










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
Print version ISSN 0373-5680On-line version ISSN 1851-7471
Rev. Soc. Entomol. Argent. vol.75 no.3-4 La Plata Dec. 2016
Trabajo Científico
El papel de la densidad y la proporción sexual de adultos en la fecundidad de Diatraea saccharalis (Lepidoptera: Crambidae) en jaulas de cría masiva
The role of density and sex ratio of adults in fertility of Diatraea saccharalis (Lepidoptera: Crambidae) in mass rearing cages
MEDINA PEREYRA, Pilar1, Mariano ORDANO2, Carmen REGUILÓN3, Analía R. SALVATORE4,6, César ACOSTA5 & Leopoldo RISSO6
1Instituto de Fisiología Animal, Fundación Miguel Lillo, Miguel Lillo 251, T4000JFE, San Miguel de Tucumán, Tucumán, Argentina. E-mail: mpmedina@lillo.org.ar
2Instituto de Entomología, Fundación Miguel Lillo, y Unidad Ejecutoria Lillo (UEL-FML-CONICET), Consejo Nacional de Investigaciones Científicas y Técnicas, Miguel Lillo 251, T4000JFE, San Miguel de Tucumán, Tucumán, Argentina.
3Instituto de Entomología, Fundación Miguel Lillo, Miguel Lillo 251, T4000JFE, San Miguel de Tucumán, Tucumán, Argentina.
4Estación Experimental Agroindustrial Obispo Colombres. Av. William Cross 3150, T4101XAC, Las Talitas, Tucumán, Argentina.
5Facultad de Bioquímica, Química y Farmacia, Universidad Nacional de Tucumán, Ayacucho 471, T4000INH, San Miguel de Tucumán, Tucumán, Argentina.
6Facultad de Agronomía y Zootecnia, Universidad Nacional de Tucumán, Av. Néstor Kirchner 1900, T4000ACS, San Miguel de Tucumán, Tucumán, Argentina.
RESUMEN. La cría de insectos para control biológico aumentativo requiere conocer los factores que afectan la fecundidad para optimizar la producción masiva. En general, las prácticas empleadas se basan en los resultados de experiencias exitosas. Sin embargo, en muchos casos la verificación de esas prácticas en condiciones distintas a las de su origen carece de soporte cuantitativo publicado y suele pasar por alto la variación derivada del manejo de los operarios (condición estándar). El objetivo de este trabajo es evaluar la fecundidad del barrenador de la caña de azúcar Diatraea saccharalis Fabricius, en función del número de adultos y de la proporción sexual (número de hembras / número de adultos) en jaulas de cría, en condiciones de producción estándar. En 25 jaulas se registraron número y sexo de las mariposas y número de huevos obtenido. Los resultados mostraron que la fecundidad óptima se obtiene con valores intermedios de densidad y con proporción sexual sesgada a machos. Se sugiere que el control conjunto de densidad y proporción sexual incrementa la eficiencia en las posturas de huevos. El presente trabajo es el primero en cuantificar ambos factores en condiciones de cría masiva y revela que las prácticas comúnmente extendidas necesitan de estudios de mayor profundidad.
PALABRAS CLAVE. Control biológico aumentativo. Cría masiva. Barrenador de caña de azúcar. Oviposición.
ABSTRACT. Insect rearing for augmentative biological control requires knowledge of the factors affecting fertility to optimize mass production. In general, the practices employed are based on the results of successful experiences. However, in many cases, the verification of such practices other than those of their original conditions lacks published quantitative support and often overlooks the variation derived from the operators handling (standard condition). The aim of this work is to evaluate the fertility of the sugarcane borer Diatraea saccharalis Fabricius, depending on the number of adults and sex ratio (number of females / number of adults) in rearing cages under standard production. In 25 cages, the number and sex of butterflies and the number of eggs obtained were recorded. Results showed that optimum fertility was obtained with intermediate density values and the sex ratio biased to males. It is suggested that the control of both density and sex ratio increases the efficiency of egg masses laid. This paper is the first to quantify both factors in mass rearing conditions, and reveals that the widespread practices commonly require further studies.
KEY WORDS. Augmentative biological control. Mass rearing. Sugarcane borer. Ovi-position.
