










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista argentina de antropología biológica
On-line version ISSN 1514-7991
Rev Arg Antrop Biol vol.14 no.1 La Plata Dec. 2012
TRABAJOS ORIGINALES
Morfología craneofacial y estructura genética en poblaciones del centro de Argentina
Mariana Fabra* y Darío A. Demarchi
IDACOR-CONICET/Laboratorio de Bioantropología. Museo de Antropología. Facultad de Filosofía y Humanidades. Universidad Nacional de Córdoba. Córdoba. Argentina
Financiamiento: FONCyT (PICT 2007-1549). Ministerio de Ciencia y Tecnología Córdoba (PID 2008).
*Correspondencia a: Mariana Fabra. Museo de Antropología. Facultad de Filosofía y Humanidades. Universidad Nacional de Córdoba. Hipólito Yrigoyen 174. 5000 Córdoba.
Argentina. E-mail: mfabra@gmail.com
Recibido: 25 Julio 2011
Aceptado: 02 Febrero 2012
Resumen
Se analizan las relaciones interpoblacionales y los roles relativos cumplidos por las fuerzas evolutivas en la diferenciación biológica de un conjunto de poblaciones, que habitaron diversas regiones geográfico-ecológicas del cono sur sudamericano, con especial atención a aquellas que ocuparon la región central de Argentina. La muestra está compuesta por 18 poblaciones, incluyendo 398 adultos masculinos, analizados en 10 variables craneofaciales. Se realizó el análisis de coordenadas principales a partir de distancias D2, así como el análisis de Relethford y Blangero, basado en el modelo de Harpending y Ward para datos morfológicos. El análisis de distancias muestra a Córdoba en una posición intermedia y relativamente diferenciada de las otras poblaciones, hecho consistente con su ubicación geográfica y que resalta ese factor como condicionante de la evolución de la población. Por otra parte, muestra similitudes con Santiago del Estero y marcadas diferencias con San Luis, la otra muestra poblacional de las Sierras Centrales. El análisis de Relethford-Blangero permite suponer para Córdoba un tamaño efectivo poblacional relativamente grande y la existencia de alto flujo génico, con las poblaciones fuera de la región, mientras que la posición de San Luis sugiere la acción de fuerzas aleatorias actuando sobre una población pequeña.
Palabras clave: Variables craneométricas; Análisis de Relethford-Blangero; Análisis discriminante; Análisis de coordenadas principales; Sierras Centrales
Abstract
The aim of this paper is to analyze population relationships and the relative roles fulfilled by the evolutionary forces in the biological differentiation of a group of populations who lived in different geographic-ecological regions of the southern cone of South America, with particular emphasis on those that occupied the central region of Argentina. The sample consists of 18 populations, including 398 adult males, analyzed by 10 craniofacial variables. Principal coordinates analysis based on D2 distance and Relethford and Blangero analysis were carried out, based on the model of Harpending and Ward for morphological data. The distance analysis showed Córdoba in an intermediate position and relatively differentiated from the other populations, a fact consistent with its geographical location, that suggests that geography was a conditioning factor in the evolution of the population. On the other hand, Córdoba showed similarities with Santiago del Estero, but with marked differences with San Luis, the other population sample from the Sierras Centrales. The Relethford-Blangero analysis suggests for Córdoba, a relatively large effective population size and the existence of high gene flow with populations outside the region, while the position of San Luis suggests the action of random forces acting on a small isolated population.
Key words: Craniometrics; Relethford-Blangero analysis; Discriminant analysis; Principal coordinates analysis; Sierras Centrales
La variación biológica de las poblaciones originarias de la Argentina ha sido extensamente estudiada a partir de la morfología craneana, utilizando medidas lineales (Cocilovo y Di Rienzo, 1984-1985; Cocilovo y Neves, 1988- 1989; Rothhammer et al., 1988-1989; Méndez y Salceda, 1995; Lalueza Fox et al., 1996; González-José et al., 2001a, b; González-José, 2003; Sardi et al., 2005), rasgos epigenéticos (Del Papa, 2008) y más recientemente, mediante análisis de morfometría geométrica (Bernal et al., 2006; Perez et al., 2007; González et al., 2010). Recientemente, han surgido aproximaciones que evalúan la diferenciación biológica entre poblaciones y los procesos evolutivos implicados, a partir de la variación morfométrica dental (Bernal et al., 2010). Asimismo, el desarrollo en años recientes de técnicas moleculares, ha permitido incrementar los estudios sobre poblaciones contemporáneas (Dipierri et al., 1998; Demarchi et al., 2001a, 2005; Dejean et al., 2004; García y Demarchi, 2006, 2009) y arqueológicas (Demarchi et al., 2001b; Carnese et al., 2010; Nores y Demarchi, 2011). Sin embargo, numerosas preguntas acerca de los orígenes, las estrategias de dispersión y la posterior evolución local de las poblaciones continúan sin respuesta.
Con respecto al estado actual del conocimiento sobre las poblaciones prehispánicas del área central del territorio argentino -particularmente aquellas que ocuparon la región austral de las Sierras Pampeanas- en las últimas décadas se han llevado adelante investigaciones que han permitido sugerir diversas y a veces contrapuestas hipótesis para explicar el poblamiento de la región. Inicialmente, basados en la evidencia arqueológica y en menor medida antropológica, los primeros trabajos asociaron a estas poblaciones con corrientes migratorias procedentes del Noroeste argentino (Cocilovo, 1984). Otros autores, señalaron semejanzas morfológicas con poblaciones de la región cuyana, en el centro-oeste del país (Canals Frau, 1944). Más recientemente, otros investigadores han verificado, a través de análisis morfométricos lineales, similitudes entre los habitantes de la región central del país y los pueblos que ocuparon Cuyo y el sur del Litoral (Cocilovo, 1984; Marcellino y Colantonio, 1993). Nuestros propios resultados obtenidos a partir de marcadores del ADN mitocondrial en poblaciones contemporáneas (García y Demarchi, 2006, 2009), rasgos epigenéticos, variables métricas lineales y análisis morfogeométricos en muestras esqueletales (Fabra et al., 2005, 2007; Fabra, 2008a, b, 2009; Fabra y Demarchi, 2009) y evidencia arqueológica (Laguens et al., 2007; Laguens, 2009) permitieron abonar hipótesis acerca de un origen común para las poblaciones que ocuparon la región central del país con los grupos humanos que habitaron Patagonia. Estos resultados se han visto complementados por aquellos obtenidos recientemente a partir de ADN mitocondrial de muestras arqueológicas de la provincia de Córdoba, los cuales sugieren diferencias estadísticamente significativas entre las frecuencias de los haplogrupos mitocondriales nativos de las poblaciones actuales y muestras arqueológicas (Nores y Demarchi, 2011). Se pudo estimar un incremento gradual en la frecuencia de haplogrupos C y D (de mayor frecuencia en poblaciones del sur de Argentina), en detrimento de haplogrupos A y B, en momentos previos a la conquista española. Además, los autores han señalado una tendencia hacia una diferenciación regional entre poblaciones que habitaron las sierras y la llanura, con una mayor preponderancia de haplogrupo B en estas últimas.
