










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
versión impresa ISSN 0373-5680versión On-line ISSN 1851-7471
Rev. Soc. Entomol. Argent. vol.76 no.1-2 La Plata jun. 2017
Trabajo Científico
Toxicidad sobre Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) de la proteína Bt expresada en hojas de diferentes eventos de maíz transgénico liberados en Paraguay
Toxicity of the Bt protein expressed in leaves of different events of transgenic corn released in Paraguay against Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae)
GÓMEZ, Víctor A., VILLALBA, Giselle E., ARIAS, Osmar R., RAMÍREZ, María B. & GAONA, Edgar F.
Facultad de Ciencias Agrarias, Universidad Nacional de Asunción (FCA-UNA). E-mail: victor.gomez@agr.una.py
Received 05 - Vil - 2016 | Accepted 14 - II - 2017 | Published 29 - VI - 2017 https://doi.org/10.25085/rsea.761201
RESUMEN. El maíz transgénico que expresa la proteína Bt es una de las estrategias más utilizadas para el control de Spodoptera frugiperda, plaga clave de este cultivo en América. Con el objetivo de verificar la susceptibilidad de S. frugiperda, se realizaron estudios de toxicidad de las proteínas Bt expresadas en hojas de diferentes eventos de maíces liberados en Paraguay, 2B587HXT (Cry1F); VT Triple PROT (Cry1A/Cry2Ab/Cry3Bb) y Fórmula TLT (Cry1Ab). La cría de S. frugiperda se realizó en laboratorio para obtener una población uniforme bajo condiciones controladas de temperatura (25 ± 2 °C), humedad relativa (60 ± 10%) y fotoperíodo de (14:10 h L:O ). Se evaluó la mortalidad y, en el caso de los sobrevivientes, el peso y longitud de las pupas, la fecundidad de las hembras, la longevidad de los adultos, la viabilidad de los huevos, y la duración de los diferentes estadios y del ciclo total. La alimentación de larvas neonatas con hojas del evento VT Triple PROT produjo 100% de mortalidad; sin embargo, en las larvas alimentadas con hojas de 2B587HXT o Fórmula TLT, la mortalidad fue 58 y 56%, respectivamente. Los sobrevivientes a estos tratamientos presentaron un ciclo de vida más largo, y fecundidad y fertilidad reducidas.
PALABRAS CLAVE. Maíz Genéticamente Modificado. Biología. Plaga.
ABSTRACT. The transgenic maize that expresses the Bt protein is one of the strategies most used for the control of Spodoptera frugiperda, key pests of this crop in America. With the objective of verifying the susceptibility of S. frugiperda; toxicity studies were conducted of the proteins Bt expressed in leaves of different events of maize released in Paraguay, 2B587hx™ (cry1F); VT triple PRO™ (Cry1A/Cry2Ab/Cry3Bb) and Formula TL™ (Cry1Ab). The rearing of S. frugiperdawas conducted in the laboratory to obtain a uniform population under controlled conditions of temperature (25 ± 2 °C), relative humidity (60 ± 10%) and photoperiod of 14:10 h (L:D). Mortality was evaluated and in the case of survivors; the weight and length of the pupae, the fertility of the females, the longevity of the adults, the viability of eggs, and the duration of the different stages and the total cycle. The supply of larvae neonates in the leaves of maize of the event VT triple PRO™ produced 100% mortality; however, in the larvae fed with leaves of the events 2B587hx™ and Formula TL™; the mortality was 58% and 56%, respectively. The survivors to these treatments had a longer life cycle, fertility and reduced fertility.
KEYWORDS. Genetically modified maize. Biology. Pest.
INTRODUCCIÓN
En Paraguay, el cultivo de maíz es de mucha importancia socioeconómica; sus granos son utilizados para la alimentación humana y animal, y en la mayoría de los casos constituye una fuente de ingresos para los productores. En 2015 se sembraron unas 838.768 ha, con un rendimiento promedio de 6.045 kg/ha (CAPECO, 2016). Debido a su gran utilidad y atendiendo ciertas ventajas en el control de plagas, en 2013 se liberaron en Paraguay los primeros eventos de maíz transgénicos que expresan la proteína de Bacillus thuringiensis(Bt), llegando a ocho el número de eventos oficialmente liberados hasta la fecha (SENAVE, 2016). El maíz convencional sembrado en Paraguay sufre serios daños a causa deSpodoptera frugiperda(Smith) (Lepidoptera: Noctuidae), una plaga de difícil control (Ruiz et al. , 2004). Esta especie es también una importante plaga en los vecinos países de Argentina y Brasil, donde ocasiona pérdidas significativas a la producción (Fernandes et al. , 2003; Murúa & Virla, 2004).