Las larvas de Diatraea saccharalis Fabricius constituyen una de las plagas más importantes en el cultivo de la caña de azúcar. Al perforar los tallos, estos insectos producen galerías que provocan pérdidas en las cañas cosechadas e inciden negativamente en el proceso de extracción fabril (Willink, 1982; González, 1984; CAÑAMIP, 2000). Debido a su hábito barrenador, el uso de insecticidas para control es efectivo en los primeros estadios larvales, antes de que penetren los tallos que les confieren protección (Schex-nayder et al., 2001). Si bien el control químico es una práctica utilizada, el control biológico es un método más efectivo, empleado en diversos países con el beneficio de minimizar el impacto de los plaguicidas y sus residuos tóxicos sobre la salud humana y ambiental (Mendonça, 1996; Capinera, 2001; Salvatore et al., 2009).
Los programas de control biológico aumentativo para el manejo de insectos plaga requieren del establecimiento de sistemas de cría masiva. Generalmente los lineamientos de estos sistemas están basados en el conocimiento del ciclo biológico de los insectos y en ensayos que corroboran el uso de ciertas prácticas de cría masiva (Alam et al., 1971). Sin embargo, en ocasiones las mismas persisten sin evaluar las posibilidades de mejoramiento y eficiencia, necesarios para la reducción de costos de producción, y suelen pasar por alto la variación derivada del manejo de los operarios (condición estándar).
Un ejemplo de control biológico de Diatraea en cañaverales se desarrolla en Brasil con la producción y liberación de Cotesia flavipes Cameron (Botelho & Macedo, 2002). El sistema de cría de Diatraea y Cotesia en América es un caso de programa de control biológico aumentativo exitoso (Vacari et al., 2012; Reguilón et al., 2014). Para favorecer la rentabilidad de estos sistemas de producción masiva es importante conocer los factores que incrementan la eficiencia del proceso de cría. En particular, una mayor producción de larvas requiere de una eficaz producción de huevos.
La cría masiva de Diatraea se realiza en jaulas con múltiples individuos, donde se incluye un número de parejas sexadas durante el estado de pupa. En términos prácticos resulta inviable seleccionar pupas para controlar la proporción sexual necesaria para optimizar la producción de huevos, por lo que la preparación de jaulas para cópula y oviposición se realiza en base a la selección de parejas con individuos adultos (Cano et al., 2006). Sin embargo, la persistencia de esta técnica se debe a una razón de practicidad, por lo que es esperable una alta variación en la proporción sexual y el número de individuos manejados por los operarios. No obstante, aunque la práctica persiste, no hay estudios que hayan cuantificado este problema y revelen los puntos a tener en cuenta para optimizar la producción de huevos.
La proporción de oocitos inmaduros en la etapa de emergencia del adulto puede estimarse mediante el índice de ovigenia (IO), el cual relaciona el número de huevos depositados y el número de oocitos maduros encontrados en la hembra. El IO toma valores en el rango 0 a 1; un IO cercano a 1 corresponde a una mayor proporción de oocitos maduros, mientras que un IO cercano a 0 significa que la maduración de oo-citos tiene que completarse en el estado adulto (Jervis & Ferns, 2004). En este último caso, la obtención de recursos necesarios para completar la oogénesis limita la capacidad de oviposición de la hembra. Un IO = 0,32 encontrado en D.
saccharalis indica la ocurrencia de maduración de los oocitos durante la fase adulta y por lo tanto la necesidad de utilización de recursos para la maduración (Parra et al., 1999). Se considera que en el estado adulto de este lepidóptero resulta innecesaria la adquisición de nutrientes como recursos energéticos para la producción de huevos (Parra et al., 1999). Por lo tanto, se ha considerado que los recursos para la maduración de huevos provienen de dos fuentes: los nutrientes incorporados en la fase larval y aquellos transferidos por los machos (Milano et al., 2010). En relación a las estrategias reproductivas, existen evidencias de que la poliandria o tendencia de las hembras para copular varias veces con distintos machos favorece la fecundidad por un incremento en la cantidad neta de producto seminal recibido (Torres Vila et al., 2004; Torres Vila & Jennions, 2005), y afecta positivamente la oviposición y longevidad de los adultos (Wedell et al., 2002).