Sin embargo, a pesar de que estos estudios han generado un cúmulo de evidencia que permite incrementar el conocimiento acerca de la historia biológica de las poblaciones del centro del país, existen temas aún por resolver. Uno es el poblamiento inicial y el proceso posterior de colonización y expansión en este espacio; otro se refiere a la evolución biológica de esas poblaciones, es decir, la importancia relativa que tuvieron sobre ellas, las fuerzas evolutivas direccionales (migración, selección) y las aleatorias (efecto fundador, deriva génica), así como los patrones espaciales que configuran dicha variación. En general, los diferentes métodos estadísticos de ordenación (análisis de componentes principales, de coordenadas principales y de escalamiento multidimensional) permiten identificar las afinidades genéticas entre poblaciones, pero no identificar qué fuerzas evolutivas configuraron la variación morfológica observada. Este tipo de aproximación, puntualmente aquellos que aplican el análisis de Relethford-Blangero (1990) -basado a su vez en el modelo de Harpending y Ward (1982), que se orientan a estimar la importancia que han tenido las diferentes fuerzas evolutivas en la diferenciación biológica de las poblaciones, no suelen ser usualmente presentadas en los estudios realizados sobre poblaciones del cono sur americano, con excepción de los trabajos de Varela y Cocilovo (2002), González-José (2003) y Varela et al. (2008), entre otros.
En este trabajo hemos utilizado el modelo de Relethford y Blangero (1990) para investigar la importancia relativa que han tenido las fuerzas evolutivas sistemáticas y aleatorias en la diferenciación biológica de las poblaciones originarias, que ocuparon el centro del territorio argentino y otras regiones geográficas del cono sur sudamericano.
Material y métodos
El área de estudio
Nuestra área de estudio se ubica en la región austral de las Sierras Pampeanas, entre los 30° y 35° de latitud Sur y 62° y 66° de longitud Oeste, abarcando los cordones montañosos de las provincias de Córdoba y San Luis y las llanuras orientales y occidentales aledañas (Fig. 1). Desde el punto de vista fisiogeográfico, esta región presenta contrastes entre sierras y llanuras: la planicie oriental, correspondiente a la llanura pampeana y las sierras en el Oeste. Entre las dos principales unidades orográficas -las Sierras de Córdoba y las de San Luis- se desarrolla una segunda llanura o planicie occidental, separando ambos cordones mediante un amplio valle intermontano. Desde un punto de vista fitogeográfico, la provincia de Córdoba representaría, dentro del dominio Chaqueño, a las provincias Chaqueña del Espinal y Pampeana, que la atravesarían como bandas longitudinales en sentido Noreste- Sudoeste (Cabrera, 1976). En el sector norte de la región, existen dos grandes humedales: la Laguna de Mar Chiquita, en el Noreste y las Salinas Grandes, en el Noroeste. Hacia el Sudoeste de la región, un sistema de numerosas lagunas saladas y dulces, de poca profundidad, forman en el sur de la Provincia de San Luis otro importante humedal con más de 180 cuerpos de agua.
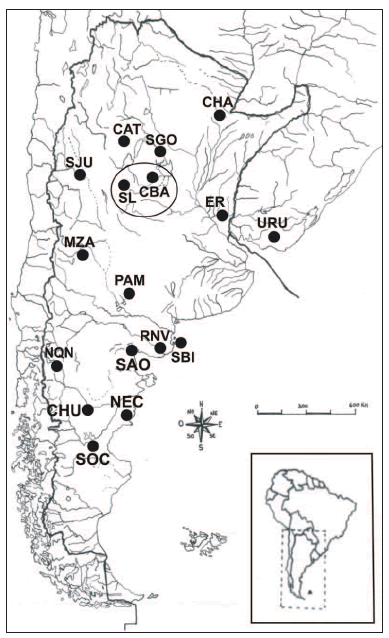
Fig. 1. Mapa del cono sur sudamericano, mostrando la localización aproximada de las 18 poblaciones incluidas en este estudio.
Cambios y continuidades: el registro arqueológico regional
Las investigaciones arqueológicas llevadas adelante a partir de finales del siglo XIX y particularmente en las últimas décadas, sugieren que hubo un poblamiento inicial hace aproximadamente 10000 años A.P., por grupos cazadoresrecolectores, portadores de una tecnología lítica conocida como puntas "cola de pescado" (Laguens et al., 2007), tecnología que ha sido datada en otros sitios de Uruguay y Argentina en la transición Pleistoceno-Holoceno (Politis et al., 2004). Aproximadamente hace 6500 años A.P. se vislumbran ciertos cambios en el registro arqueológico, particularmente con respecto al estilo tecnológico de ciertos objetos (reducción en el tamaño y cambios en la forma de puntas de proyectil, sugiriendo modificaciones en las estrategias de caza hacia presas de menor tamaño y en el uso de nuevos instrumentos, tales como propulsores o atlatls), así como en el uso del paisaje (mayor ocupación del espacio, incremento en el número de sitios arqueológicos) acompañado por condiciones climáticas más templadas. Un cambio hacia condiciones secas y semi-áridas se habría producido hace aproximadamente 4200 años A.P., con una duración aproximada de 2000 años, cuyas consecuencias más notables fueron la erosión y deflación de los suelos, el déficit hídrico, la disminución del nivel de lagos y ríos y el desarrollo de las Salinas Grandes, implicando cambios no sólo en la distribución de los recursos potencialmente explotables, sino en la oferta ambiental y estrategias de subsistencia (v.g. almacenamiento de granos y semillas) (Laguens, 1999; Laguens et al., 2007).