En Estados Unidos, a fines de la década de los '90, se evaluaron los primeros híbridos transgénicos Bt11 y MON810 (Yield GardT), para el control deS. frugiperda, con los que se lograron importantes reducciones en la infestación del cogollo y las espigas (Buntin et al. , 2001). El uso de esta tecnología aumentó a través de los años. En 2015, el 29% (53.600.000 ha) del maíz sembrado en el mundo estaba genéticamente modificado. Estados Unidos, Brasil y Argentina son los países con mayores superficies sembradas con estos cultivos (ISAAA, 2016).
La eficacia del maíz Bt para controlar a S. frugiperda se evaluó en diferentes países, y existen reportes de éxitos y fracasos (Williams et al., 1997; Waquil et al. , 2002; Zenner et al., 2009; Giaveno et al., 2010; Flores & Balbi, 2014; Grandis de Lima & Assmann 2015; Monnerat et al., 2015). Por otro lado, en Puerto Rico, Brasil y el sur de Estados Unidos se detectó resistencia d e S. frugiperdaa maíces Bt que expresan las proteínas Cry1F o Cry1Ab (Storer et al., 2010; Farias et al., 2014; Huang et al. , 2014; Omoto et al., 2016).
En Paraguay, los productores han detectado daños producidos por S. frugiperda en maíces Bt. Huang et al., (2011) mencionan que pueden existir variabilidad de expresión de la proteína entre variedades y en diferentes estructuras de las plantas, exponiendo a la plaga a dosis subletales. Por esta razón, para encarar estrategias pro-activas de manejo de resistencia dentro de un programa de manejo integrado de plagas (MIP), para prolongar la vida útil de estas tecnologías en campo, es de fundamental importancia evaluar la eficacia de los eventos liberados en el Paraguay sobre S. frugiperda.
Por lo mencionado anteriormente, este trabajo tiene como objetivo general evaluar en larvas deS. frugiperda la toxicidad de diferentes eventos de maíz Bt sembrados en Paraguay, y en particular, registrar el porcentaje de mortalidad y, en los sobrevivientes, determinar la duración de los estadios de larva, prepupa y pupa, el peso y longitud de las pupas, la fecundidad y longevidad de los adultos, la viabilidad de los huevos y el ciclo total del insecto.
MATERIALES Y MÉTODOS
Recolección y cría de S. frugiperda
Se realizaron colectas manuales de larvas de S. frugiperda en cultivos de maíz no Bt, en puntos estratégicos que corresponden a la región de mayor producción de este rubro en Paraguay, ubicados en el Distrito de Pirapó, Departamento de Itapúa, Paraguay (latitud: 26° 51' 36", longitud: 55° 32' 42"). Las larvas fueron colocadas en recipientes plásticos con tapas ndividuales que contenían dieta artificial (Parra, 2001 Da Silva & Parra, 2013), y trasladadas al laboratorio de cría de insectos antes de pasar las 48 h. Una vez en el laboratorio, las larvas fueron transferidas a otros vasos de plástico de 50 ml que contenían dieta artificial y ubicadas en salas de cría en condiciones controladas de temperatura (25 ± 5 °C), humedad relativa (60 ± 10%), y fotoperíodo (14:10 horas L: O), hasta el estadio de pupa. Las pupas fueron retiradas y desinfestadas con hipoclorito de sodio al 0,2% en agua destilada para uego ser colocadas sobre papel de filtro en platos plásticos cubiertos con vasos transparentes de 250 cm3. Los adultos emergidos (diez machos y diez hembras) fueron trasladados a jaulas de apareamiento que consistían en tubos de PVC de color blanco, de 24 cm de altura y 14,5 cm de diámetro, revestidos nternamente con papel blanco como sustrato de oviposición. Las jaulas fueron ubicadas en platos plásticos, sobre papel de filtro, y tapadas en la parte superior con telas tipo tul sujetadas con gomitas para favorecer la aireación. Los adultos fueron alimentados con una solución de agua y miel de abeja al 10% presentada en trozos de algodón, ubicados en la parte superior de la jaula en contenedores de 1,5 cm de longitud y 2 cm de diámetro. Se procedió a retirar los huevos cada 48 h y se los acondicionó en vasos de plásticos de 50 ml con un trozo de papel de filtro humedecido en la parte superior para evitar la desecación. Los vasos fueron mantenidos en cámaras climatizadas tipo B.O.D (temperatura 24 ± 2 ºC;) humedad relativa 60 ± 10%, y fotoperíodo 14:10 h L:O Una parte de la primera generación de larvas eclosionadas (F1) fue usada para el trabajo experimental (larvas neonatas con máximo de 24 h de edad) y otra parte se separó para el mantenimiento de la población en el laboratorio.
