










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. vol.75 no.3-4 La Plata dic. 2016
Trabajo Científico
Preferencia de oviposición de Peregrinus maidis (Hemiptera: Delphacidae) en distintas plantas hospederas
Oviposition preference of Peregrínus matáis (Hemiptera: Delphacidae) on different host plants
DIAZ, Claudia N.1, Erica LUFT ALBARRACIN2 & Mariela ALDERETE3
1Instituto de Invertebrados, Fund. M. Lillo, Miguel Lillo 251, T4000JFE, San Miguel de Tucumán, Argentina.
2PROIMI-Biotecnología (CONICET), Div. Control Biológico, Av Belgrano y Pje. Caseros, (4000) San Miguel de Tucumán, Argentina. E-mail: erluft@hotmail.com
3Cátedra de Ecología, Fac. Cs. Nat. e IML, Universidad Nacional de Tucumán, Tucumán, Argentina.
Received: 01-IV-2016; accepted: 21-IX-2016; published: 23-XII-2016
RESUMEN. La chicharrita del maíz, Peregrinus maidis (Ashmead), es una especie que ha sido registrada en la mayoría de las regiones tropicales. Esta asociada a diversos cultivos de importancia económica. En América tropical, esta especie es un vector de diversas enfermedades virales en maíz, Zea mays L. El objetivo de esta investigación es evaluar la preferencia de oviposición de P. maidis sobre diferentes especies vegetales hospederas, mediante un ensayo de elección múltiple. Se ofrecieron cuatro especies de Poaceae: Z. mays L., Brachiaria sp., Cynodon dactylon (L.) Person y Sorghum halepense (L.) Person. Se detectó que las hembras de P. maidis son capaces de depositar sus huevos en todas las especies vegetales ofrecidas, aunque prefieren significativamente el sorgo de alepo, maíz y pasto bra-quiaria (91% de los casos). Por otra parte, se registró un alto porcentaje de plantas disponibles sin huevos, sugiriendo que P. maidis no ovipone durante un período de tiempo considerable. Los resultados son discutidos en el contexto de las interacciones agroecológicas presentes en un sistema de cultivo de maíz.
PALABRAS CLAVES. Poaceae. Maíz. Chicharrita del maíz. Comportamiento de oviposición. Selección de plantas hospederas.
ABSTRACT. The corn planthopper, Peregrinus maidis (Ashmead) is a species that has been recorded in most regions of the Tropics. It is associated to several crops of economical importance. In tropical America, the species is a vector of several viral diseases, mainly in maize, Zea mays L. The aim of this research is to evaluate the oviposition preference of P. maidis on different host plants, by means of a multiple choice test. In this study, four species of Poaceae, Z. mays, Brachiaria sp., Cynodon dactylon (L.) Person and Sorghum halepense (L.) Person, were offered. We found that P. maidis females are able to lay their eggs in all offered plant species, although they significantly preferred Johnsongrass, maize and Brachiaria (91% of cases). Moreover, a high percentage of plants without eggs were recorded, suggesting that P. maidis spends considerable time without ovipositing. Results are discussed in the context of P. maidis agroecological interactions in the crop system of maize.
KEY WORDS. Poaceae. Maize. Corn planthopper. Oviposition behaviour. Host plant selection.
Peregrinus maidis (Ashmead) (Hemiptera: Delphacidae), es una chicharrita asociada a diversos cereales de importancia económica, como el maíz Zea mays L., el sorgo Sorghum bicolor L. y el mijo Panicum miliaceum L., a nivel mundial. En América tropical este delfácido es una reconocida plaga del maíz, vector de diversas enfermedades virales, entre ellas el Virus del Mosaico del Maíz (MMV), el Virus de la Hoja Blanca del Maíz (HBMV), el Virus del Mosaico del Bandeado del Maíz (MStpV), el Virus del Enanismo Rugoso del Maíz (MRDV), y los virus Maize Sterile Stunt y Maize stripe (MSpV) (Laguna & Giménez Pecci, 2012). Además del maíz, estas enfermedades también afectan al sorgo y otras gramíneas (Tsai & Brown, 1994). En Argentina, las enfermedades de mayor relevancia transmitidas por P. maidis son el Virus del Mal de Río Cuarto (MRCV), actualmente la enfermedad más perjudicial que afecta al cultivo de maíz, y en menor medida el MMV (Giménez Pecci et al., 2002). Virla et al. (2004) detectaron que P. maidis se comporta como un vector experimental del MRCV, es decir, en condiciones controladas en laboratorio, es capaz de transmitir el virus. Dada su importancia sanitaria, es necesario realizar estudios biológicos y de comportamiento de P. maidis para acrecentar el conocimiento sobre esta plaga en el país.
