










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de la Sociedad Entomológica Argentina
Print version ISSN 0373-5680On-line version ISSN 1851-7471
Rev. Soc. Entomol. Argent. vol.76 no.3-4 La Plata Dec. 2017
Trabajo científico - Article
Tabla de vida y parámetros poblacionales de Lobesia botrana (Den. et Schiff.) en condiciones de laboratorio
Life table and population parameters of Lobesia botrana (Den. et Schiff.) under laboratory conditions
HERRERA, María E., DAGATTI, Carla V. & BECERRA, Violeta C.
Estación Experimental Agropecuaria Mendoza INTA. San Martín 3853, C.P. 5507, Luján de Cuyo, Mendoza, Argentina. Email: herrera.mariae@inta.gob.ar
Received 11 - V - 2017 | Accepted 04 - X - 2017 | Published 27 - XII - 2017 https://doi.org/10.25085/rsea.763404
RESUMEN. Lobesia botrana Den. & Schiff. es plaga cuarentenaria para Argentina. Su reciente introducción en la provincia de Mendoza requiere el estudio de aspectos relacionados al ciclo de vida y parámetros poblacionales. El objetivo de este trabajo fue construir una tabla de vida horizontal, determinar los parámetros demográficos, la duración media de los períodos pre-reproductivo, reproductivo, post-reproductivo, la longevidad adulta, la fecundidad y fertilidad de las hembras bajo condiciones de laboratorio. Los parámetros demográficos de L. botrana obtenidos fueron: tasa intrínseca de crecimiento poblacional (rm) de 0,137, tasa neta de reproducción (Ro) de 66,82, tiempo generacional (T) de 30,51, tasa de incremento finito (λ) de 1,073 y el tiempo de duplicación (D) de 9,76 días. La fecundidad bruta obtenida (Mx) de 134,84 ± 15,68 huevos por hembra, la duración media en días de los períodos pre-reproductivo fue de 1 ± 0,81, reproductivo 6,25 ± 1,25 y post-reproductivo de 2 ± 0,68, la longevidad de la hembra 9,25 ± 1,67 y del macho 8,64 ± 0,83 días.
PALABRAS CLAVE. Estudios biológicos. Polilla de la vid. Vitis vini fera .
ABSTRACT. Lobesia botrana Den. & Schiff. is a quarantine pest for Argentina. Its recent introduction in Mendoza requires studies of life cycle and population parameters. The aim of this work is to construct a horizontal life table, determine the demographic parameters, the average duration of the pre - reproductive, reproductive, post - reproductive periods, adult longevity, fecundity and fertility under laboratory conditions. The demographic parameters of L. botrana obtained were: intrinsic rate of increase (rm) of 0.137, net reproductive rate (Ro) of 66.82, mean generation time (T) of 30.51, finite rate of increase (λ) of 1.073, and the doubling time (D) of 9.76 days. The gross fecundity obtained (Mx) of 134.84 ± 15.68 eggs per female, the mean duration in days of pre-reproductive periods was 1 ± 0.81, the reproductive 6.25 ± 1.25 and the post-reproductive period of 2 ± 0.68, female longevity 9.25 ± 1.67 and male 8.64 ± 0.83 days.
KEYWORDS. Biological studies. European grapevine moth. Vitis vini fera .
INTRODUCCIÓN
Lobesia botrana Den. & Schiff., un microlepidóptero de la familia T ortricidae, es la principal plaga de los viñedos europeos. En el año 2008 se establece en Chile, durante 2009 llega a
California y en octubre de 2010 el Servicio Nacional de Sanidad Agropecuaria declara en Argentina la emergencia ftosanitaria (SENASA, 2010). La introducción de esta plaga cuarentenaria, comúnmente conocida como "polilla de la vid", fue detectada por primera vez en el país en dos predios en la provincia de Mendoza. Lobesia botrana es nativa de la región Paleártica, está ampliamente distribuida en Europa Occidental, Asia Central y Norte de frica (Razowski, 2003). Este lepidóptero ocasiona la disminución de los rendimientos debido a la destrucción de bayas. Además provoca una importante prdida de la calidad de la fruta por las heridas que produce en los granos, lo que favorece la aparición de podredumbres causadas por diversos hongos (Mondy et al., 1998; Armendariz et al., 2007). En uvas para vinificar, los residuos fúngicos generan aromas y sabores desagradables en los vinos, además de los problemas de clarificación durante el proceso de vinificación. Asimismo, causa perjuicios económicos indirectos, a travs de las regulaciones que imponen los países importadores en el comercio internacional (Mendoza, 2011).