Material vegetal
Para obtener las hojas de los diversos eventos de maíz Bt se realizaron siembras en invernadero. El evento 2B587HXT que expresa la proteína Cry1F fue sembrado en abril. El material VT Triple PROT con las proteínas Cry1A.105/Cry2Ab2/Cry3Bb en junio, y el maíz Bt Fórmula TLT que expresa la proteína Cry1Ab con el testigo híbrido convencional 2B587T en el mes de setiembre (Tabla I). Las hojas de los diferentes eventos de maíz utilizadas en los experimentos se obtuvieron de plantas de 30 días contados desde la emergencia, las cuales fueron cortadas al momento de ser utilizadas con sacabocados de 4,5 cm de diámetro.
Tabla I. Eventos de maíz transgénico con los que fueron alimentadas las larvas d e S. frugiperda

Bioensayos
Determinación de mortalidad
Los experimentos se realizaron en vasos plásticos de 50 ml que, para mantener la humedad, contenían en su base una mezcla de agar-agua al 2% cubierta con papel de filtro. En cada vaso se colocó sobre el papel un círculo de 4,5 cm de hoja de maíz y luego se depositaron sobre él ocho larvas neonatas. Inicialmente, los círculos de hojas fueron reemplazados cada dos días, pero a partir del tercer estadio de desarrollo larval fueron reemplazados en forma diaria. Cada tratamiento se repitió doce veces, totalizando 96 larvas por tratamiento. Se realizaron observaciones diarias para registrar la mortalidad de las larvas. Para el cálculo del porcentaje de mortalidad se tuvo en cuenta la diferencia con el total de individuos que no completaron su ciclo, es decir que no llegaban al estadio adulto.
Duración de los estadios larval y prepupal
Se realizaron observaciones diarias para registrar los cambios de estadio de las larvas sobrevivientes. Para la determinación de la duración de cada estadio, fueron utilizados como referencia aquellos individuos que lograban completar el estadio; es decir, aquellos que conseguían mudar de larva a prepupa; y de prepupa a pupa.
Duración del estadio de pupa, y peso y longitud por sexo
Para la determinación de duración del estadio pupal se tuvo en cuenta desde el primer día de formación de la misma hasta la emergencia del adulto, teniendo siempre en cuenta solo los individuos que lograban completar este estadio. Veinticuatro horas después de empupar, se determinó el sexo, el peso y la longitud de cada pupa. El peso se determinó con una balanza de precisión (RADWAG WTB 200 Exactitud 0,001 g), y la longitud (mm) con una regla graduada mediante un microscopio estereoscopio (Labo America, Inc, USA). Posteriormente las pupas fueron colocadas en bandejas con papel de filtro humedecido en la parte inferior y tapadas con vasitos plásticos, identificados con su respectiva numeración y tratamiento. Se mantuvieron así hasta la emergencia de los adultos.
Relación de sexos y porcentaje de emergencia
Para la relación de sexos y el porcentaje de emergencia se utilizaron las siguientes fórmulas (García & Iannacone, 2011);
%H = N°H x 100/N°AE
%M = N°M x 100/N°AE
Donde: %H = Porcentaje de hembras.; %M = Porcentaje de machos.; NºH = Número de hembras.; NºM = Número de machos.; NºAE = Número de adultos emergidos.
%E = T - (PnE + AmE/T)
Donde %E = Porcentaje de emergencia.; T: Número total de pupas.; PnE = Pupas no emergidas.; AmE = Adultos emergidos.
Periodos de pre-oviposición, oviposición, postoviposición (días), fecundidad de hembras y viabilidad de huevos
De los adultos emergidos, se seleccionaron diez parejas para observaciones del periodo de pre-oviposición, oviposición, post-oviposición, fecundidad de hembras y viabilidad de huevos; los adultos restantes fueron colocados individualmente en tubos PVC y alimentados con miel al 10% hasta su muerte.
Los parámetros evaluados en este estadio fueron el período de preoviposición, oviposición y postoviposición (días), el número de huevos por hembra (fecundidad) y la fertilidad (viabilidad), el número de posturas, y la longevidad (días) de los adultos.
Para cuantificar la fecundidad se utilizó la formula (García & Iannacone, 2011).
Fecundidad = N°HV/N°vs
Dónde: Nº HV = Número de hembras; NºHvs = Número de huevos puestos.
La fertilidad o viabilidad de acuerdo a lo señalado por Hernández et al.(2010).