Aproximadamente hace 1500 años A.P. aparece en el registro arqueológico regional una serie de evidencias que sugieren el desarrollo de nuevas estrategias de subsistencia, así como en las producciones tecnológicas, tales como la producción de alimentos y la manufactura de la cerámica. Estas fueron prácticas que se incorporaron gradualmente, posiblemente desde poblaciones de la región chaqueña, a las tradicio nes basadas en la caza y recolección, así como el establecimiento en poblados permanentes, habría derivado en cambios a largo plazo en el estilo de vida de estas poblaciones, modificando la vida y la forma de relacionarse entre las personas. Asimismo, se observa una tendencia hacia una mayor diferenciación a nivel intrarregional, probablemente debido a las diversas adaptaciones realizadas por estas poblaciones a los ambientes (v.g. sierras y llanuras) y a formas de marcar diferencias con otros grupos por ejemplo, a través de diversos estilos tecnológicos o comportamientos mortuorios (Fabra et al., 2009).
La muestra
El conjunto de individuos relevado para la región central de Argentina procede de dos provincias, Córdoba (CBA) y San Luis (SL). CBA incluye a 70 cráneos de individuos adultos masculinos, procedentes de 25 sitios arqueológicos preservados en museos públicos y privados. La muestra de SL incluye solamente 8 individuos adultos masculinos, de diferentes sitios. En el caso de CBA, se poseen 24 fechados radiocarbónicos obtenidos mediante AMS (Accelerator Mass Spectrometry) directamente de colágeno, los cuales permiten ubicar a estos individuos en un rango cronológico de 4525±20 y 387±41 14C años A.P. Para el resto de las muestras, se supone en función de los contextos arqueológicos, que pueden corresponder al Holoceno tardío (~2500- 400 años A.P.). No se dispone de cronologías absolutas para las muestras de SL incluidas en este estudio; de todas formas, los contextos arqueológicos sugieren que pueden corresponder al Holoceno tardío.
Con el fin de llevar adelante los análisis interpoblacionales y estimar semejanzas biológicas, se incluyeron en el análisis 320 individuos adultos masculinos, agrupados en 17 poblaciones prehispánicas que habitaron diversas regiones geográficas del cono sur de Sudamérica (ver Tabla 1, Fig. 1). Respecto a la asignación cronológica de las muestras, se consideró a) la información radiocarbónica disponible y b) la información sobre deformaciones artificiales del cráneo, ya
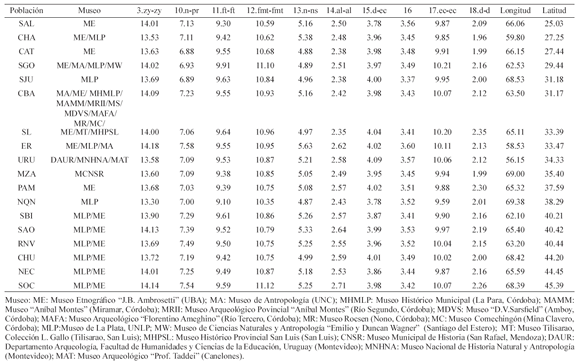
que se han observado correspondencias en otras regiones del país -región pampeana, patagónica, sudeste de la región pampeana- entre ciertas clases de deformaciones craneales intencionales y la temporalidad de las muestras1. Tomando en cuenta estas correspondencias y en los casos de no disponer de información cronológica absoluta, se consideró a esta práctica como indicadora de la cronología relativa de las muestras. Las del noreste de la Patagonia son del Valle de Río Negro (RNV, n=14), cercanas a la localidad de San Antonio Oeste (SAO, n=8) y a la península de San Blas e Isla Gama (SBI, n=34). Estas muestras corresponden a grupos de cazadores-recolectores terrestres, definidos en estudios etnográficos como Tehuelches. En función de la información cronológica relativa que ofrecen las deformaciones craneales que presentan algunas muestras, así como los contextos de entierro, las muestras de RNV y SAO proceden de un rango cronológico que va desde el Holoceno medio (~3500-2500 años A.P.) al tardío (~2500-400 años A.P.), mientras que la muestra de SBI ha sido asignada al Holoceno tardío final. Las muestras del centro de Patagonia proceden de diversas regiones de la provincia de Chubut y fueron denominadas en función de la región geográfica de procedencia: Valle del Río Chubut y costa central (NEC, n=16), Lago Colhue Huapi y Río Mayo (SOC, n=7) y provincia de Chubut (CHU, n=12). Estas muestras han sido consideradas, en función de la deformación craneana artificial, como del Holoceno tardío (~1500-400 años A.P.). Las muestras del noroe ste de Patagonia (NQN, n=6) fueron obtenidas en expediciones de fines del siglo XIX y proceden de diversos sitios. Al no poseer cronologías absolutas, se consideró a la deformación artificial como indicador relativo de la antigüedad de las muestras y se las adscribió a momentos finales del Holoceno tardío (~1500-400 años A.P.). Por otra parte, los individuos relevados del oeste de las región pampeana (PAM, n=6) proceden de un único sitio arqueológico datado en 1030-370 14C años A.P. (sitio Chenque I, Parque Nacional Lihue Calel, provincia de La Pampa) y corresponden a cazadores-recolectores continentales. Las muestras de la región chaco-santiagueña (SGO, n=36, provincia de Santiago del Estero), noroeste argentino (CAT, n=9, provincia de Catamarca; SAL, n=11, provincia de Salta) y norte de la región cuyana (SJU, n=16, provincia de San Juan), proceden de poblaciones que adoptaron la agricultura y desarrollaron asentamientos permanentes y sedentarios en el Holoceno tardío. La muestra del sur de Cuyo (MZA, n=23, provincia de Mendoza) fue datada mediante métodos absolutos y se la ubicó en el Holoceno tardío y corresponde a grupos con economía mixta, basados principalmente en la caza, la recolección y en menor medida, el cultivo del maíz y otros vegetales. Finalmente, las muestras del noreste argentino (CHA, n=18, provincia del Chaco) y noreste de la región pampeana (ER, n=16, delta del río Paraná, provincia de Entre Ríos; URU, n=10, República Oriental del Uruguay) representan poblaciones con economía mixta, basada en la agricultura, caza, pesca y recolección, que también fueron adscriptas a momentos tardíos del Holoceno. En la Tabla 1 se presentan los promedios de las 10 variables craneométricas para cada una de las 18 poblaciones estudiadas, así como la información respecto a su ubicación geográfica (latitud y longitud).