Peregrinus maidis se distribuye en áreas tropicales y subtropicales de todo el mundo y es considerada una plaga importante en América (principalmente en México, Perú, Venezuela), África y Oceanía (Tsai et al., 1986; Rioja, 2006; Singh & Seetharama, 2008). En la Argentina este del-fácido se registró por primera vez en provincias del norte, asociado al maíz y posteriormente fue encontrado en Buenos Aires en cultivos de sorgo (Tesón & Remes Lenicov, 1989; Remes Lenicov et al., 2006). Exhibe hábitos polífagos, está presente en una gran variedad de cultivos cerealeros y en las malezas asociadas a estos agroeco-sistemas. Éstas son utilizadas como hospederas para la alimentación y oviposición (Caldwell & Martorell, 1950; Remes Lenicov & Virla, 1999; Remes Lenicov & Mariani, 2001; Remes Lenicov et al., 2006). El rol de los hospederos alternativos en la epidemiología de las enfermedades es de gran significancia en el estudio ecológico de este vector (Tsai & Perrier, 1993; Tsai, 1996).
El maíz tiene una gran importancia para la economía de la Argentina, es el segundo cultivo de granos después de la soja, y junto con el trigo representan el 86 % de la producción de cereales de la Argentina. Su principal destino es la exportación, además de cumplir un rol en la rotación de cultivos y la generación de bioenergías (Eyhérabide, 2012). Dentro de los factores que limitan la producción de maíz se destacan las enfermedades (Saini, 2005), que pueden estar presentes a través de reservorios en la vegetación natural circundante al cultivo de maíz. Luft Albarracin et al., (2008) detectaron que el agroecosistema del maíz en el noroeste argentino está compuesto principalmente por mono-cotiledóneas consideradas malezas. Éstas son perjudiciales para estos hábitats debido a que representan una vegetación perenne y resistente a herbicidas, afectando la producción del cultivo (Gudelj et al., 2005).
Los delfácidos se hallan íntimamente asociados a las plantas hospederas, para su alimentación, reproducción, protección contra las condiciones desfavorables del ambiente, y como refugio contra los enemigos naturales, encontrándose el 65% de las especies relacionadas con monocotiledóneas. Varios trabajos, a nivel mundial han abordado investigaciones referidas a la explotación del hospedante como recurso para la reproducción por delfácidos (Mochida, 1964; Khan & Saxena, 1985; Tavella & Arzone, 1992; Iitomi, 1995; Tsai, 1996; Hattori & Sogawa, 2002). En Argentina existen algunos antecedentes sobre el ciclo de vida y comportamiento de oviposición de Delphacodes kuscheli Fennah (Virla & Remes Lenicov, 1991; Brentassi & Remes Lenicov, 1999), y la preferencia de alimentación y sitios de oviposición de Chionomus haywardi (Muir) (Virla & Maragliano, 1993).
El estudio del comportamiento de oviposición en insectos vectores de enfermedades, con un amplio rango de especies vegetales hospederas, como es el caso de P. maidis, permite profundizar el conocimiento sobre su biología reproductiva y el riesgo de transmisión de fitopatógenos en distintas especies vegetales, tanto en cultivos como en malezas presentes en los agroecosistemas. Este conocimiento puede ser aplicado como una herramienta para el manejo integrado de plagas, en casos en que esta chicharrita adquiera el estatus de plaga. Por lo tanto, el objetivo del presente trabajo es evaluar la preferencia de oviposición de P. maidis sobre cuatro especies vegetales de Poaceae presentes en el agroecosistema maíz en Argentina.