Aunque esta estrategia reproductiva es muy común entre los lepidópteros (Arnqvist & Nilsson, 2000), se sabe que las hembras de D. saccharalis muestran una alta ocurrencia de una sola cópula (Pérez & Long, 1964; Walker, 1965; Linares, 1987). Sin embargo, el conocimiento sobre el rol de la proporción sexual y la densidad en la producción de huevos del barrenador de la caña de azúcar es escaso, principalmente en condiciones de cría masiva. Por una parte, estudios previos se han realizado a pequeña escala (Campos Guevara, 1980). Por otro lado, la literatura disponible sobre producción y manejo a gran escala, solo hace referencia a la introducción de un número determinado de parejas (aproximadamente 40) de mariposas en jaulas de cría (Cano et al., 2006). Sin embargo, ningún estudio ha considerado la variación en el manejo de los operarios, que en condiciones de cría masiva es variable dado el volumen de material biológico manipulado. Más aún, no hay evidencias cuantitativas de cómo la densidad y la proporción sexual afectan la fecundidad en condiciones estándar de reproducción masiva. En consecuencia, el objetivo de este trabajo es evaluar cómo influyen la densidad y la proporción sexual en la fecundidad de D. saccharalis en condiciones de cría masiva.
MATERIALES Y MÉTODOS
Procedimiento
La cría desarrollada para este estudio se inició a partir de huevos de D. saccharalis procedentes de Biocontrol, Riberão Preto, Brasil y se estableció en el Centro de Investigaciones sobre Regulación de Poblaciones de Organismos Nocivos, Tucumán, Argentina (CIRPON, autorización cuarentenaria de SENASA Argentina, Servicio Nacional de Sanidad y Calidad Agroalimentaria). La dieta artificial para alimentación de larvas y otras características del sistema de producción masiva de D. saccharalis se describen en Reguilón et al. (2014). Para la obtención de polillas se ubicaron jaulones de estructura metálica (60 cm de ancho x 90 cm de alto x 30 cm de profundidad), cubiertos por una malla plástica (entramado de 4 mm) sujetada por velcro, en una habitación con temperatura media de 25,5 ºC, humedad relativa media de 65% y fotoperíodo 12L: 12O. En el interior de los jaulones se colocaron 15 bandejas de poliestireno expandido (22 x 17 cm) que contenían aproximadamente 300 pupas por bandeja, provenientes de un mismo lote o fecha de oviposición.
Una vez ocurrida la emergencia, los adultos se colocaron en tubos de PVC (referidos como “jaulas”) de 10 cm de diámetro y 18 cm de alto, cuyo interior se cubrió con una hoja de papel de 80 g/m2 como sustrato de oviposición. Como en otras situaciones estándar de cría masiva, que requieren minimizar el tiempo de operación en el proceso, se estableció una unidad de captura que consistió en la captura manual de adultos que se colocaron en un tubo de vidrio de 60 ml (3 cm de diámetro x 9 cm de alto) hasta completar el volumen del tubo. Esta técnica suele ser usada en condiciones de cría masiva para reemplazar el uso de aspiradores manuales o eléctricos que requieren mayor tiempo de captura y que por lo tanto son inviables en cría masiva. Además, debido a que son mariposas nocturnas, los adultos durante el día están en reposo y permiten una rápida captura con la técnica del tubo. De este modo, cada jaula (N = 25) fue completada con tres unidades de captura. Las hojas de papel fueron retiradas a las 24 horas y marcadas para la posterior evaluación de la producción de huevos. En cada jaula de cría se registró el número de mariposas y el sexo de cada una. Solo se tomaron en cuenta las posturas del primer día posterior al armado de la jaula (jaulas de un día). Esto permitió reducir el número de factores involucrados y favorecer la viabilidad del ensayo.
Para estimar el número de huevos por hoja se utilizó un modelo de regresión lineal en el que la variable respuesta (número de huevos por hoja) se estimó a partir de la relación entre el número de huevos contados por postura y el área de postura correspondiente. Para esto se tomaron fotografías digitales de posturas individuales, que fueron procesadas con el programa Image J (Rasband, 1997). Dado que las fotografías tenían asociada una escala, fue posible estimar el área cubierta por cada postura en mm2. De esta manera, el número total de huevos por hoja pudo ser predicho por el área total cubierta por huevos en la hoja de oviposición. Mediante un análisis de regresión (R2 = 0,81, F1,48 = 198,3; p < 0.00001; Fig. 1) el número de huevos se estimó como:
H = a + β * A;
Donde
H = número de huevos
a = intercepto (3,93)
β = coeficiente (2,55)
A = área de una postura medida en Image J
Consecuentemente, fue posible estimar el número total de huevos por hoja midiendo solamente el área cubierta por huevos en cada hoja de jaula.