Varios lepidópteros fueron criados en laboratorio con el objeto de complementar los parámetros obtenidos con modelos fenológicos. El estudio del desarrollo de Cydia pomonella L. en laboratorio y a campo, permitió a Rock & Shaffer (1983) determinar el umbral de desarrollo para cumplir las distintas etapas de su ciclo. Shaffer & Gold (1985) desarrollaron un modelo de simulación que predice la fenología de una población de C. pomonella basado en la técnica de cohortes iterativas. Sporleder et al. (2004) establecieron un modelo para predecir el tiempo de desarrollo de Phthorimaea operculella Zell., en zonas productoras de papa.
La reciente introducción de L. botrana en la provincia de Mendoza requiere el estudio de su comportamiento y para ello es imprescindible conocer sus parámetros biológicos y poblacionales. Esta información sirve de base para obtener modelos de predicción útiles en programas de monitoreo a campo, contribuyendo de esta manera al manejo integrado de la plaga.
La construcción de tablas de vida y fertilidad permiten comprender la dinámica poblacional, las tasas de supervivencia y reproducción (Carey, 1993; Cividanes, 2002), así como estudiar aspectos de la biología de una especie, tales como tiempo de desarrollo y fecundidad (Kazak et al., 2002). Las tablas de vida representan de manera sintética y numérica las principales características concernientes a la población en estudio (Rabinovich, 1978) y constituyen una herramienta básica para elaborar estrategias de control (Southwood & Henderson, 2000) permitiendo estudiar el impacto de los enemigos naturales sobre una población en programas de control biológico. Contrastando las tablas de vida obtenidas de poblaciones con y sin enemigos naturales, es posible evaluar la repercusión de un controlador a través de la cuantificación de los parámetros demográficos (Bellows Jr. et al., 1992). En ensayos de control químico en laboratorio, es posible construir tablas de vida y observar la infuencia de los insecticidas sobre los parámetros de la tabla de vida
y de fertilidad (Stark et al., 2007). Además sirven de base para representar las curvas de supervivencia de una población. Entre ellas se distinguen cuatro tipos para clasificar a los organismos. Tipo I: cuando la probabilidad de sobrevivir en todas las etapas es la misma, ocurriendo una muerte masiva hacia la edad final. Tipo II: el número de muertes se mantiene constante por unidad de tiempo. Tipo III: el número de individuos que muere a medida que la población envejece es cada vez menor, y tipo IV: donde la mortalidad afecta prioritariamente a los individuos jóvenes, luego la mortalidad se reduce notablemente (Rabinovich, 1982).
La capacidad de reproducción de una especie constituye un aspecto fundamental para el estudio de una población de organismos vivos (Rabinovich, 1982). En ese sentido, las tablas de vida resultan una herramienta apropiada para conocer importantes parámetros biológicos y demográficos (Maia et al., 2000). La tasa intrínseca de incremento poblacional (rm), es un parámetro que evalúa el impacto de la nutrición de una población sobre su potencial reproductivo. En laboratorio sirve para comparar dietas artificiales (Portilla et al., 2013). También puede ser usada para evaluar el nivel de resistencia a plagas (Birch, 1948; Southwood & Henderson, 2000). Las plantas hospedantes que logran bajos valores de rm presentan comparativamente mayor resistencia que aquellas en las que los insectos muestran valores de incremento altos (Razmjou et al., 2006). Este estadístico también ha sido utilizado en el estudio de enemigos naturales para la elaboración de estrategias de control (Orphanides & Gonzáles, 1971).
En virtud de la escasa información referida a los aspectos biológicos y poblacionales de la polilla de la vid en nuestra zona y dada la importancia como plaga cuarentenaria para Argentina se propuso como objetivos del presente trabajo construir una tabla de vida horizontal para L. botrana, determinar los parámetros poblacionales, la proporción sexual, la fecundidad y fertilidad bajo condiciones de laboratorio, información de base para la aplicación de técnicas de manejo de la plaga.