Tabla 1. Promedios de las 10 variables craneométricas de las poblaciones estudiadas. Individuos masculinos
Métodos analíticos
Para controlar la variación en función de diferencias sexuales, solamente se trabajó con individuos masculinos. En cuanto al análisis bioantropológico de los restos, se consideraron los procedimientos establecidos por Buikstra y Ubelaker (1994) para la determinación del sexo y estimación de edad. Primeramente, los restos fueron acondicionados e inventariados. Para la determinación del sexo, se tuvieron en cuenta principalmente las características de la pelvis (región subpúbica, escotadura ciática, surco preauricular) y en el caso de no estar presente o estar muy fragmentada, se analizó la morfología craneana (desarrollo de la cresta nucal, tamaño del proceso mastoideo, ángulo del margen supraorbital, prominencia de la glabela, proyección de la eminencia mental). Para la estimación de la edad, se examinó cada caso y se lo incorporó a las categorías de infante, niño, adolescente o adulto. Posteriormente, se seleccionó el criterio apropiado para la definición de la edad dentro de alguna de estas categorías. Se tuvieron en cuenta aquellos cambios que se producen en la sínfisis púbica, en la superficie auricular del ilión y en el grado de fusión de las epífisis (Buikstra y Ubelaker, 1994).
Para poner a prueba la influencia de la deformación craneana artificial sobre el conjunto de medidas craneanas presentado por Buikstra y Ubelaker (1994), se realizó un análisis de la varianza (ANOVA). Para minimizar los efectos de la deformación artificial en las medidas craneanas, se eliminaron aquellas variables afectadas significativamente por este factor. Esta opción se prefirió a aquella que supone eliminar de los análisis a los individuos que presentan deformación artificial, ya que de esta forma se reduce significativamente la muestra. Las pruebas de ANOVA (datos no mostrados) pusieron de manifiesto la influencia significativa sobre las variables que miden el ancho del neurocráneo. Sobre el resto de las variables, principalmente faciales, los resultados fueron no significativos. Estos resultados confirman los obtenidos por Cocilovo (1975) a partir de variables craneofaciales y más recientemente por Perez (2006) a partir de estudios de morfometría geométrica, sobre la nula influencia de la deformación craneana artificial sobre las variables faciales. Así, para el estudio de la variabilidad morfológica craneofacial, en este trabajo se registraron 10 variables métricas lineales: ancho bicigomático (3.zy-zy), altura facial superior (10.n-pr), ancho frontal mínimo (11.ft-ft), ancho facial superior (12.fmt-fmt), altura nasal (13.n-ns), ancho nasal (14.al-al), ancho orbital (15.d-ec), altura orbital (16), ancho biorbital (17. ec-ec) y ancho interorbital (18.d-d). Los números y letras entre paréntesis indican la codificación otorgada por Buikstra y Ubelaker. El registro craneofacial fue realizado por uno de los autores (MF) para minimizar el error interobservador.
Análisis estadísticos
Modelo de Harpending y Ward (1982)
El análisis de Relethford-Blangero (1990), basado en el modelo de Harpending y Ward (1982), permite analizar los roles relativos cumplidos por las fuerzas evolutivas direccionales y aleatorias, en la diferenciación biológica entre poblaciones. Según este modelo, bajo una presión sistémica uniforme, existe una correlación linear y negativa entre la heterocigosis esperada (He) -en el caso de rasgos morfológicos se considera de manera análoga la variación fenotípica- y la distancia de cada población al centroide o media regional (rii), que correspondería al ancestro común hipotético antes de la subdivisión de la población. Según el modelo, las poblaciones que hayan experimentado aislamiento (dando lugar a la deriva génica) tendrán una variabilidad genética (o fenotípica) inferior a la prevista por el modelo, ubicándose por debajo de la línea de regresión. Por el contrario, las poblaciones que hayan recibido flujo génico externo mayor al promedio regional, presentarán una diversidad genética superior a la predicha y se ubicarán por encima de la línea de regresión.
Si bien el modelo fue desarrollado para ser aplicado a frecuencias alélicas, Relethford y Blangero extendieron su uso al análisis de caracteres cuantitativos, para lo cual John Relethford creó el programa RMET (actualmente en su versión 5.0). El programa cuenta con varias opciones, de acuerdo con las características de las poblaciones y de las variables estudiadas. En este trabajo se utilizó la corrección para el error de muestreo debido al tamaño muestral pequeño (Relethford et al., 1997) y se estimó una heredabilidad promedio de 0.55, tal como sugieren Relethford y Blangero (1990) para rasgos craneométricos. También se obtuvieron a partir del programa RMET 5.0 las distancias D2 entre pares de poblaciones y su proyección en las dos primeras coordenadas principales.
Resultados
Análisis de distancias
La Figura 2 representa las afinidades relativas entre muestras poblacionales basadas en la proyección de las dos primeras coordenadas principales, construidas a partir de la matriz de distancias D2. La distribución de las muestras en la figura permite observar cierta estructura espacial: las poblaciones del noroeste argentino y norte de Cuyo (SJU, SGO, CAT) se agrupan en el cuadrante superior derecho, mientras que las poblaciones del centro (NEC, SOC) y noreste de Patagonia (SAO, RNV, SBI) se agrupan en el cuadrante inferior izquierdo. La muestra de Uruguay aparece cerca de La Pampa (PAM) en el cuadrante inferior derecho. Llamativamente ER y SAL se ubican en el extremo izquierdo de la figura, muy lejos de las poblaciones de la región pampeana y del noroeste, respectivamente. CBA ocupa una posición central en el arreglo, a relativamente bajos valores de distancia de SGO y MZA, ambos hechos en coincidencia con su localización geográfica. Por otra parte, la otra muestra de las Sierras Centrales (SL) se ubica en el extremo derecho de la figura, a grandes valores de distancia del resto de las muestras.
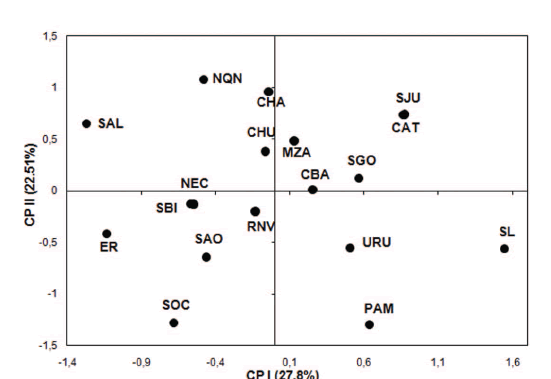
Fig. 2. Mapa genético construido a partir del análisis de coordenadas principales de las distancias D2.