El diseño de estudio consistió en montar las observaciones sobre las condiciones de cría masiva. Idealmente, debería ser un diseño factorial cruzado (ver Discusión). Sin embargo, se optó por diseñar el estudio en función de cómo normalmente se opera en cría masiva, una cuestión generalmente pasada por alto en los estudios de cría masiva y en los libros sobre la materia. Para cuantificar el número de mariposas por jaula, una vez muertas, se identificó el número de individuos por sexo. Con estos datos pudo estimarse la densidad y la proporción sexual, y relacionarlas con el número total de huevos por hoja de jaula. La proporción sexual fue expresada como número de hembras / número total de mariposas.
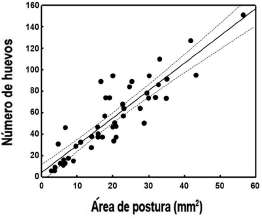
Fig. 1. Relación entre el número de huevos de D. saccharalis y el área medida por postura individual. Y= 3,93+2,55*área de postura (R2 = 0.81, F1,48 = 198.3, p < 0.00001).
Análisis de datos
Para estimar los efectos de la densidad y proporción sexual se aplicó un análisis de regresión múltiple, dado que las variables explicatorias son continuas. En este modelo se incluyeron los componentes lineales y cuadráticos del número total de mariposas por jaula y de la proporción sexual. Previamente estas variables fueron estandarizadas como (promedio de X) - Xi / (desvío estándar de X). La variable de respuesta fue el número de huevos estimado sin transformar, como fue descripto más arriba. Este análisis fue llevado a cabo con el programa STATISTICA 7.0 (Statsoft Inc., 2004).
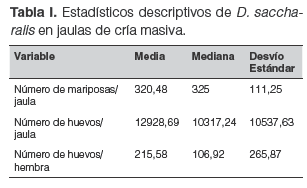
En la tabla I se muestra el resumen estadístico para cada una de las variables consideradas.
Los resultados del análisis de regresión múltiple aplicado sobre el número de huevos estimado mostraron que la cantidad de huevos que las mariposas ovipusieron en una jaula dependió linealmente de la proporción sexual y de manera no lineal del número de mariposas (Tabla II).
La proporción sexual obtenida por jaula se relacionó negativamente con el número de huevos estimado. Es decir, una proporción sexual sesgada a hembras (mayor número de hembras) disminuyó la cantidad de huevos obtenidos por hoja de oviposición (Fig. 2).
Por otra parte, el número de individuos por jaula mostró una relación no lineal con el número de huevos estimado. Los mayores valores del número de huevos estimado se relacionaron con valores intermedios de número de mariposas (aproximadamente 300; Fig. 3).


DISCUSIÓN
En este trabajo se evaluó el efecto de la densidad y la proporción sexual sobre la fecundidad de D. saccharalis. Los resultados evidencian dos puntos importantes para una implementación eficiente de la cría masiva de D. saccharalis. El primero es que el número de individuos que pueden ser incorporados por jaula es alto. Las recomendaciones sobre la utilización del espacio indican el empleo de 1 individuo / 10 cm2 de área (Campos Guevara, 1980). Sin embargo, en las condiciones experimentales del presente estudio, el área por individuo fue sensiblemente menor, por lo que se obtuvo una alta producción de huevos aun con grandes cantidades de adultos. El espacio (asociado a la densidad) juega un papel importante por un efecto de hacinamiento y competencia de los machos, y parece explicar la existencia de valores promedios de densidad óptimos para obtener una mayor producción de huevos, por encima de los cuales el rendimiento disminuye. El segundo punto es que la proporción sexual debería ser sesgada a machos. El efecto de este factor sobre la fecundidad podría explicarse por dos motivos o por la combinación de ambos. Por una parte, una mayor ocurrencia de machos disponibles contribuye a la eficiencia en la cantidad de hembras copuladas. La proporción óptima encontrada de 3:1 (machos:hembras) coincide con lo observado por Campos Guevara (1980) quien reporta proporción sexual sesgada a machos favorece la fecundidad al aumentar la posibilidad de encuentros de hembras con machos vírgenes, que resulta en una mayor producción de huevos. Este efecto se debe al tamaño del espermatóforo. Los machos vírgenes poseen un espermatóforo más grande que los que copularon previamente, debido a que sufren una depleción de los precursores de esta estructura luego de cada cópula (Torres Vila & Jennions, 2005).