El material entomológico utilizado se obtuvo en la Estación Experimental Agropecuaria Mendoza del Instituto Nacional de Tecnología Agropecuaria (EEA Mendoza INTA), de acuerdo a la técnica de cría desarrollada por Herrera et al. (2016).
Se realizó el seguimiento de una cohorte de 250 huevos. Cada postura fue colocada individualmente en recipientes provistos de dieta artificial (Ferreira et al., 2003). Se registró diariamente el número de sobrevivientes en cada una de las fases de desarrollo del insecto. Con los resultados obtenidos se confeccionó la tabla de vida utilizando el método específico u horizontal (Rabinovich, 1978; Portilla et al., 2013). Los parámetros utilizados en la tabla de vida fueron: edad (x) que designa la edad exacta de L. botrana desde el nacimiento hasta la muerte; número de sobrevivientes a la edad x (Nx); proporción de individuos sobrevivientes a la edad x (lx); proporción de la cohorte que sobrevive desde el nacimiento hasta la edad x (px); fracción de la cohorte que sobrevive entre dos estadios sucesivos (qx); fracción de la cohorte original que muere en el intervalo de la edad x y la siguiente (dx); supervivencia por edades o expectativa de vida (Lx); supervivencia total a la edad x (Tx) y esperanza de vida o cantidad promedio de tiempo que queda por vivir para los individuos que sobreviven a una edad determinada (ex).
Una vez alcanzado el estado adulto se formaron 92 parejas que fueron consideradas réplicas. Cada pareja se colocó individualmente en una botella de PVC transparente de 500 ml utilizada como cámara de reproducción, la que fue observada diariamente a fin de determinar el periodo de pre-oviposición, oviposición y post-oviposición, y registrar el número de huevos colocados por hembra / día para el cálculo de la fecundidad neta (mx) y la determinación de la fecundidad bruta (Mx), siendo esta la capacidad de reproducción de una hembra durante toda su vida. La fertilidad de las posturas se determinó considerando el número de huevos que eclosionaron. Los datos obtenidos se utilizaron para la confección de una tabla de fertilidad, que describe la capacidad de las hembras de edad x de dejar descendencia. Se estimaron los parámetros demográficos y se registró el momento de muerte de los adultos para calcular la longevidad media por sexo. Se calculó la fecundidad neta (mx) que representa el número de hembras obtenidas por cada hembra de edad x. La tasa de reproducción (Ro) que se define como el número promedio de descendientes dejados por cada hembra durante su vida. La tasa intrínseca de crecimiento poblacional (rm) expresa la capacidad del aumento en el número de hembras por hembra por unidad de tiempo. La tasa de incremento finito (λ) indica el número de individuos que se agregan a la población por individuo y por unidad de tiempo. El valor reproductivo por edad (Vx) que se define como la contribución de un individuo de edad x a la población de la siguiente generación. El tiempo generacional (T) es la edad promedio de las hembras en una cohorte en el momento del nacimiento de su descendencia. El parámetro D representa el tiempo que necesita una población para duplicarse.