MATERIALES Y MÉTODOS
Preparación de plantas, insectos y diseño de experimentos
Se usaron individuos de P. maidis provenientes de una cría estable, mantenida en un invernáculo bajo condiciones controladas (24± 2 °C, 60 - 80% HR, y fotoperiodo natural), en el laboratorio de la división de Control Biológico de PROIMI (CONICET, Tucumán). Para conocer la preferencia en la selección de plantas de P. maidis para la oviposición, se realizó un ensayo de elección múltiple en una cámara de cría bajo condiciones controladas (24± 1°C, 70 % HR y fotoperiodo 14/10 hs, luz/oscuridad). Se utilizaron cuatro especies vegetales de la familia Poaceae como sustrato de oviposición: Z. mays L. (maíz), Brachiaria sp. (pasto braquiaria), Cynodon dactylon (L.) Person (gramón) y Sorghum halepense (L.) Person (sorgo de alepo). Estas especies fueron elegidas por estar ampliamente distribuidas en los sistemas agrícolas de la Argentina (Rodríguez et al., 1997; Luft Albarracin et al., 2008). El material correspondiente al sorgo de alepo, pasto braquiaria y gramón fue extraído de un campo experimental ubicado detrás del laboratorio de Control Biológico de PROIMI, San Miguel de Tucumán (26º48`36.2”S, 65º14`28.3”O, 470 msnm). La unidad experimental fue una jaula de vidrio de 20 x 20 x 35 cm de alto, sellada con dos aberturas cubiertas con tela voile como respiraderos.
En cada jaula se colocaron las cuatro especies vegetales en macetas individuales de 250 cm3 con una disposición espacial aleatoria. Para uniformar la superficie foliar de las cuatro plantas se consideró un área foliar de 30 cm2, se midió el área foliar tomando como referencia al área que presenta en promedio una planta de maíz en estado vegetativo V3 (área foliar: largo x ancho max. x 0,75) (Montgomery, 1911). En el caso del sorgo de alepo y pasto braquiaria, esto se logró cultivando dos plantas por maceta mientras que para el gramón, cuya área foliar es menor a las otras especies, se cultivó una cantidad mayor de plantas por maceta. Para el maíz, se sembró una planta por maceta dos semanas previas al inicio de los ensayos y la variedad usada fue Leales 25.
Una vez armada la unidad experimental, se liberó una hembra de P. maidis oviplena (hembra en el periodo de oviposición, con abdomen voluminoso, lleno de huevos maduros). Transcurridos 10 minutos de la liberación de la hembra, se observó su ubicación dentro de la jaula, y se registró si P. maidis estaba asentada en un sustrato vegetal (especie vegetal), lo cual indicaba su preferencia, o fuera de las plantas (en macetas, paredes de jaula), exhibiendo movimientos triviales que se consideraron como de no preferencia.
Se realizaron observaciones cada dos horas en las primeras 10 horas, una observación a las 24 horas y otra a las 48 horas, obteniéndose un total de siete observaciones por cada jaula. Una vez terminado el ensayo, se contabilizó la cantidad de huevos depositados en cada planta y por jaula. Cada jaula fue considerada una réplica, contabilizándose en total 41 repeticiones. Dado que la hembra de P. maidis deposita los huevos endofíticamente, principalmente en el interior de las vainas foliares, en la nervadura central de las hojas y sus cercanías, el recuento de los mismos se llevó a cabo con un microscopio estereoscópico. Para ello, se realizó la disección de toda la planta.
Análisis de datos
En el ensayo de elección múltiple, se calculó un índice de Manly de preferencia de selección de recurso vegetal (Manly et al., 2003): donde oi representa la proporción de plantas con huevos pertenecientes a la especie vegetal i (i = maíz, sorgo de alepo, gramón, pasto braquiaria). ui es la cantidad de plantas de la especie vegetal i donde se registraron huevos y ut es la cantidad total de plantas utilizadas para la oviposición. Por otra parte, πi representa la proporción de plantas de la especie i disponibles para oviponer, siendo con Ai representando a la cantidad de plantas disponibles en la categoría i y At al tamaño poblacional total de las especies de plantas disponibles.
Teniéndose en cuenta que m es igual al total de categorías (en este caso cuatro tipos de sustratos vegetales diferentes), hay tres resultados posibles para interpretar la preferencia: I, no hay preferencia para oviponer en la especie vegetal i, 1 α i = m; II , hay preferencia para oviposición en la especie vegetal i cuando 1 α i > m y, III, la especie vegetal i es evitada para oviponer cuando α i < 1m. Para evaluar si P. maidis depositaba los huevos de manera selectiva significativamente, se realizó una prueba de χ2 de Pearson aplicando la siguiente fórmula: Σ= y se comparó con χ2 con m - 1 grado de libertad. Nuevamente, ui es la cantidad observada de plantas con huevos en la categoría i, mientras que u+πi representa la cantidad esperada bajo la hipótesis de no selección (al azar).
Para cada réplica de ensayo de elección múltiple se registró la fecundidad total, como la suma de la cantidad de huevos colocados en todos los sustratos por la hembra, y la simple preferencia computada como ya mencionada.