Modelo de Harpending y Ward
En la Figura 3 se presenta el resultado del análisis de la estructura poblacional a partir del modelo de Harpending y Ward (1982). Lo primero que surge es la gran dispersión de las muestras en ambos ejes. Sorprende el amplio rango de variación fenotípica entre muestras y también la gran variación respecto a la distancia al origen (rii) hecho que refleja, de acuerdo al modelo, la diferenciación a partir del ancestro común. CBA y CHU y en menor medida, SGO y NEC, se ubican cercanas al origen y por encima de la recta de regresión, lo cual sugiere poca diferenciación de la población ancestral hipotética, debido a un tamaño efectivo relativamente grande. La variabilidad fenotípica, mayor a la esperada, sugiere aporte genético superior al promedio regional desde fuera de la región o bien, desde poblaciones no incluidas en el presente estudio. SAO, RNV, MZA y en menor grado SBI y URU, se ubican también relativamente próximas a la población ancestral, pero presentan una variación fenotípica por debajo a la esperada, lo cual sugiere presión selectiva uniforme y/o flujo génico restricto entre esas poblaciones. PAM, CAT, NQN y CHA muestran valores intermedios de rii, pero difieren ampliamente en su variación fenotípica. Mientras CAT presenta la mayor variación de todas las muestras estudiadas, PAM es la segunda menos variable, estando NQN y CHA también por debajo de la línea de regresión. La gran diversidad morfológica observada en CAT sugiere, de acuerdo al modelo, elevado flujo migratorio externo. Sin embargo, dado el pequeño tamaño muestral (n=9) y la falta de información respecto al origen de la muestra -procedencia de los individuos y cronología-, nos lleva a sospechar que un valor tan alto de variación podría deberse a que la misma está conformada por individuos provenientes de diferentes poblaciones (temporal y/o espacialmente). Por último, aparece en la figura un grupo de poblaciones que se han diferenciado marcadamente de la población ancestral, pero a través de diferentes mecanismos. SJU, SOC y ER, por encima de la recta de regresión, habrían sufrido aporte migratorio externo mayor al promedio, mientras SAL se ubica exactamente sobre la recta, es decir se ajusta al modelo. Por su parte SL se ubica muy por debajo de la misma, siendo la muestra con menor variación fenotípica, hecho que sugiere que se ha diferenciado a partir de deriva génica.
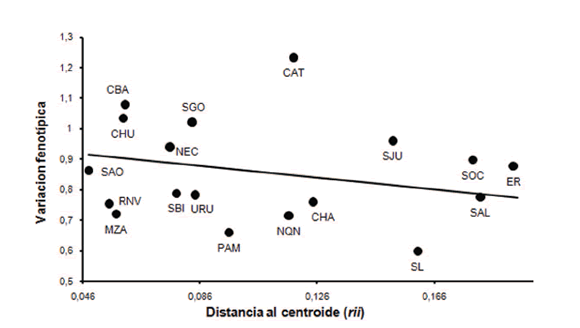
Fig. 3. Análisis de la estructura genética de la población a partir del modelo de Harpending y Ward (1982).
Como subproducto del análisis de Relethford y Blangero, a partir del promedio de
las distancias de cada subpoblación al centroide (rii) puede obtenerse el coeficiente de diferenciación intermuestral. El valor obtenido en este análisis fue muy alto (Fst=0,146 considerando una h2=0,55 y Fst=0,089 con una h2=1), del mismo orden que el encontrado entre poblaciones continentales, al cual contribuyeron principalmente algunas de las poblaciones muy diferenciadas (ER, SOC, SL, SAL) a partir de eventos de deriva y/o aporte migratorio externo.
Discusión
El análisis de distancias muestra a CBA cerca del centro de la figura y relativamente aisla da, aunque en cierta proximidad de SGO y MZA y distanciada de los grupos del noroeste argentino, Cuyo, Pampa y Patagonia. Este resultado se ajusta a su localización geográfica y sugiere que esta variable ha sido un factor muy importante en la evolución de la población a lo largo del tiempo, a través de un relativo aislamiento y a la vez, sin barreras insalvables. Estas características habrían permitido la diferenciación in situ de la población local pero, al mismo tiempo, recibiendo un aporte migratorio importante desde otras regiones.
Si bien la explicación dada en el párrafo anterior resulta del todo plausible, estos resultados muestran ciertas discrepancias con estudios previos basados tanto en morfometría clásica (Fabra, 2009), geométrica (Fabra y Demarchi, 2009), como también a partir de rasgos epigenéticos (Fabra et al., 2005) y moleculares (García y Demarchi, 2006, 2009), donde las distancias genéticas mostraban semejanzas entre CBA y poblaciones de Patagonia. Los factores que influyen sobre los patrones geográficos de diferenciación biológica, son usualmente de compleja interpretación. La falta de consistencia en el patrón evidenciado a nivel interpoblacional puede deberse, según explica Ossemberg (1977) no al tipo de variables per se, sino al conjunto utilizado, ya que las agrupaciones pueden variar en función de las variables incluidas en cada estudio. Esta autora señala que también puede influir el tipo de distancia biológica utilizada, la composición y el n muestral. Al respecto, la incorporación por primera vez de una muestra de tamaño considerable de la población SGO, que presenta la menor distancia tanto con CBA como con SL y por otro lado, su semejanza con poblaciones del noroeste argentino, puede ser una de las causas que explique la configuración final del mapa génico. Previamente, otros investigadores ya habían sugerido vínculos entre poblaciones de las Sierras Centrales y de la región chaco-santiagueña (Cocilovo, 1984; Cocilovo y Di Rienzo, 1984-1985), así como entre estas poblaciones y otras del noreste y centro de Patagonia y noreste de la región pampeana (Marcellino, 1995-1996).
Otro resultado de este estudio que merece ser destacado, es la evidente diferenciación en la morfología craneofacial de las dos muestras del centro del país (CBA y SL), diferencia que había sido notada previamente a partir de estudios que abordaron la variación craneofacial sobre muestras esqueletales (Fabra, 2008a, 2009; Fabra y Demarchi, 2009) y la composición genética, a partir de estudios de ADN mitocondrial sobre poblaciones actuales (García y Demarchi, 2009; Pauro et al., 2010). En la presente investigación, CBA y SL ocupan posiciones bien alejadas entre sí, diferencias que podrían estar sugiriendo histo rias evolutivas distintas para ambas poblaciones.