RESULTADOS
A continuación (Tabla I) se exponen los valores de la tabla de vida de L. botrana en condiciones de laboratorio
Tabla I. Tabla de vida horizontal de una cohorte de huevos (n=250) de Lobesia botrana. x: edad exacta de L. botrana desde el nacimiento hasta la muerte. Nx: número de sobrevivientes a la edad x. lx: proporción de individuos sobrevivientes a la edad x. px: proporción de la cohorte que sobrevive desde el nacimiento hasta la edad x. qx: fracción de la cohorte que sobrevive entre dos estadios sucesivos. dx: fracción de la cohorte original que muere en el intervalo de la edad x y la siguiente. Lx: supervivencia por edades o expectativa de vida. Tx: supervivencia total a la edad x. ex: esperanza de vida o cantidad promedio de tiempo que queda por vivir para los individuos que sobreviven a una edad determinada
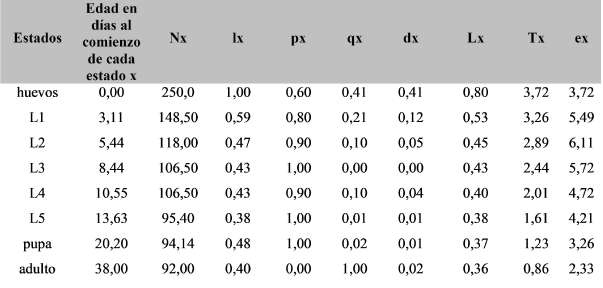
La figura 1 representa la supervivencia y la fecundidad de la cohorte de L. botrana criadas en condiciones de laboratorio. La curva de supervivencia obtenida responde a la de tipo IV (Rabinovich, 1982), donde la mortalidad se observa en el momento de eclosión de huevos y el primer estadio larval. Superada esta etapa, la mortalidad se reduce considerablemente y la curva se estabiliza lográndose una supervivencia constante en los estados sucesivos (Rabinovich, 1978). La curva de fecundidad muestra que las hembras comenzaron a oviponer el día 30. El periodo fecundo de la polilla se prolongó durante 7 días, obteniendo el máximo de posturas al día 34. Este valor luego decrece hasta hacerse 0 el día 37.
Fig. 1. Supervivencia (lx) expresada como proporción en base 250 individuos vivos y fecundidad (mx) expresada como el número de huevos colocados por hembra de Lobesia botranacriada a 25 ºC de temperatura y 25% de humedad relativa.
La fecundidad bruta de la polilla de la vid para las condiciones del ensayo fue de 134,84 ± 15,68 huevos / hembra a lo largo de su vida, y la fertilidad de las posturas del 91,6% (Fig. 2).
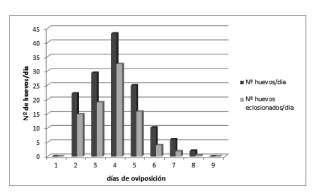
Fig. 2. Fecundidad y fertilidad de Lobesia botrana en condiciones de laboratorio: temperatura 25 ± 2 ºC, luminosidad 16:8 L/O y humedad relativa 30-50%.
La figura 3 muestra la curva del valor reproductivo por edades, se observa un crecimiento sostenido a partir del décimo día de vida hasta llegar a un pico el día 20, antes del inicio de la actividad reproductiva de la plaga, a partir de ese momento declina progresivamente hasta la muerte de los individuos de la cohorte.
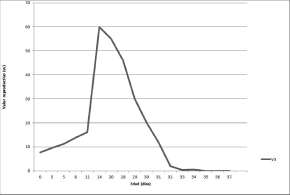
Fig. 3. Valor reproductivo específico por edades de Lobesia botranaen condiciones de laboratorio.
La duración media en días de los períodos pre-reproductivo, reproductivo, post-reproductivo y la longevidad adulta de L. botrana, para las condiciones del ensayo, se exponen en la Tabla II.
Los parámetros demográficos de L. botrana obtenidos bajo las condiciones del experimento (Tabla III) demuestran que cada hembra es reemplazada por 66,82 hembras en un tiempo medio de 30,61 días. Además, la tasa de incremento finito indica que por cada hembra contemporánea, habrá 1,073 hembras al día siguiente. La población se duplicaría en 9,76 días.

DISCUSIÓN
BIBLIOGRAFÍA CITADA
La curva de supervivencia obtenida por Moreau et al. (2006) es similar a la de este estudio, mientras que la curva de fecundidad difere a la observada por Moreau et al. (2015) dado que polillas alimentadas sobre dieta a base de Pinot noir, mantuvieron la oviposición estable durante los primeros cuatro días y luego disminuyó. En las colonias mantenidas sobre otros cultivares, como Chardonnay, el número de huevos colocados disminuyeron linealmente con los días de oviposición.
Moreau et al. (2006) observaron que la fecundidad bruta de la polilla alimentada sobre la variedad Chardonnay fue de 104 huevos / hembra y la fertilidad del 91,34%. Por su parte Nadel (2011) determinó que la fecundidad de hembras criadas sobre la dieta artificial desarrollada por Guennelon et al. (1970, 1975) fue 10 huevos / hembra y la fertilidad del 81%.