Por otra parte, para evaluar la existencia de asociación lineal entre preferencia de oviposición (porcentaje de huevos colocados en un sustrato vegetal particular por cada organismo) y fecundidad (cantidad de huevos total registrada por cada individuo), se puso a prueba el uso de correlación no paramétrica (Spearman). Además, se compararon separadamente los promedios de las variables de preferencia de oviposición, y fecundidad total entre los cuatro sustratos vegetales. Debido a la falta de homogeneidad de varianzas, se aplicó la prueba de ANAVA (análisis de varianza) no paramétrica Kruskal-Wallis junto con comparaciones de a pares de las medias de los rangos (Di Rienzo et al., 2008)
Por otro lado, para detectar si las hembras utilizaban de manera diferente el tiempo sobre el sustrato vegetal u otro tipo de sustrato, de total de las siete observaciones se cuantificó la proporción de tiempo que cada hembra dedicó a cada tipo de sustrato (sustrato vegetal o no vegetal). Las diferencias en la proporción de tiempo fueron evaluadas con un ANAVA no paramétrico Kruskal-Wallis, considerando los cuatro sustratos vegetales y el sustrato no vegetal (5 tratamientos). Además, para confirmar la presencia de una asociación lineal entre el uso del tiempo por parte del individuo (proporción de tiempo sobre el sustrato vegetal y sobre sustrato no vegetal) versus fecundidad se puso a prueba la correlación según el estadístico r Spearman
Todos los análisis de este trabajo fueron realizados mediante el uso del software estadístico INFOSTAT (Di Rienzo et al., 2008)
RESULTADOS
P. maidis utilizó las cuatro especies vegetales para depositar sus huevos, pero el número de huevos depositados en cada una de ellas fue variado. Del total de 575 huevos contabilizados en todo el ensayo de elección múltiple, un 91% fueron colocados en maíz, sorgo de alepo y pasto braquiaria (Tabla I). La restante especie de gramínea fue menos seleccionada.
Con respecto al índice de Manly de preferencia de selección de recurso vegetal, el valor puntual umbral fue de 0,25 (1/4), detectándose qué P. maidis presentó una preferencia por maíz y sorgo de alepo (a = 0,3 para ambos), evitó oviponer en gramón (a = 0,15) y en pasto braquiaria fue indiferente (a = 0,25); aunque no se registró una preferencia de manera significativa estadísticamente por parte del vector al momento de oviponer en diferentes sustratos vegetales (χ2= 4,24; p > 0,05).
Por otra parte, los promedios de preferencia en oviposición y fecundidad entre los cuatro sustratos vegetales resultaron significativamente diferentes (H = 7,83; H = 7,58 respectivamente, p < 0,05; n=41) (Figs. 1 y 2). No hubo diferencias significativas entre los tratamientos maíz, sorgo y braquiaria. Mientras que sí se detectaron diferencias entre maíz-sorgo-braquiaria y el tratamiento gramón pero sin diferencias entre este último tratamiento y braquiaria. Los menores valores de preferencias en oviposición y fecundidad fueron registrados en gramón. Además, sin tener en cuenta el tipo de sustrato vegetal donde se realizó la oviposición, se encontraron altas correlaciones positivas entre preferencia de oviposición versus fecundidad (r2 = 0,93; p < 0,05, n=164).


El promedio de la proporción de tiempo que empleó la hembra de P. maidis en cada especie de planta y por fuera de la planta (sustrato no vegetal) mostró diferencias significativas, siendo mayor por fuera de la planta, seguido por el maíz, sorgo de alepo y braquiaria conjuntamente y menor en el gramón (H = 37,55; p < 0,05, Fig. 3). Se confirma el uso del tiempo y la fecundidad por parte del insecto al comprobarse una correlación negativa entre la fecundidad y tiempo que el insecto pasó sin elegir ninguna planta donde oviponer (r = -0,53; p < 0,05) y una correlación positiva entre el tiempo en que el insecto se posó para oviponer en una planta y la cantidad de huevos colocados (r = 0,71; p < 0,05).