Fertilidad = Hvs/N°N
Dónde: NºHvs = Número de huevos puestos; Nº N = Número neonatas
Duración del ciclo de vida
Para la determinación de esta variable fueron contabilizados los días transcurridos en los insectos sobrevivientes para completar todos los estadios de larva a adulto, en cada tratamiento.
Análisis estadístico
Los resultados de cada uno de los parámetros fueron sometidos a un análisis de varianza (ANOVA), y la comparación de medias de los tratamientos se realizó a través del test de Tukey (a = 5%), mediante el programa estadístico INFOSTAT-2014 (Di Rienzo et al., 2014)
RESULTADOS Y DISCUSIÓN
Mortalidad d e S. frugiperdae n maíces Bt
Los porcentajes de mortalidad entre los días 7 y 63 después del inicio de la alimentación con hojas de maíces Bt y el híbrido convencional se presentan en la Tabla II. Comparado con los maíces que expresan la proteína Bt, el maíz convencional 2B587T, produjo un bajo porcentaje de mortalidad. En el día 7, los eventos 2B587HXT (Cry1F) y Fórmula TLT(Cry1Ab) produjeron una mortalidad del 30 y del 15% con diferencias estadísticas (F = 96,5; gl = 33; P = 0,0001), respectivamente, siendo estos resultados menores al obtenido por Zenner et al.(2009): 61,7% de mortalidad a los siete días alimentando larvas con maíz Bt (Cry1Ab). Las mortalidades en los grupos expuestos a las proteínas Cry1F (2B587HXT) y Cry1Ab (Fórmula TLT) no presentaron diferencias significativas a partir de los 14 días llegando a un máximo de 58 y 56%, respectivamente. El evento VT Triple PROT con las proteínas Cry1A.105/Cry2Ab2/Cry3Bb produjo un 100% de mortalidad a los 21 días diferenciándose estadísticamente de los demás eventos (F = 151,1; gl = 33; P = 0,0001). Se ha reportado que híbridos que expresan la proteína Cry1F son los más resistentes al daño de S. frugiperda; seguidos por los híbridos con la proteína Cry1Ab (Waquil et al., 2002). Vilella et al. (2002) reportaron que después de cuatro generaciones en condiciones de laboratorio, la alimentación de S. frugiperdacon maíz con la proteína Cry1Ab aumentó la supervivencia.
Fallas en el control de S. frugiperda fueron mencionadas por Huang et al. (2011), Tabashnik et al. (2013), y Sumerford et al.(2013). Giaveno et al.(2010) obtuvieron un 41% de mortalidad con proteínas Cry1Ab y 39% con proteínas Cry1F. En Paraguay no existen estudios poblacionales sobre la susceptibilidad de S. frugiperdaa los maíces Bt comerciales. Los resultados de la presente investigación son importantes debido a la valoración de la toxicidad de los eventos de maíz Bt en condiciones controladas.
Tabashnik et al. (2013) afirmaron que en un mismo evento de más de una proteína de Bt pertenecientes a diferentes grupos y la adopción de áreas de refugio se constituyen en estrategias para retrasar la evolución de la resistencia en campo. En el presente trabajo se pudo verificar que el evento VT Triple PROT, que expresa más de una proteína Bt, ocasionó la mortalidad total en larvas d eS. frugiperda.
Duración y porcentaje de los estadios larval y prepupal de los insectos sobrevivientes a los maíces Bt y convencional
La Tabla III presenta la duración de los estadios larval y prepupal. En el caso de las hembras, las expuestas al evento Formula TLT presentaron un estadio larval de 21,03 días, significativamente mayor que la de los otros grupos (F = 65,86; gl = 41; P = 0,0001). Las hembras alimentadas sobre el evento 2B587HXT presentaron un estadio pupal de 2,92 días, significativamente mayor que los demás tratamientos (F = 13,19; gl = 41; P = 0,0001). En el caso de los machos, la duración de la etapa larval fue significativamente menor en los individuos alimentados sobre el evento 2B587HXT (F = 65,86; gl = 41; P = 0,0001), mientras que en el estadio prepupal este evento produjo una duración significativamente mayor que los otros tratamientos (F = 13,19; gl = 41; P = 0,0001). En todos los casos, la duración del estadio larval fue inferior a lo mencionado por Murúa et al. (2008), quienes afirman que el estadio larval de S. frugiperda dura aproximadamente 25 días cuando se alimenta con hojas de maíz convencional. Giolo et al. (2002) obtuvieron una duración media de 2 días en el estadio prepupal, alimentando a las larvas d e S. frugiperdacon hojas de maíz convencional. Este resultado fue superior al obtenido en la presente investigación, excepto con el material Bt 2B587HXT que ocasionó una mayor duración en el estadio prepupal.
