridula (L.) Stål. Revista de la Facultad de Ciencias Agropecuarias 10: 17-20. [ Links ]
31. MENDES, L. O. T. 1938. Lista dos inimigos naturaes de Dysdercus spp. observados no estado de Sao Paulo. Revista de Entomologia 9: 215-217. [ Links ]
32.O’HARA, J. E. 1985. Oviposition strategies of the Tachinidae, a family of beneficial parasitic flies. Agriculture and Forestry Bulletin, University of Alberta 8: 31-34.
32.O’HARA, J. E. 2008. Tachinid flies (Diptera: Tachinidae). En: J. Capinera (ed.), Encyclopedia of Entomology 4 (S-Z), Springer-Verlag, pp. 3675-3686.
33. O ’HARA, J. E. & M. WooD. 2004. Catalogue of the Tachinidae (Diptera) of America North of Mexico. Memoirs on Entomology, International 18: 1-410.
34. PANIZZI, A. R. & F. SLANSKY Jr. 1985. Review of phytophagous pentatomids (Hemiptera: Pentatomidae) associated with soybean in the Americas. Florida Entomologist 68: 184-214. [ Links ]
35. PANIZZI, A. R. & J. G. SMITH. 1976. ocorrencia de Pentatomidae em soja no Paraná durante 1973/74. Biológico 42: 173-176. [ Links ]
36. PAPE, T. 1992. Phylogeny of the Tachinidae family-group (Diptera:Calyptratae). Tijdschrift voor Entomologie 135: 43-86. [ Links ]
37. SANDS D. A. & M. CooMBS. 1999. Evaluation of the Argentinian parasitoid, Trichopoda giacomellii (Diptera: Tachinidae) for biological control of Nezara viridula (Hemiptera: Pentatomidae) in Australia. Biological Control 15: 19-24. [ Links ]
38. SALERNO, G., S. COLAZZA & F. BIN. 2002. Nezara viridula parasitism by the tachinid fly Trichopoda pennipes ten years after its accidental introduction into Italy from the New Word. Biocontrol 47: 617-624. [ Links ]
39. SHAHJAHAN, M. 1968. Superparasitization of the Southern green stink bug by the tachinid parasite Trichopoda pennipes pilipes and its effect on the host and parasite survival. Journal of Economic Entomology 61: 1088-1091. [ Links ]
40. STIREMAN, J. O., J. E. O ’HARA & D. M. WOOD. 2006. Tachinidae: evolution, behavior, and ecology. Annual Review of Entomology 51: 525-555.
41. TOWNSEND, C. H. T. 1908. The taxonomy of the muscoideanflies, including descriptions of new genera and species. Smithsonian Institution 1803: 1-138. [ Links ]
42. TOWNSEND, C. H. T. 1934-1944. Manual of Myiology, 15 parts, Charles Townsend and Filhos Itaquaquectuba, Sao Paulo, Brasil. [ Links ]
43. TOWNSEND, C. H. T. 1937. New fly parasites of Dysdercus. Revista de Entomologia 7: 316-318. [ Links ]
44. TOWNSEND, C. H. T. 1938. Further fly parasites of Dysdercus. Revista de Entomologia 8: 347-348. [ Links ]
45. TRUMPER, E. & J. EDELSTEIN. 2008. El complejo de chinches fitófagas en soja: revisión y avances en el estudio de su ecología y manejo. 1ª Edic. C.A. de Bs. As. Inst. Nacional de Tecnología Agropecuaria- INTA, Buenos Aires. [ Links ]
46. WOOD, D. M. 1987. Tachinidae. En: McAlpine J. F, B. V. Peterson, G. E. Shewell, H. J.Tes key, J. R. Vockeroth & D. M. Wood (eds.), Manual of Nearctic Diptera, Vol. 2. Agric. Can. Monogr. 28: 675–1332.