DISCUSIÓN Y CONCLUSIÓN
Al evaluar el comportamiento de preferencia de P. maidis sobre las especies vegetales utilizadas, el estudio indicó que las hembras del vector eligen oviponer en sorgo de alepo, maíz y braquiaria. Numerosos estudios (Chelliah & Basheer, 1965; Namba & Higa, 1971; Tsai, 1996) demostraron que diferentes especies de sorgo son los hospederos preferidos de P. maidis para su alimentación y desarrollo, lo cual estaría asociado a que algunas especies del género Sorghum se postulan como los hospederos ancestrales de esta chicharrita (Nault, 1983). El sorgo de alepo, una maleza común en los sistemas agrícolas, podría actuar como una planta alternativa para P. maidis cuando el cultivo de maíz no está presente, y contribuir de esta manera con la permanencia persistente de sus poblaciones en los agroecosistemas.
Con respecto a la planta de maíz, este estudio coincide con lo reportado por otros autores (Tsai, 1996; Catindig et al., 1996), los cuales observaron que P. maidis deposita un mayor número de huevos en plantas de maíz, cuando se le ofrecen distintas especies vegetales.
Por otra parte, el gramón fue la especie de gramínea menos seleccionada por P. maidis para depositar sus huevos. En este vegetal, a su vez, se observó que, en general los huevos fueron depositados en los tallos de las plantas, por lo que se infiere que la baja oviposición se debe a que esta planta presenta hojas y tallos muy delgados. En este sentido, Begoña Riquelme et al. (2013) encontraron que Aerenea quadriplagiata (Boheman) (Coleoptera: Cerambicidae) prefieren oviponer en plantas de Amaranthus L. con tallo de diámetro 30% mayor entre los cultivares ofrecidos.
La elección de plantas por parte de un insecto depende de características tales como: defensas químicas, defensas físicas, la madurez y dureza del tejido foliar, entre otras (Awmack & Leather, 2002; Mannion et al., 2003; Meiners et al., 2005; Coley et al., 2006).
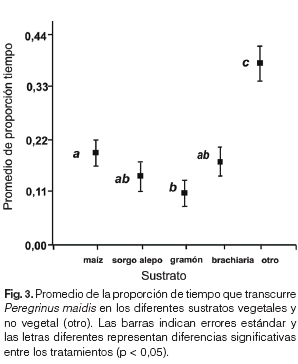
Es interesante confirmar la coincidencia de resultados obtenidos entre los valores puntuales del índice de Manly de preferencia de selección de recurso vegetal y los resultados del ANAVA, aunque la prueba de χ2 de Pearson detectó que P. maidis depositó los huevos de manera aleatoria. Esto podría deberse a que los 41 insectos sólo utilizaron 63 plantas de un total de 164 disponibles para oviponer. El alto porcentaje de plantas disponibles sin posturas podría deberse a que en ambientes donde la chicharrita percibe olores de distintos sustratos vegetales pasaría mayor tiempo sin oviponer. Este estudio demostró que al ofrecerle a P. maidis diferentes sustratos vegetales para oviponer, utilizó mayor tiempo en un sustrato no vegetal (38%). Estos resultados coinciden con observaciones realizadas en un ensayo de no elección donde se utilizaron jaulas de 500 cm3 PET con respiraderos y en cada una se colocó una planta de maíz con igual superficie foliar que en el ensayo de elección múltiple y se liberó una hembra oviplena durante 48hs (N = 14). La cantidad de huevos contabilizada fue superior en promedio a lo registrado en ensayo de elección múltiple en las jaulas de vidrio de 20 x 20 x 35cm (35 ± 13,43 huevos). Sería interesante confirmar estos resultados previos obtenidos en ensayos con jaulas comparables. Además, se registró una correlación positiva entre el tiempo que el vector se posa sobre un sustrato vegetal y la cantidad de huevos colocados en el ensayo de elección múltiple.
Por otra parte, la evidencia de que P. maidis tuvo una menor fecundidad en la planta de maíz cuando estuvo enfrentada a una oferta variada de otras especies vegetales, puede ser útil para el manejo de esta plaga. Una mayor heterogeneidad vegetal en el paisaje agrícola, como la presencia de bordes no cultivados en un maizal, además de proporcionar refugio para los enemigos naturales, provocaría un menor ataque de la plaga a las especies preferidas, entre ellas el cultivo de maíz. Los estímulos distintos ante diversos sustratos, interferiría en la detección de sustratos vegetales preferidos y, consumiría un mayor tiempo en la búsqueda de un sustrato conveniente para alimentarse y oviponer. En este sentido, Costa et al. (2009) señalaron que Eurysacca melanocampta Meyrick (Lepidoptera: Gelechiidae) prefirió una variedad de planta de Chenopodium quinoa Willdenow para oviponer, y esta preferencia se basó en estímulos del olfato que dependen de los metabolitos y compuestos químicos emitidos por la planta hospedera. Por otra parte, Shenk et al. (1983) mencionaron que la gran diversidad de plantas asociadas a labranza cero en el cultivo de maíz puede reducir el ataque de insectos herbívoros, aumentar el número de insectos depredadores o proveer plantas alternativas para la alimentación; esto se traduce en un incremento del rendimiento del cultivo.