Longitud, peso y duración de pupas por sexo
La Tabla IV muestra la longitud, el peso y la duración del estadio de pupa. Tanto los machos como las hembras presentaron diferencias significativas que dependieron de los tratamientos. Las hembras que presentaron el mayor tamaño fueron las alimentadas con el evento Formula TLT (F = 266,5; gl = 35; P = 0,0001). Con respecto al peso de las pupas, alimentadas con maíz convencional presentaron un peso significativamente mayor que los producidos por el evento 2B587HXT (F = 40,6; gl = 51; P = 0,0001). En este sentido, Zenner et al. (2009) encontraron que las pupas originadas de larvas alimentadas con maíz que expresa la toxina Cry1Ab presentaron menor peso que las alimentadas con maíz convencional.
Todas las pupas provenientes de las larvas alimentadas con maíces Bt presentaron diferencias significativas (F = 346,4; gl = 41; P = 0,0001) entre sí en cuanto a la duración, siendo las provenientes de la alimentación con el evento 2B587HXT las que más permanecieron en ese estadio. Las toxinas ingeridas por los individuos de S. frugiperda alimentados con maíces Bt pueden causar un efecto adverso en su biología, arrojando como resultado un aumento en la duración de los estadios del ciclo de vida. Zenner et al. (2005) demostraron en larvas de S. frugiperda que la alimentación con hojas de maíz que expresa la proteína Cry1Ac aumenta la duración del estadio pupal, tanto en machos (10,88 días) como en hembras (11,60 días).
Tabla II. Porcentaje de mortalidad de larvas S. frugiperdaalimentada sobre distintos eventos de maíz Bt.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Relación de sexos y porcentaje de emergencia de adultos
Del total de pupas de cada tratamiento, la relación macho/hembra no presentó diferencias estadísticas atribuibles a los tratamientos (Tabla V). Zenner et al. , (2005) encontraron una relación de 1:1,36 en larvas alimentadas con proteína Cry1Ac.
Por otro lado, se observó que los individuos alimentados con maíz Bt presentaron deformaciones en el desarrollo de las alas, seguidas de mortalidad. Esto produjo una reducción del 11% en el porcentaje de emergencia de adulto alimentados con el evento Fórmula TLT y del 22% en los alimentados con el evento 2B587HXT, respecto al porcentaje de emergencia de individuos alimentados con el maíz convencional 2B587.
Periodo de pre-oviposición, oviposición, postoviposición, fecundidad de las hembras y viabilidad de los huevos
No se observaron diferencias significativas en cuanto a la duración de los periodos de pre-oviposición, oviposición y post-oviposición (Tabla VI). Sin embargo, el período de pre-oviposición fue menor en los insectos alimentados con maíz convencional; además, estos individuos presentaron mayores períodos de oviposición y de post-oviposición, en comparación con alimentados con maíz Bt. Estos resultados podrían revelar una posible antibiosis por efecto de la toxina Bt. Alimentando larvas de S. frugiperda con diferentes genotipos de maíz convencional, Machado et al. (2004) obtuvieron una duración de 3,94 ± 0,584 días en la pre-oviposición; una oviposición de 6,11 ± 0,469 días y post-oviposición de 5,50 ± 1,348 días; datos superiores a los obtenidos en esta investigación.
Tabla III. Duración del estadio larval y prepupal de hembras y machos de S. frugiperdacuyas larvas fueron alimentadas sobre distintos eventos de maíz Bt.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Tabla IV. Longitud, peso y duración de la etapa pupal de S. frugiperdacuyas larvas fueron alimentadas sobre distintos eventos de maíz Bt.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Tabla V. Porcentaje de emergencia y proporción de machos y hembras de S. frugiperdacuyas larvas fueron alimentadas sobre distintos eventos de maíz Bt
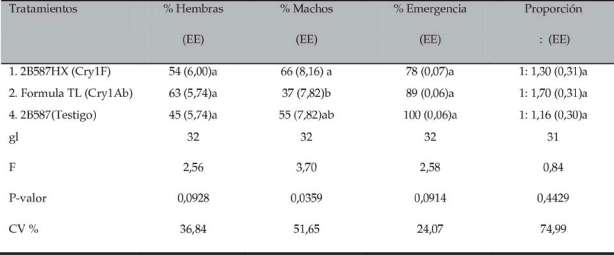
CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Los insectos alimentados con el maíz híbrido convencional 2B587T depositaron mayor cantidad de huevos 605,3 (EE = 91,88), y el porcentaje de viabilidad fue mayor que los valores correspondientes a los individuos alimentados con maíz Bt (Tabla VII). La menor fecundidad 146,6 (EE = 91,88) se registró en los individuos que recibieron el tratamiento con el evento Fórmula TLT, aunque su fertilidad fue mayor que la correspondiente al evento 2B587HXT. Estos resultados indican que la alimentación con hojas de los eventos Bt afectó la fecundidad de las hembras y la viabilidad de los huevos.