Con respecto a la estructura poblacional, abordada en este trabajo a partir del modelo de Harpending y Ward (1982), resulta interesante destacar que un conjunto de muestras de la región central de argentina (CBA, SGO), noreste de Patagonia (CHU, NEC, SAO, RNV) y sur de Cuyo (MZA) muestran poca diferenciación con respecto a la población hipotética ancestral, característica que reflejaría un tamaño efectivo grande de la población o bien ausencia de aislamiento, lo cual impidió la diferenciación de las mismas. Algunas poblaciones evidenciaron bajos niveles de diversidad fenotípica y marcada diferenciación de la población ancestral. Si bien en general esta posición suele interpretarse como evidencia de deriva génica, en este caso podría deberse, por lo menos para las muestras de NQN y SL, al reducido tamaño muestral (n=6 y 8 individuos, respectivamente). A diferencia de estas dos poblaciones, CHA cuenta con un tamaño muestral relativamente alto (n=18), por lo cual podría aceptarse sin mayores inconvenientes, que la baja diversidad fenotípica puede deberse a la acción de fuerzas aleatorias que habrían favorecido la pérdida de la diversidad en esta población. Resultados similares fueron reportados por Demarchi y colaboradores (2001b) para ADN mitocondrial y Demarchi (2009) para microsatélites autosómicos en poblaciones Wichi de la provincia del Chaco.
El alto valor de variación fenotípica encontrado para la muestra CAT resulta sospechoso y podría deberse a un error de muestreo más que a una característica poblacional. Esta observación podría hacerse extensiva a la mayoría de las muestras, aunque en menor medida y pone una nota de precaución sobre el alcance de las conclusiones a las que puede arribarse a partir de muestras poblacionales constituidas de manera más o menos incierta, integrada tal vez por individuos que vivieron en diferentes tiempos y que pertenecían a diferentes poblaciones démicas.
Ya se mencionó que el valor de diferenciación genética entre las poblaciones (Fst=0,146) es sumamente elevado. Valores similares han sido reportados anteriormente en poblaciones sudamericanas contemporáneas, como en el caso de 11 poblaciones del Gran Chaco (Demarchi et al., 1998) que presentaron un índice de diferenciación sumamente elevado, a partir de medidas antropométricas (Fst=0,142). También se observó una diferenciación morfológica muy significativa entre muestras de procedencia arqueológica, en el trabajo de González-José et al. (2001b). En este estudio, los autores remarcan que el aislamiento por distancia explicaría las diferencias biológicas observadas entre las poblaciones, principalmente sudamericanas y por ende, los valores de diferenciación más elevados que los obtenidos para poblaciones asiáticas. El Fst obtenido para el conjunto de poblaciones sudamericanas en ese estudio, fue de 0,162; sin embargo, cuando se eliminan del análisis las muestras paleoamericanas, el valor de diferenciación genética disminuye a 0,116 (González-José et al., 2001b:160). Vale destacar que estos valores fueron obtenidos a partir de una estimación de h2=1, por lo cual constituyen valores mínimos.
Recientemente, en un estudio sobre poblaciones nativas sudamericanas a partir de microsatélites autosómicos, Demarchi (2009) ha mencionado la importancia de complementar los estudios acerca de la historia biológica de las poblaciones, con análisis que permitan evaluar cómo determinadas fuerzas evolutivas han moldeado la variación observada. Es importante resaltar que, a pesar de que las distancias biológicas y la estructura de la población son principalmente investigadas en referencia a la deriva genética y la migración, algunas de las variables craneométricas incluidas en este análisis, están fuertemente condicionadas por la selección natural. Resultados obtenidos sobre las mismas poblaciones estudiadas aquí, pero a partir de técnicas de autocorrelación y autorregresión espacial (Fabra y Demarchi, 2011), en coincidencia con lo observado por otros investigadores (González-José, 2003; Bernal et al., 2006), indican que la morfología nasal está fuertemente influenciada por variables ambientales, tales como la altitud y la temperatura.
En síntesis, el presente estudio ha permitido estimar para la muestra de Córdoba a) una morfología craneofacial que la coloca, en función de la distancia biológica calculada, en una posición intermedia y relativamente diferenciada de otras poblaciones, hecho consistente con su ubicación geográfica y que resalta ese factor como condicionante de la evolución de la población, b) similitudes con Santiago del Estero y c) marcadas diferencias con San Luis, la otra muestra poblacional de las Sierras Centrales. Por otra parte, el análisis de Relethford-Blangero permite suponer para Córdoba un tamaño efectivo poblacional relativamente grande y la existencia de alto flujo genético con las poblaciones fuera de la región, mientras que la posición de San Luis sugiere la acción de fuerzas aleatorias actuando sobre una población pequeña.
Agradecimientos
Un agradecimiento especial a los museos que permitieron el acceso a las colecciones bajo su cuidado, en especial a Mirta Bonnin y Roxana Cattáneo (Museo de Antropología, F.F.yH. Universidad Nacional de Córdoba. Argentina), Myriam Tarragó, Inés Baffi, Mónica Berón y Claudia Aranda (Museo Etnográfico "Juan Bautista Ambrosetti". Universidad de Buenos Aires. Argentina), Héctor Pucciarelli (División Antropología. Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. Argentina), Paula Novellino (Museo Municipal de Historia de San Rafael. Mendoza. Argentina), Andrés Chazarreta (Museo de Ciencias Naturales y Antropológicas "Emilio y Duncan Wagner". Santiago del Estero. Argentina), Mónica Sans y Leonel C. Pérez (Departamento Arqueología. Facultad de Humanidades y Ciencias de la Educación. Universidad de la República. República Oriental del Uruguay), Arturo Toscano (Museo Nacional de Historia Natural y Antropología. Ministerio de Educación y Cultura. República Oriental del Uruguay), Alfredo Casaravilla Nodar (Museo Arqueológico Prof. Taddei. Canelones. República Oriental del Uruguay). Un agradecimiento a Paola Franco por la ilustración del mapa de la Figura 1. Finalmente, los autores desean agradecer las sugerencias y comentarios del editor y evaluadores, los cuales sin duda contribuyeron a mejorar la calidad del trabajo.
1Recientes estudios han establecido que la deformación de tipo circular es más frecuente en restos óseos anteriores a 3000 años A.P., asignables al Holoceno medio, mientras que la deformación tabular oblicua y tabular erecta se hallan presentes en restos asignables al Holoceno tardío, particularmente los individuos que presentan deformaciones craneanas del tipo tabular oblicua, poseen fechados radiocarbónicos cercanos al 2000 A.P. y las deformaciones de tipo tabular erecta, una edad radiocarbónica de 500 años A.P. aproximadamente (Berón y Baffi, 1996, 2004; Barrientos, 2001).