Si bien P. maidis no es considerada una plaga en la Argentina, sí lo es en otros países latinoamericanos como México, Perú y Venezuela (Rioja, 2006; Singh & Seetharama, 2008). La realización de futuras investigaciones que tengan en cuenta el papel que juegan los enemigos naturales de P. maidis, en su área de distribución, así como la interacción tritrófica: planta-insecto-enemigo natural, sería de utilidad para prevenir problemas sanitarios en los agroecosistemas. En la Argentina P. maidis podría encontrarse en bajas densidades controlado por sus enemigos naturales, tales como hongos entomopatógenos y parasitoides (Virla & Luft Albarracin, 2012) que conviven en un sistema heterogéneo en cuanto a la presencia de sustratos vegetales alternativos al maíz, principalmente en los bordes de los cultivos. Por lo tanto, sería importante realizar experimentos en condiciones de campo para poder explicar las interacciones ecológicas entre el cultivo y el vector, así como también entre éste y sus enemigos naturales, a fin de proponer al productor herramientas útiles en el marco de un programa de manejo integrado de plagas.
De este estudio, se concluye que P. maidis, cuando se lo expone al maíz y representantes de vegetación natural (sorgo de alepo, pasto braquiaria y gramón) prefiere oviponer en sorgo de alepo, maíz y braquiaria. Sin embargo, la fecundidad resulta afectada negativamente por los diferentes estímulos percibidos por el insecto. Sería interesante realizar estudios sobre los volátiles que liberan las plantas hospederas de P. maidis en los sistemas de cultivo, para evaluar su potencial rol como factor regulador de las poblaciones de esta chicharrita, y disminuir así la propagación de patógenos.
BIBLIOGRAFÍA CITADA
- Awmack, C.S. & Leather, S.R. (2002) Host plant quality and fecundity in herbivorous insects. Annual Review of Entomology, 47, 817-844. [ Links ]
- Begoña Riquelme, M.V., Ansa, M.A. & Santadino, M.V. (2013) Preferencia de oviposición del barrenador del amaranto, Aerenea quadriplagiata (Coleoptera, Cerambycidae) en condiciones de campo. Revista Colombiana de Entomología, 39(1), 76-80. [ Links ]
- Brentassi, M.E. & Remes Lenicov, A.M.M. (1999) Oviposición de Delphacodes kuscheli (Homoptera-Delphacidae) sobre plantas de cebada en condiciones de laboratorio. Revista de la Facultad de Agronomía, 104(1), 67-74. [ Links ]
- Catindig, J.D., Barrion, A.T. & Litsinger, J.A. (1996) Plant host range and life history of the corn delphacid, Peregrinus maidis (Ashmead) (Hemiptera: Delphacidae). Asia Life Sciences, 5, 35-46. [ Links ]
- Caldwell, J.S. & Martorell, L.F. (1950) Review of the Auchenor-rhynchous Homoptera of Puerto Rico. Part II. The Fulgoroidea. Journal Agricultural of University of Puerto Rico, 34, 133-270. [ Links ]
- Chelliah, S. & Basheer, M. (1965) Biological studies of Per-egrinus maidis (Ashmead) (Araeopidae: Homoptera) on sorghum. Indian Journal of Entomology, 27, 466-471. [ Links ]
- Coley, P.D., Bateman, M.L. & Cursar, T.A. (2006) The effects of plant quality on caterpillar growth and defense against natural enemies. Oikos, 11 5 (2), 219-228. [ Links ]
- Costa, J.F., Cosio, W., Cardenas, M., Yábar, E. & Gianoli, E. (2009). Preference of quinoa moth: Eurysacca melanocamp-ta Meyrick (Lepidoptera: Gelechiidae) for two varieties of quinoa (Chenopodium quinoa Willd.) in olfactometry Assays. Chilean Journal of Agricultural Research, 69(1), 71-78. [ Links ]
- Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M. & Robledo, C.W. (2008). InfoStat, versión 2008, Grupo InfoStat, FCA, UNC, Argentina. [ Links ]