Otra explicación a este hecho es el grado de poliandría o monandría. Si bien tiene una base genética que confiere a las hembras una capacidad diferencial para utilizar las donaciones de nutrientes de los machos (Wedell et al., 2002), se conoce que la existencia de factores fisiológicos y ambientales pueden regular la estrategia reproductiva en condiciones de laboratorio (Torres Vila et al., 1997). Es así que especies poliándricas forzadas a exhibir una estrategia monándrica ven disminuido su rendimiento en términos de fecundidad y longevidad, siendo su fecundidad significativamente menor que la de las monándricas en las mismas condiciones. Por el contrario, pueden recrearse las condiciones necesarias para que, hembras que en general exhiben la monandría como estrategia reproductiva, copulen con más de un macho (Torres Vila, 2013). Por lo tanto, la ocurrencia de apareamientos sucesivos aumentaría debido a la alta densidad de individuos y la proximidad forzada de ambos sexos en un ambiente espacialmente confinado.
En crías de laboratorio bajo condiciones controladas, Linares (1987) reporta para D. saccharalis una proporción de sexos de 1:0,87 machos:hembras. El hecho de que los machos no son un recurso limitado, debido a que su tasa reproductiva natural es mayor que la de las hembras, resulta en una proporción sexual sesgada a machos (Wedell et al., 2002), favoreciendo de este modo la posibilidad de optar por proporciones sexuales con menos hembras.
En conjunto, una mayor eficiencia en la obtención del número de huevos se daría en jaulas con aproximadamente 300 individuos y una proporción sexual sesgada a machos. Este estudio es el primero que considera ambas variables en condiciones de producción masiva. La realización de futuros estudios con niveles controlados de densidad de adultos sería de utilidad para estimar la magnitud del efecto de la interacción entre la proporción sexual y el número de individuos por jaula de cría. Esto permitiría controlar y balancear la cantidad de adultos de manera más eficiente para la producción de huevos, permitiendo manejar menos individuos con mejores resultados y con la consecuente disminución de los costos en el proceso de cría masiva.
AGRADECIMIENTOS
A Pablo Pereyra (Sección Iconografía, Fundación Miguel Lillo) por la asistencia gráfica. Al financiamiento, apoyo técnico y logístico del Ingenio y Refinería San Martín de Tabacal S. R. L., como parte del Proyecto "Control biológico de Diatraea saccharalis con Cotesia flavipes en extensiones cañeras del NOA", mediante convenio 2007-2010 con la Fundación Miguel Lillo.
BIBLIOGRAFÍA CITADA
- Alam, M.M., Bennett, F.D. & Carl, K.P. (1971) Biological control of Diatraea saccharalis (F.) in Barbados by Apanteles flavipes Cam. and Lixophaga diatraea T . T . Entomophaga, 16(2), 151-158. [ Links ]
- Arnqvist, G. & Nilsson, T. (2000) The evolution of polyandry: multiple mating and female fitness in insects. Animal Behaviour, 60(2), 145-164. [ Links ]
- Botelho, P.S. & Macedo, N. (2002) Cotesia flavipes para o controle de Diatraea saccharalis. Controle biológico no Brasil. Parasitóides e predadores (eds. Parra, J. R.,. Botelho, P.S.M, Correa Ferreira, B.S. &. Bento, J.M.S), pp. 409-426, Editorial Manole, São Paulo. [ Links ]
- Campos Guevara, L.A. (1980) Frequéncia de cópula da Diatraea saccharalis (Fabr., 1974) (Lepidoptera: Crambidae) em condições de campo e laboratório. Agropecuaria Técnica, 1(1), 24-31. [ Links ]
- Cano, M.A., Dos Santos, E.M. & Pinto, A.de S. (2006) Produção de Cotesia flavipes para o controle da broca da cana. Controle de pragas da cana de açúcar. Boletim Técnico Biocontrol 1, pp. 21-24. [ Links ]
- Cañamip (2000) Manejo integrado de barrenadores en caña de azúcar. Cengicaña, Guatemala. [ Links ]
- Capinera, J.L. (2001) Sugarcane Borer, Diatraea saccharalis (Fabricius) (Insecta: Lepidoptera: Pyralidae). Featured Creatures from the Entomology and Nematology Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida (disponible en http://creatures.ifas.ufl.edu) [ Links ]
- González, V. (1984) Perspectivas en la investigación sobre control del taladrador (Diatraea spp.) y la candelilla (Aeneolamia spp.) en caña de azúcar. En: II Seminario sobre los problemas de la candelilla y el taladrador en caña de azúcar y pastos, Barquisimeto, Venezuela, pp. 239-244. [ Links ]
- Jervis, M.A. & Ferns, P.N. (2004) The timing of egg maturation in insects: ovigeny index and initial egg load as measures of fitness and of resource allocation. Oikos, 170(3), 449-461. [ Links ]
- Linares, B.A. (1987) Influencia de la temperatura en el desarrollo de Diatraea saccharalis Fabricius. Caña de Azúcar. Revista del Instituto Nacional de Investigaciones Agropecuarias, Venezuela, 5(2), 43-66. [ Links ]
- Mendonça, A.F. (1996) Pragas da cana de açúcar. Maceió: Insetos & Cía., Maceió, Alagoas, Brasil.