Los insectos alimentados con el híbrido convencional presentaron el mayor número de posturas 2,40 (EE = 0,34) (Tabla VIII). El menor número de posturas 0,70 (EE = 0,34) correspondió a los insectos alimentados con el evento Fórmula TLT. Zenner et al., (2005) obtuvieron 16 posturas en individuos alimentados con algodón transgénico (proteína Cry1Ab). En la presente investigación; insectos alimentados con los eventos Bt fueron afectados en el número de posturas con relación a los insectos alimentados con el maíz híbrido convencional.
Tabla VI. Periodos de pre- oviposición, oviposición y post- oviposición en parejas de S. frugiperdacuyas larvas fueron alimentadas sobre distintos eventos de maíz Bt.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Duración de los diferentes estadios de S. frugiperda
En la Tabla IX se detalla la duración de los estadios d e S. frugiperdaalimentada con diferentes eventos de maíz Bt y no Bt. Los individuos alimentados con maíz convencional fueron más precoces que los alimentados con maíz Bt. Un resultado similar fue obtenido por Cássio, (2002) quien registró una etapa larval en promedio de 18,75 días en S. frugiperda alimentadas con hojas de maíz. Zenner et al., (2009) observaron una mayor duración de la etapa larval en individuos alimentados con eventos transgénicos, comparados con un testigo sin la proteína Bt. En este trabajo; larvas d e S. frugiperda alimentadas con Fórmula TLT presentaron mayor duración del estadio con relación al híbrido convencional. Mendes et al., (2011) en estudios con S. frugiperda concluyeron que la etapa larval es mayor en los insectos alimentados con maíz Bt.
Estos resultados demuestran que existe una diferencia en la toxicidad de las proteínas de los eventos de maíz Bt sobre S. frugiperda, evidenciada en los aspectos biológicos estudiados de la plaga, y, en este caso específico, en su capacidad reproductiva (menor fecundidad y fertilidad), que deriva a una disminución de descendientes.
Resultados preliminares en laboratorio nos llevan a mencionar que se detectaron diferencias en el control de S. frugiperda entre los maíces Bt utilizados por los productores en Paraguay. Se deberá sumar mayor número de investigaciones relacionadas para verificar la eficacia de los materiales a efectos de obtener informaciones importantes para el manejo adecuado de los maíces Bt liberados en el Paraguay.
Tabla VII. Medias de número de huevos (fecundidad), números de neonatas obtenidas y fertilidad o viabilidad de huevos en diez casales.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Tabla VIII. Total de posturas obtenidas en 10 casales instalados de larvas alimentadas con maíces Bt y maíz convencional en condiciones de laboratorio.
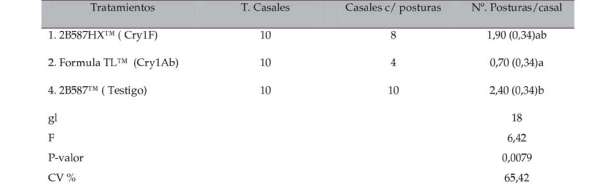
CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
Tabla IX . Medias de duración (días) de estadios de S. frugiperdaalimentados con maíces Bt y convencional bajo condiciones de laboratorio.

CV = Coeficiente de Variación. EE = error estándar. En cada columna, diferentes letras denotan diferencias significativas detectadas por el Test de Tukey P < 0,05.