- Eyhérabide, G.H. (2012) El maíz y su cultivo en Argentina. Contexto general y tecnológico. Enfermedades del maíz producidas por virus y mollicutes en Argentina (ed. Giménez Pecci, M.P., Laguna, I.G. & Lenardón, L.), pp. 7-24. INTA, C.A.B.A., Argentina. [ Links ]
- Giménez Pecci, M.P., Oliveira, E., Resende, R., Laguna, I., Con-ci, L., Avila, A., Herrera, P., Galdeano, E., Virla, E. & Nome, C. (2002) Ocorrencia de doenças causadas por molicutes e por vírus em milho nas províncias de Tucumán e de Córdoba na Argentina. Fitopatologia Brasileira, 27, 403-407. [ Links ]
- Gudelj, V., Vallone, P., Galarza, C. & Masiero, B. (2005) Evaluación de diferentes alternativas de herbicidas para el control de malezas en el cultivo del maíz. Revista Técnica de Asociación Argentina de Productores en Siembra Directa, 143-148. [ Links ]
- Hattori, M. & Sogawa, K. (2002) Oviposition behavior of the rice brown planthopper, Nilaparvata lugens (Stal), and its electronic monitoring. Journal of Insect Behavior, 15, 283-293. [ Links ]
- Iitomi, A. (1995) Oviposition site of Sogatella furcifera (Hor-vath) on rice plant in northern Japan. Annual Report of the Society of Plant Protection of North Japan, 46, 111-114. [ Links ]
- Khan, Z.R. & Saxena, R.C. (1985) Behavioral and Physiologi-cal Responses of Sogatella furcifera (Homoptera: Delphacidae) to Selected Resistant and Susceptible Rice Culti-vars. Journal of Economic Entomology, 78(6), 1280-1286. [ Links ]
- Laguna, L.I. & Giménez Pecci, M.P. (2012) Panorama Mundial de las enfermedades causadas por virus en el cultivo de maíz. Enfermedades del maíz producidas por virus y mol-licutes en Argentina (ed. Giménez Pecci, M.P., Laguna, I.G. & Lenardón, L.), pp. 31-40. INTA, C.A.B.A., Argentina.
- Luft Albarracin, E., Paradell, S. & Virla, E. (2008) Cicadellidae (Hemiptera: Auchenorrhyncha) associated with maize crops in northwestern Argentina, influence of the sow-ing date and phenology of their abundance and diversity. Maydica, 53, 289-296.
- Manly, B.F.J., Mcdonald, L.L., Thomas, D.L., Mcdonald, T.L. & Erickson, W.P. (2003) Resource selection by animals -Statistical design and Analysis for field studies. Kluwer academic publishers, London.
- Mannion, C., Hunsberger, A., Peña, J. & Osborne, L. (2003) Oviposition and larval survival of Diaprepes abbreviatus (Coleoptera: Curculionidae) on select host plants. Florida Entomologist, 86, 165-173.
- Meiners, T., Hacker N.K. & Hilker, M. (2005) Response of the elm leaf beetle to host plant induced by oviposition and feeding: the infestation rate matters. Entomologia Experi-mentalis et Applicata, 11 5 , 171-176.
- Mochida, O. (1964) On oviposition in the brown planthopper, Nilaparvata lugens (Stal). (Hom., Auchenorrhyncha). II. The number of eggs in an egg group, especially in relation to the fecundity. Japanese Journal of Applied Entomology and Zoology, 8, 141-148.
- Montgomery, E.G. (1911) Correlation studies of corn. Ne-braska Agricultural Station Annual Report, Lincoln, 24, 108-159.
- Namba, R. & Higa S.Y. (1971) Host plant studies of the corn planthopper, Peregrinus maidis (Ashmead), in Hawaii. Proceedings of the Hawaiian Entomological Society, 21, 105-108.
- Nault, L.R. (1983). Origin of leafhopper vectors of maize pathogens in Mesoamerica, En: Proceedings International Maize Virus Disease Colloquium & Workshop, 1976, Wooster, Ohio. Pp. 75-82.
- Remes Lenicov, A.M.M. & Mariani, R. (2001) Hallazgo del vector del virus del mosaico del maíz Peregrinus maidis (Homoptera: Delphacidae) en la provincia de Buenos Aires. Neotropica, 47, 48.
- Remes Lenicov, A.M.M. & Virla, E.G. (1999) Delfácidos asociados al cultivo de maíz en la República Argentina (Insecta: Homoptera: Delphacidae). Revista de la Facultad de Agronomía, 104, 1-15.