- Milano, P., Filho, E.B, Parra, J.R.P., Oda, M.L. & Cônsoli, F.L. (2010) Efeito da alimentação da fase adulta na reprodu-ção e longevidade de espécies de Noctuidae, Crambidae, Tortricidae e Elachistidae. Ecology, Behavior & Bio-nomics. Neotropical Entomology, 39(2), 172-180.
- Parra, J.R.P., Milano, P. , Cônsoli, F.L., Zerio, N.G. & Haddad, M.L. (1999) Efeito da nutrição de adultos e da umidade na fecundidad de Diatraea saccharalis (Fabr.) (Lepidoptera: Crambidae). Anais da Sociedade Entomológica do Brasil, 28(1), 49-57.
- Pérez, R. & Long, W.H. (1964) Sex attractant and mating be-havior in the sugarcane borer. Journal of Economic Entomology, 57(5), 688-691.
- Rasband, W. (1997) Image J. National Institutes of Health, USA (disponible en http://rsb.info.nih.gov/ij/)
- Reguilón, C., Medina Pereyra, P., Ordano, M., Salvatore, A., Barros, M.V., Morsoletto Santos, E. & Vicente Cano, M.A. (2014) Evaluación de los efectos de la composición de la dieta artificial para la cría de Diatraea saccharalis (Le-pidoptera: Crambidae) y Cotesia flavipes (Hymenoptera: Braconidae). Revista de la Facultad de Ciencias Agrarias de la Universidad Nacional de Cuyo, 46(1), 45-57.
- Salvatore, A.R., López, G. & Willink, E. (2009) Plagas en el cultivo de caña de azúcar. Manual del Cañero (eds. Romero, E., Digonzelli, P.A. & Scandaliaris, J.) pp. 117-122. Estación Experimental Agroindustrial Obispo Colombres, Las Talitas, Tucumán, Argentina.
- Schexnayder, H.P. Jr., Reagan, T.E. & Ring, D.R. (2001) Sam-pling for the sugarcane borer (Lepidoptera: Crambidae) on sugarcane in Louisiana. Journal of Economic Entomo-logy, 94(3), 766-771.
- StatSoft, Inc. (2004) STATISTICA (data analysis software sys-tem), version 7 (disponible en www.statsoft.com)
- Torres Vila, L.M. (2013) Polyandry-fecundity relationship in in-sects: methodological and conceptual problems. Journal of Evolutionary Biology, 26, 325-334.
- Torres Vila, L.M. & Jennions, M.D. (2005) Male mating history and female fecundity in the Lepidoptera: do male virgins make better partners? Behavioral Ecology and Sociobio-logy, 57(4), 318-326.
- Torres Vila, L.M., Rodríguez Molina, M.C. & Jennions, M.D. (2004) Polyandry and fecundity in the Lepidoptera: can methodological and conceptual approaches bias out-comes? Behavioral Ecology and Sociobiology, 55(4), 315-324.
- Torres Vila, L.M., Stockel, J. & Rodríguez Molina, M.C. (1997) Physiological factors regulating polyandry in Lobesia bo-trana (Lepidoptera: Tortricidae). Physiological Entomolo-g y, 22(4), 387-393.
- Vacari, A.M., De Bortoli, S.A., Borba, D.F. & Martins, M.I.E.G. (2012) Quality of Cotesia flavipes (Hymenoptera: Braconi-dae) reared at different host densities and the estimated cost of its commercial production. Biological Control, 63(2), 102-106.
- Walker, D.W. (1965) Bionomics of the sugarcane borer Diatra-ea saccharalis (Fab.). A description of the mating behav-ior. Proceeding of the Entomological Society of Washington, 67(2), 80-83.
- Wedell, N., Wiklund, C. & Cook, P.A. (2002) Monoandry and polyandry as alternative lifestyles in a butterfly. Behavioral Ecology, 13(4), 450-455.
- Willink, E. (1982) Especies de Diatraea en cañaverales del Noroeste Argentino. Estación Experimental Agroindustrial Obispo Colombres, Tucumán, Boletín 140, 1-38.