La estimación de la edad de mayor valor reproductivo es importante cuando se desea controlar una especie plaga, dado que permite evaluar la estrategia de control (Rabinovich, 1972). En el caso d e L. botrana, sobre la base del conocimiento de su dinámica poblacional a campo, este concepto podría ser aplicable para la primera y segunda generación a campo, ya que a partir de la tercera, de acuerdo a los estudios de Dagatti & Becerra (2015), existe un alto grado de superposición de estados de la plaga, por lo que resulta difícil establecer en ese momento del año la estrategia de control.
Los valores medios de períodos pre-reproductivo, reproductivo y longevidad obtenidos diferen de los observados por Moreau et al. (2006), quienes determinaron que el período de pre-oviposición de la polilla de la vid alimentada sobre uvas de la variedad Chardonnay, fue de 4,81 ± 0,46 días, el de oviposición fue de 5,4 ± 0,33 días y la longevidad de los adultos de 10,21 ± 0,30 días.
CONCLUSIÓN
La tabla de vida de L. botranaproveyó información básica para la comprensión de su ciclo de vida, capacidad de reproducción y supervivencia en condiciones controladas. Las tablas obtenidas a partir de ensayos bajo condiciones controladas pueden tener un valor relativo para evaluar el potencial real de la especie en condiciones de campo, debido a que muchos factores no contemplados en laboratorio infuyen en la supervivencia y en las tasas de crecimiento y fecundidad. Sin embargo, estos estudios biológicos permiten obtener datos necesarios para el análisis del potencial biótico bajo condiciones específicas, y pueden ser utilizados para la simulación de modelos para uso en el campo que incluyan otros factores.
BIBLIOGRAFÍA CITADA
Armendariz, I., Campillo, G., Perez-Zanz, A., Capilla, C., Juárez, J.S., & Miranda, L. (2007) La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Boletín de Sanidad Vegetal - Plagas, 33, 447-489. [ Links ]
Bellows Jr., T.S., Van Driesche, R.G., & Elkinton, J.S. (1992) Life-table construction and analysis in the evaluation of natural enemies. Annual review of entomology, 37(1), 587-612. [ Links ]
Birch, L.C. (1948) The Intrinsic Rate of Natural Increase of an Insect Population. Journal of Animal Ecology, 17, 15-26. [ Links ]
Carey, J.R. (1993) Applied Demography for Biologists. Oxford University Press, New York. [ Links ]
Cividanes, F.J. (2002) Tabelas de vida de fertilidade de Brevicoryne brassicae (L.) (Hemiptera: Aphididae) em condicoes de campo. Neotropical Entomology, 31, 419-427.
Dagatti, C.V., & Becerra, V.C. (2015) Ajuste de modelo fenológico para predecir el comportamiento de Lobesia botrana (Lepidoptera: Tortricidae) en un viñedo de Mendoza, Argentina. Revista de la Sociedad Entomológica Argentina, 74(3-4), 117-122.
Ferreira, A., Bastos, M., & Aguiar, A. (2003) Criação de traca da uva Lobesia botranaDen. y Schiff. En: Actas do Vl Encontro Nacional de Protecçao Integrada, 2003, Castelo Branco-Portugal. pp. 83-88.
Guennelon, G., Sender, C., D'arcier, E., & Audemanrd, H. (1970) Mise au point d'un milieu artificiel pour l' élevageau laboratorio des larves de l' Eudemis de la vigne Lobesia botranaDen. et Schiff. (Lepidoptera Tortricidae). Annales de Zoología, Ecología Animal, 2, 51-77.
Guennelon, G., D'arcier, E., & Trincal, J. (1975) Description d' une production massive de l' Eudemis de la vigne sur milieu artificiel Lobesia botranaDen. et Schiff. (Lepidoptera Tortricidae). Annales de Zoología, Ecología Animal, 7, 295-309.
Herrera, M.E., Dagatti, C.V., & Becerra, V.C. (2016) Método práctico de cría masiva de Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae) en condiciones de laboratorio. Revista de la Sociedad Entomológica Argentina, 75(3-4), 160-164.
Kazak, C., Yildiz, S., & Sekeroglu, E. (2002) Biological characteristics and life tables of Neoseiulus umbraticus Chant (Acari, Phytoseiidae) at three constant temperatures. Anzeiger für Schädlingskunde, 75(5), 118-121.