- Remes Lenicov, A.M.M., Paradell, S. & Catalano, M.I. (2006) Hemípteros auquenorrincos asociados al cultivo de sorgo en la Argentina (Insecta: Hemiptera). Revista de Investigación Agropecuaria, 35, 3-20.
- Rioja, T.C. (2006) Biología y Enemigos Naturales de Pereg-rinus maidis (Ashmead) (Hemiptera: Delphacidae) en el Valle de Azapa. IDESIA, 24, 1-48.
- Rodríguez, N., Faya De Falcón, L. & Pieri, S. (1997) Malezas. Reconocimiento de semillas y plántulas. INTA. EEA Manfredi-EEA Paraná.
- Saini, E.D. (2005) Insectos perjudiciales al cultivo de maíz y sus enemigos naturales. INTA, C.A.B.A., Argentina.
- Shenk, M.D., Saunders, J. & Escobar, G. (1983) Labranza Mínima y no Labranza en Sistemas de Producción de Maíz (Zea mays) para áreas Tropicales Húmedas de Costa Rica. Serie Técnica, Boletín Técnico N 8. Centro agronómico Tropical de Investigación y enseñanza (CAT-IE), Turrialba, Costa Rica.
- Singh, B.U. & Seetharama, N. (2008) Host plant interactions of the corn planthopper, Peregrinus maidis Ashm. (Homoptera: Delphacidae) in maize and sorghum agroecosys-tems. Arthropod-Plant Interactions, 2, 163-196.
- Tavella, L. & Arzone, A. (1992) Aspetti nutrizionale in Zyginidia pullula (Boheman), Empoasca vitis (Goethe) e Grapho-cephala fennahi Young. (Homoptera Auchenorrhyncha). Bollettino di Zoologia agraria e di Bachicoltura Serie II, 24(2), 137-146.
- Tesón, A. & Remes Lenicov, A.M.M. (1989) Contribución al estudio de los Fulgoroideos Argentinos IV. (Homoptera, Fulgoroidea) observaciones sobre tres especies halladas en cultivos de maíz (Zea mays L.). Revista de la Sociedad Entomológica Argentina, 47, 101-107.
- Tsai, J.H. (1996) Development and oviposition of Peregrinus maidis (Homoptera: Delphacidae) on various host plants. Florida Entomologist, 79(1), 19-26.
- Tsai, J.H. & Brown, L.G. (1994) Maize Stripe Tenuivirus. Plant Pathology Circular No. 363. Florida Department of Agriculture and Consumer Services, Division of Plant Industry, Gainesville, FL.
- Tsai, J.H. & Perrier, J.L. (1993) Morphology of the digestive and reproductive systems of Peregrinus maidis (Homoptera: Delphacidae). Florida Entomologist, 76, 428-436.
- 139-146, 2016
- Tsai, J.H., Wilson, S.W. & Faan, H.W. (1986) Peregrinus maidis (Homoptera: Delphacidae) from southern China. Journal of the New York Entomological Society, 94, 442-443.
- Virla, E. & Maragliano, R. (1993) Preferencias alimentarias y sitios de oviposición de Delphacodes haywardi (Muir, 1929) en diferentes hospeda dores, en condicio nes de laboratorio (Insecta: Homoptera: Delphacidae). Revista de la Sociedad Entomológica Argentina, 52(1-2), 1-6.
- Virla, E.G. & Luft Albarracin, E. (2012) Potencialidad de especies antagonistas en el manejo racional de poblaciones de vectores de enfermedades del maíz causadas por virus en Argentina. Enfermedades del maíz producidas por virus y mollicutes en Argentina (ed. Giménez Pecci, M.P., Laguna, I.G. & Lenardón, L.), pp. 151-168. INTA, C.A.B.A., Argentina.
- Virla, E. & Remes Lenicov, A.M.M. (1991) Ciclo de vida de Delphacodes kuscheli criado sobre distintos hospedantes en condiciones de laboratorio (Insecta-Homoptera-Delphacidae). En: Taller de actualización sobre "Mal de Río Cuarto". Pergamino-Buenos Aires. INTA-CIMMYT. Pp. 104-115.
- Virla, E., Miotti, I., Giménez Pecci, M.P., Carpane, P. & Laguna, G. (2004) Peregrinus maidis (Hem.: Delphacidae), new experimental vector of the «Mal de Río Cuarto» disease to corn. Biocell, 28(1), 54.