Literatura citada
1. Barrientos G. 2001. Una aproximación bioarqueológica al estudio del poblamiento prehispánico tardío del sudeste de la región pampeana. Intersecciones en Antropología (NS) 2:3-18.
2. Bernal V, Perez SI, González PN. 2006. Variation and causal factors of craniofacial robusticity in Patagonian huntergatherers from Late Holocene. Am J Hum Biol 18:748- 765. [ Links ]
3. Bernal V, Perez SI, González PN, Sardi ML, Pucciarelli HM. 2010. Spatial patterns and evolutionary processes in southern south America: a study of dental morphometric variation. Am J Phys Anthropol 142:95-104. [ Links ]
4. Berón M, Baffi E. 1996. Adscripción cronológica de dos individuos con deformación craneana circular en la Provincia de La Pampa, Argentina. Buenos Aires: IV Congreso de la Asociación Latinoamericana de Antropología Biológica. Segundas Jornadas Nacionales de Antropología Biológica. p 63.
5. Berón M, Baffi E. 2004. Variabilidad de las estructuras mortuorias en el Holoceno Tardío. Cuenca de los Lagos Posadas y Salitroso (Pcia. de Santa Cruz) y Área de Lihué Calel (Pcia. de La Pampa). En: Civalero MT, Fernández P, Guraieb A, editores. Contra viento y marea. Arqueología de la Patagonia. Buenos Aires: Instituto Nacional de Antropología y Pensamiento Latinoamericano. Sociedad Argentina de Antropología. p 387-402.
6. Buikstra J, Ubelaker D. 1994. Standards for data collection from human skeletal remains. Arkansas Archeological Survey Research Series No 44. Fayeteville: Arkansas.
7. Canals Frau S. 1944. El grupo huarpe-comechingón. Anales del Instituto de Etnografía Americana 5:9-41.
8. Cabrera AL. 1976. Regiones fitogeográficas argentinas. En: Kugler F, editor. Enciclopledia argentina de agricultura y jardinería. Buenos Aires: Editorial Acme.
9. Carnese FR, Mendisco F, Keyser C, Dejean CB, Dugoujon JM, Bravi CM, Ludes B, Crubézy E. 2010. Paleogenetical study of pre-columbian samples from Pampa Grande (Salta, Argentina). Am J Phys Anthropol 135:438-447. [ Links ]
10. Cocilovo JA. 1975. Estudio de dos factores que influencian la morfología craneana en una colección andina: el sexo y la deformación artificial. Revista del Instituto de Antropología 3(2):197-212.
11. Cocilovo JA. 1984. Una nueva aproximación al conocimiento de la población prehistórica de la provincia de Córdoba. Comechingonia 2(3):85-104.
12. Cocilovo JA, Di Rienzo J. 1984-1985. Un modelo biológico para el estudio del poblamiento prehispánico del territorio argentino. Correlación fenético-espacial. Relaciones de la Sociedad Argentina de Antropología 16:119-135.
13. Cocilovo JA, Neves WA. 1988-1989. Afinidades biológicas entre las poblaciones prehistóricas del Litoral de Brasil y de Argentina. Primera aproximación. Relaciones de la Sociedad Argentina de Antropología 17(2):31-56. [ Links ]
14. Dejean C, Keyser-Tracqui C, Crubézy E, Blandin P, Goicoechea AS, Avena SA, Carnese FR. 2004. Paleogenética en muestras esqueletales precolombinas de Pampa Grande, Prov. de Salta, Argentina. Venezuela: VIII Congreso de la Asociación Latinoamericana de Antropología Biológica.
15. Del Papa M. 2008. Estructuración espacial de la variación biológica humana en la República Argentina durante el Holoceno tardío final a través de los rasgos epigenéticos craneofaciales. Revista Argentina de Antropología Biológica 10(2):21-41.
16. Demarchi DA. 2009. Microsatélites, distancias genéticas y estructura de poblaciones nativas sudamericanas. Revista Argentina de Antropología Biológica 11(1):73-88.
17. Demarchi DA, Colantonio SE, Marcellino AJ. 1998. Population structure in the Gran Chaco and the uniqueness of the Ayoreo. Am J Hum Biol 10(1):120-121. [ Links ]
18. Demarchi DA, Panzetta-Dutari GM, Motran CC, Basualdo MA, Marcellino AJ. 2001a. Mitochondrial DNA haplogroups in Amerindian populations from the Gran Chaco. Am J Phys Anthropol 115:199-203.
19. Demarchi DA, Panzeta-Dutari GM, Colantonio S, Marcellino AJ. 2001b. Absence of the 9-bp deletion of mitocondrial DNA in prehispanic inhabitants of Argentina. Hum Biol 73:575-582. [ Links ]
20. Demarchi DA, Salzano F, Altuna ME, Fiegenbaum M, Hill K, Hurtado AM, Tsunetto L, Petzl-Erler ML, Hutz M. 2005. APOE polymorphism distribution among native Americans and related populations. Annals of Human Biology 32(3):351-365.
21. Dipierri JE, Alfaro E, Martínez-Marignac VL, Bailliet G, Bravi CM, Cejas S, Bianchi NO. 1998. Paternal directional mating in two Amerindian subpopulations located at different altitudes in northwestern Argentina. Hum Biol 70(6):1001-1010. [ Links ]
22. Fabra M. 2008a. Relaciones biológicas craneales entre poblaciones prehispánicas de la región central del territorio argentino y otras del Cono Sur sudamericano. Revista Anales de Arqueología y Etnología 61-62:123-145.
23. Fabra M. 2008b. Patrones geográficos de diferenciación craneofacial entre poblaciones de la región central del país y otras de Argentina: aportes desde la genética del paisaje. Revista del Museo de Antropología 1(1):13-24.
24. Fabra M. 2009. El poblamiento prehispánico de Córdoba: una interpretación a partir de evidencias bioantropológicas. Córdoba: Serie Tesis de Postgrado. Centro de Publicaciones Facultad de Filosofía y Humanidades. Universidad Nacional de Córdoba. Argentina.
25. Fabra M, Demarchi DA. 2009. Variabilidad craneofacial en poblaciones del sector austral de las Sierras Pampeanas: aportes desde la morfometría geométrica. Revista Relaciones de la Sociedad Argentina de Antropología 34:1-24.