Maia, A.H.N., Luiz, A.J.B., & Campanhola, C. (2000) Statistical inference on associated fertility life table parameters using jackknife technique: computational aspects. Journal of Economic Entomology, 93, 511-518.
Mendoza, G. (2011) Daños producidos por Lobesia botrana en los viñedos mendocinos. En: SAVE 2011, Simposio Argentino de Viticultura y Enología,2011, Mendoza. P. 125.
Mondy, N., Charrier, B., Fermaud, M., Pracros, P., & Corio-Costet, M.F. (1998) Mutualism between a phytopathogenic fungus (Botrytis cinerea) and a vineyard pest (Lobesia botrana). Positive effects on insect development and oviposition behaviour. Comptes Rendus de l' Académie des Sciences-Series III-Sciences de la Vie, 321(8), 665-671.
Moreau, J., Benrey, B., & Thiéry, D. (2006) Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana(Lepidoptera: Tortricidae). Bulletin of Entomological Research, 96, 205-212.
Moreau, J., Monceau, K., & Thiéry, D. (2015) Larval food infuences temporal oviposition and egg quality traits in females of Lobesia botrana. Journal of Pest Science, 89, 439-448.
Nadel H. (2011) A Rearing System for European Grapevine Moth at the Otis Insect Containment Facility. CPHST Otis Laboratory Report, pp. 20-26. USDA, Buzzards Bay, MA.
Orphanides, G.M., & Gonzales, D. (1971) Fertility and life tables studies with Trichogramma pretiosum and T. retorridum (Hymenoptera: Trichogrammatidae). Annals of the Entomological Society of America, 64(4), 824-834.
Portilla, M., Ramos-Morales, J., Rojas, G., & Blanco, C. (2013) Life tables as tools of evaluation and quality control for arthropod mass production. Mass production of beneficial organisms (ed. Morales-Ramos, J.A., Rojas, M.G., & Shapiro-Ilan, D.I.), pp. 241-275. Academic Press, Londres.
Rabinovich, J.E. (1972) Valor reproductivo: un parámetro poblacional útil en el control de insectos perjudiciales. Acta Biológica Venezolana, 8, 25-34.
Rabinovich, J.E. (1978) Ecología de las poblaciones animales. Organización de los Estados Americanos, Washington, D.C.
Rabinovich, J.E. (1982) Introducciónala ecología de poblaciones animales. CECSA. México.
Razmjou, J., Moharramipour, S., Fathipour, Y., & Mirhosein, S.Z. (2006) Effect of cotton cultivar on performance of Aphis gossypii (Homoptera: Aphididae) in Iran. Journal of Economic Entomology, 99, 1820-1825.
Razowski, J. (2003) Tortricidae of Europe. Volume 2. Olethreutinae, Franisek Slamka, Bratislava.
Rock, G.C., & Shaffer, P.L. (1983) Developmental rates of codling moth (Lepidoptera: Olethreutidae) reared on apple at four constant temperatures. Environmental Entomology, 12(3), 831-834.
SENASA (2010) Resolución 122/2010. http://www.senasa.gov.ar/normativas/ resolucion-122-2010-senasa-servicio-nacional-de-sanidad-y-calidad-agroalimentaria [Acceso: diciembre de 2016]
Shaffer, P.L., & Gold, H.J. (1985) A simulation model of population dynamics of the codling moth, Cydia pomonella. Ecological modelling, 30(3-4), 247-274.
Southwood, T.R.E., & Henderson, P.A. (2000) Ecological Methods.3rd Edition. Willey- Blackwell Publishing, Oxford.
Sporleder, M., Kroschel, J., Quispe, M.R.G., & Lagnaoui, A. (2004) A temperature-based simulation model for the potato tuberworm, Phthorimaea operculella Zeller (Lepidoptera; Gelechiidae). Environmental Entomology, 33(3), 477-486.
Stark, J.D., Sugayama, R.L., & Kovaleski, A. (2007) Why demographic and modeling approaches should be adopted for estimating the effects of pesticides on biocontrol agents. BioControl, 52(3), 365-374.