CONCLUSIONES
En base a los resultados obtenidos y en las condiciones en las que se realizó el estudio, se concluye lo siguiente: (a) se observó un 100% de mortalidad en los insectos que durante 21 días se alimentaron sobre el evento VT Triple PROT, que expresa las proteínas Cry1A.105/Cry2Ab2/Cry3Bb; (b) los eventos 2B587HXT (Cry1F) y Fórmula TLT
BIBLIOGRAFÍA CITADA
Buntin, G.D., Lee, R.D., Wilson, D.M., & McPherson, R.M. (2001) Evaluation of Yield Gard transgenic resistance for control of fall armyworm and corn earworm (Lepidoptera: Noctuidae) on corn. Florida Entomologist, 84(1), 37-42. [ Links ]
CAPECO (Cámara Paraguaya de Exportadores y Comercializadores de Cereales y Oleaginosas) (2016) Área de siembra, producción y rendimiento: estadística soja (en línea). Paraguay. Cons: 4-VII-2016. Disponible en http://www.capeco.org.py/estadisticas.php [ Links ]
Cássio, R. (2002) Danos e biología de Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae) em genotipos de milho. Brasil/ESALQ. (en línea). Cons: 4-11-2016 Disponible en http://repository.usp.br/result.php?assunto=CRUZAMENTO%20VEGETAL&search %5B%5D=+authorUSP.keyword:%22SILOTO,%20ROMILD 0%20C%C3%81SSIO%22. [ Links ]
Da Silva, C.S.B., & Parra, J.R.P. (2013) New method for rearing Spodoptera frugiperda in laboratory shows that larval cannibalism is not obligatory. Revista Brasileira de Entomologia, 57(3), 347-349. [ Links ]
Di Rienzo, J.A., Casanoves, E, Balzarini, M.G., González, L, Tablada, M., & Robledo, C.W. (2014) InfoStat versión 2014 Grupo InfoStat, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. AR. Disponible http://www.infostat.com.ar [ Links ]
Farias, J.R., Andow, D.A., Horikoshi, R.J., Sorgatto, R., Fresia, R, Dos Santos, A.C., & Omoto, C. (2014) Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Protection, 64, 150-158. [ Links ]
Fernandes, O.D., Parra, J.R.P, Neto, A.F, Pícolr, I., Borgatto, A.F, & Demétrio, C.G.B. (2003) Efeito do milho genéticamente modificado mon810 sobre a Lagarta-do-cartucho Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae). Revista Brasileira de Milho e Sorgo, 2(2), 25-35. [ Links ]
Flores, F, & Balbi, E. (2014) Evaluación del daño de oruga militar (Spodoptera frugiperda) en diferentes híbridos comerciales de maíz transgénicos. (en línea). Argentina. Cons: 6-11-2016. Disponible en http://inta.gob.ar/sites/ default/files/script-tmp- inta_plagas_maiz_mj_14.pdf [ Links ]
García, J., & lannacone, J. (2011) Viabilidad de huevos y modelo de jaula para la cría artificial masiva de Anastrepha fraterculus (Díptera, Tephritidae). Revista de la Sociedad Entomológica Argentina, 70(3-4), 267-276. [ Links ]
Giaveno, C.D., Paravano, A.S., Celia, M., & Curis, M.C. (2010) Efecto de diferentes maíces Bt en la biología de Spodoptera frugiperda (J.E. Smith) (Lepidoptera, Noctuidae). Revista FAVE, Ciencias Agrarias, 9(1-2), 45-54 [ Links ]
Giolo, F, Grützmacher, A., Garcia, M., & Busato, G. (2002) Parámetros biológicos de Spodoptera frugiperda (J.E. Smith, 1797) (Lepidoptera: Noctuidae) oriundas de diferentes localidades e hospedeiros. Revista Brasileira Agrociéncia, 8(3), 219-224. [ Links ]
Grandis De Lima, L, & Assmann, A.S. (2015) Desfolha causada pela Spodoptera frugiperda em milho com diferentes biotecnologías. Brasil. Cultivando O Saber, Ed. especial., 56-66. [ Links ]
Hernández, E., Rivera, J.P, Orozco-Dávila, D., Salvador, M., & Toledo, J. (2010) An artificial larval diet for rearing of Anastrepha striata (Díptera: Tephritidae). Folia Entomológica Mexicana, 93, 167-174. [ Links ]
Huang, F, Andow, D.A., & Buschman, L.L. (2011) Success of the high dose/refuge resistance management strategy after 15 years of Bt crop use in North América. Entomologia Experimentalis et Applicata,\AO, 1-16. [ Links ]
Huang, F, Qureshi, J.A., Meagher, R.L. Jr., Reisig, D.D., Head, G.P, Andow, D.A., Ni X., Kerns, D., Buntin, G.D., Niu, Y, Yang, F, & Dangal, V. (2014) CrylF resistance in fall armyworm Spodoptera frugiperda: single gene versus pyramided Bt maize. PlosOne, 9(11), 1-10. [ Links ]
ISAAA (International Service for the Acquisition of Agri-biotech Applications) (2016) Biotech crop highlights in 2015. Pocked Nº 16. Ithaca, NY. (en línea). USA. Cons: 6-III-2016. Disponible en: http://www.isaaa.org/resourses/publications/ pocketk/16/default.asp
Machado, L., Rodrigues, L., Guedes, L.M., & Stoffel, C.F. (2004) Fertilidade e longevidade de Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) em genótipos de milho. (en línea). Brasil. Cons: 4-III-2016. Disponible en: http://dx.doi.org/10.1590/S0103-84782004000200002
Mendes, S., Brasil K., Lopes, M., Waquil, M., & Waquil, J. (2011) Respostas da lagarta-do-cartucho a milho geneticamente modificado expressando a toxina Cry 1A(b). BR. Pesquisa Agropecuaria, 46(3), 239-244.