26. Fabra M, Demarchi DA. 2011. Geographic patterns of craniofacial variation in pre-hispanic populations from the southern cone of South America. Human Biology 83(4):491-507. [ Links ]
27. Fabra M, Laguens A, Demarchi DA. 2005. Análisis intra e inter poblacional de rasgos craneanos no métricos en aborígenes prehispánicos del actual territorio de Córdoba. Revista Argentina de Antropología Biológica 7(2):47-65.
28. Fabra M, Laguens A, Demarchi DA. 2007. Human colonization of the central territory of Argentina: design matrix models and craniometric evidence. Am J Phys Anthropol 133(4):1060-1066.
29. Fabra M, Salega MS, González CV. 2009. Comportamiento mortuorio en poblaciones prehispánicas de la región austral de las Sierras Pampeanas durante el Holoceno. Revista Arqueología 15:165-186. [ Links ]
30. García A, Demarchi DA. 2006. Linajes parentales amerindios en poblaciones del norte de Córdoba. Revista Argentina de Antropología Biológica 8(1):57-71. [ Links ]
31. García A, Demarchi DA. 2009. Incidence and distribution of native american mtDNA haplogroups in Central Argentina. Human Biology 81(1):59-69. [ Links ]
32. González-José R. 2003. El poblamiento humano de la Patagonia: Análisis de la variación craneofacial en el contexto del poblamiento de América. Tesis Doctoral. Universidad de Barcelona. España.
33. González-José R, Dahinten S, Hernández M. 2001a. The settlement of Patagonia: a matrix correlation study. Human Biology 73(2):233-248.
34. González-José R, Dahinten S, Hernández M. 2001b. Craniometric variation and the settlement of the Americas: testing hypothesis by means or R-matrix and matrix correlation analysis. Am J Phys Anthropol 116:154-165.
35. González PN, Perez SI, Bernal V. 2010. Ontogeny of robusticity of craniofacial traits in modern humans: a study of South American populations. Am J Phys Anthropol 142:367-379. [ Links ]
36. Harpending HC, Ward RH. 1982. Chemical systematics and human populations. En: Nitecki M, editor. Biochemical aspects of evolutionary biology. Chicago: University of Chicago Press. p 213-256.
37. Laguens A. 1999. Arqueología del contacto hispano-indígena. Un estudio de cambios y continuidades en las Sierras Centrales de Argentina. Oxford: BAR International Series 801.
38. Laguens A. 2009. El poblamiento inicial del sector austral de las Sierras Pampeanas de Argentina desde la Ecología del Paisaje. Revista Anales de Arqueología y Etnología 61-62:67-107.
39. Laguens A, Demarchi DA, Fabra M. 2007. Un estudio arqueológico y bioantropológico de la colonización humana en el sector sur de las Sierras Pampeanas. Actas V y VI Jornadas de Investigadores en Arqueología y Etnohistoria del Centro-Oeste del país. Universidad Nacional de Río Cuarto. p 119-134.
40. Lalueza Fox CM, Hernández M, García Moro C. 1996. Craniometric analysis in groups from Tierra del Fuego/ Patagonia and the peopling of the south extreme of the Americas. Human Evolution 11(3-4):217-224. [ Links ]
41. Marcellino AJ. 1995-1996. Eran "ándidos" los aborígenes del agroalfarero de Icaño? Nueva contribución a la craneología de Santiago del Estero. Anales de Arqueologia y Etnologia 50-51:135-166.
42. Marcellino AJ, Colantonio S. 1993. Relaciones morfológicas de los aborígenes prehispánicos del territorio argentino. VII: La región serrana de Córdoba. Revista de la Junta Provincial de Historia de Córdoba 15.
43. Mendez M, Salceda S. 1995. Metric and non metric variants in prehistoric populations of Argentina. Rivista di Antropologia (Roma) 73:145-158. [ Links ]
44. Nores R, Demarchi DA. 2011. Análisis de haplogrupos mitocondriales en restos humanos de sitios arqueológicos de la provincia de Córdoba. Revista Argentina de Antropología Biológica 13(1):43-54.
45. Ossenberg NS. 1977. Congruence of distance measures based on cranial discrete traits, cranial measurements and linguistic- geographic criteria in five Alaskan populations. Brief Communication. Am J Phys Anthropol 47:93-98.
46. Pauro M, García A, Bravi CM, Demarchi DA. 2010. Distribución de haplogrupos mitocondriales alóctonos en poblaciones rurales de Córdoba y San Luis. Revista Argentina de Antropología Biológica 12(1):47-55.
47. Perez SI. 2006. El poblamiento holocénico del sudeste de la región pampeana: un estudio de morfometría geométrica craneofacial. Tesis Doctoral. Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. Argentina.
48. Perez SI, Bernal V, González PN. 2007. Morphological differentiation of aboriginal human populations from Tierra del Fuego (Patagonia): implications for South American peopling. Am J Phys Anthropol 133:1067-1079. [ Links ]
49. Politis GG, Messineo PG, Kaufmann CA. 2004. El poblamiento temprano de las llanuras pampeanas de Argentina y Uruguay. Complutum 15:207-224.
50. Relethford JH, Blangero J. 1990. Detection of differential gene flow from patterns of quantitative variation. Human Biology 62:5-25.
51. Relethford JH, Crawford MH, Blangero J. 1997. Genetic drift and gene flow in post-famine Ireland. Human Biology 69:443-465. [ Links ]
52. Rothhammer F, Acuña M, Llop E. 1988-1989. La población de Sudamérica: nuevos aportes basados en el análisis de cráneos arcaicos y frecuencias génicas de aborígenes contemporáneos. Relaciones de la Sociedad Argentina de Antropología 17(2):19-30.
53. Sardi ML, Ramirez Rozzi F, González-José R, Pucciarelli HM. 2005. South Amerindian craniofacial morphology: diversity and implications for Amerindian evolution. Am J Phys Anthropol 128:747-756. [ Links ]
54. Varela HH, Cocilovo JA. 2002. Genetic drift and gene flow in a prehistoric population of the Azapa Valley and coast, Chile. Am J Phys Anthropol 118:259-267.
55. Varela HH, O´Brien TG, Cocilovo JA. 2008. The genetic divergence of prehistoric populations of the south-central Andes as established by means of craniometric traits. Am J Phys Anthropol 137:274-282. [ Links ]