Monnerat, R., Martins, E., Macedo, C., Queiroz, P., Praça, L., Soares, M., Moreira, H., Grisi, I., Silva, J., Soberon, M., & Bravo, A. (2015) Evidence of field-evolved resistance of Spodoptera frugiperdato Bt corn expressing Cry1F in Brazil that is still sensitive to modified Bt toxins. University of Tennessee, United States. PlosOne, 10(4)1-12.
Murúa, M.G., & Virla, E.G. (2004) Presencia invernal de Spodoptera frugiperda(Smith) (Lepidoptera: Noctuidae) en el área maicera de la Provincia de Tucumán, Argentina. Revista de la Facultad de Agronomía, 105(2), 46-52.
Murúa, M.G., Vera, M.T., Abraham, S., Juárez, M.L., Prieto, S., Head, G.P., & Willink, E. (2008) Fitness and mating compatibility of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) populations from different host plant species and regions in Argentina. Annals of the Entomological Society of America, 101(3), 639-649.
Omoto, C., Bernardi, O., Salmeron, E., Sorgatto, R., Dourado, P., Crivellari, A., Carvalho, R., Willse, A., Martinelli, S., & Head, G. (2016) Field-evolved resistance to Cry1Ab maize by Spodoptera frugiperda in Brazil. Pest Management Science, 9, 1727-1736.
Parra, J.R.P. (2001) Técnicas de criação de insetos para programas de controle biológico.FEALQ, Piracicaba. Brasil.
Ruiz, E.M., Cabral, C.C., & Pino, C.D. (2004) Eficiencia de Bacillus thuringiensis Linea HD-1 en el control de Spodoptera frugiperda (Lepidoptera: Noctuidae) en condiciones de campo y de laboratorio en el cultivo de maíz dulce Zea mays saccharata. Investigación Agraria, 6(1), 10-14.
SENAVE (Servicio Nacional de Calidad y Sanidad Vegetal y de Semillas) (2016) Maíz. Listado de eventos con modificación genética liberados en el país. (en línea). Paraguay. Cons: 4-IV-2016. Paraguay. Disponible en http://www.senave.gov.py/docs/servicios/ bioseguridadAgricola/
Storer, N.P., Babcock J.M., Schlenz M., Meade T., Thompson G.D., Bing J.W., & Huckaba R.M. (2010) Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda(Lepidoptera: Noctuidae) in Puerto Rico. Journal of Economic Entomology, 103(4), 1031-1038.
Sumerford, D.V, Head, G.P., Shelton, A., Greenplate, J., & Moar, W. (2013) Field evolved resistance: assessing the problem and ways to move forward. Journal of Economic Entomology, 106(4), 1525-1534.
Tabashnik, B.E., Brevault, T., & Carriere, Y. (2013) Insect resistance to Bt crops: lessons from the first billion acres. Nature Biotechnology, 31, 510-521.
Vilella, F.M., Waquil, J.M., Vilela, E.F., Siegfired, B.D., & Foster, J.E. (2002) Selection of the fall armyworm Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) for survival on Cry1A(b) Bt toxin. Revista Brasileira de Milho e Sorgo, 1(3), 12-17.
Waquil, J.M., Villela, F.M., & Foster, J.E. (2002) Resistencia do milho (Zea mays L.) transgénico a lagarta-do-cartucho, Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae). Revista Brasileira de Milho e Sorgo, 1(3), 1-11.
Williams, W.P., Sagers, J.B., Hanten, J.A., Davis, F.M.,& Buckley, P.M. (1997) Transgenic corn evaluated for resistance to fall armyworm and southwestern corn borer. Crop Science, 37(3), 957-962.
Zenner, D.P.I., Álvarez, J.A., Mejía, C., & Bayona, M.A. (2005) Influencia de la toxina Cry1Ac del Bacillus thuringiensis sobre el desarrollo del cogollero del maíz, Spodoptera frugiperda (J. E. Smith). Revista U.D.C.A Actualidad& Divulgación Científica, 8(2), 129-139.
Zenner, I.D.P., Arévalo, H., Mejía, C., & Díaz, J. (2009) Spodoptera frugiperda: respuesta de distintas poblaciones a la toxina Cry1Ab. Revista Colombiana de Entomología, 34(1), 41-50.

