










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkRevista argentina de antropología biológica
versão On-line ISSN 1514-7991
Rev Arg Antrop Biol vol.14 no.1 La Plata dez. 2012
TRABAJOS ORIGINALES
Análisis de procrustes y el estudio de la variación morfológica
Sebastián Torcida1* y S. Ivan Perez2
1Grupo de Estadística. ECOSISTEMAS. Facultad de Ciencias Exactas. UNCPBA. Tandil. Argentina
2División Antropología. Museo de La Plata. Universidad Nacional de La Plata. CONICET. La Plata. Argentina
Financiamiento: CONICET. UNLP. UNCPBA.
*Correspondencia a: Sebastián Torcida. Grupo de Estadística. ECOSISTEMAS. Facultad de Ciencias Exactas. Campus. UNCPBA. 7000 Tandil. Argentina. E-mail: unisebas@gmail. com
Recibido: 04 Septiembre 2011
Aceptado: 02 Mayo 2012
Resumen
El estudio de la variación morfológica ha ocupado un lugar central en la antropología, así como en el resto de las ciencias naturales, a partir de su surgimiento en el siglo XVIII. Desde un punto de vista cuantitativo, el estudio de la variación morfológica busca obtener información acerca del tamaño y la forma de una estructura. Tradicionalmente, estas propiedades han sido estudiadas a través de variables lineales tales como "ancho", "longitud", "altura" y empleando métodos estadísticos multivariados. En las décadas del 1980 y 1990 las técnicas cuantitativas de análisis de la variación morfológica fueron revolucionadas por el desarrollo y la aplicación de un método para estudiar coordenadas cartesianas de puntos anatómicos: el Análisis de Procrustes. Desde entonces, este método ha ganado relevancia en el campo de la antropología biológica. El objetivo de este artículo es revisar los principios del Análisis de Procrustes aplicado a coordenadas de puntos anatómicos y discutir el empleo de diferentes versiones del mismo en bioantropología. El texto está ordenado como sigue: en primer lugar revisamos diversas nociones básicas involucradas en los estudios morfométricos; entre ellos, los conceptos de tamaño y forma. En segundo lugar discutimos los principios del Análisis de Procrustes y señalamos las diferencias entre sus dos versiones más importantes: la superposición por cuadrados mínimos y la superposición robusta por medianas repetidas. Finalmente, mostramos y discutimos algunos ejemplos de la aplicación de estos métodos en el campo de la antropología biológica.
Palabras clave: Morfometría geométrica; Puntos anatómicos; Análisis de Procrustes por cuadrados mínimos y por medianas repetidas
Abstract
The study of morphological variation has occupied a central place in anthropology, as well as in the rest of the natural sciences, from its emergence in the eighteenth century. From a quantitative point of view, the study of morphological variation seeks information about the size and shape of a structure. Traditionally, these properties have been studied using linear variables such as "width", "length", "height" and using multivariate statistical methods. In the eighties and nineties, the quantitative techniques for analyzing morphological variation were revolutionized by the development and implementation of a method to study the Cartesian coordinates of anatomical landmarks, the Procrustes analysis. Since then, this method has gained prominence in the field of biological anthropology. The aim of this paper is to revise the principles of the Procrustes analysis applied to coordinates of anatomical landmarks and discuss the use of its different versions in bioanthropology. The text is organized as follows: first, there is a review of several basic concepts involved in the morphometric studies, among them are the concepts of size and shape. Secondly, the principles of the Procrustes analysis are discussed, and the differences between its two major versions, the least-squares fit and the repeated-medians robust fit, are pointed out. Finally, some examples of the application of these methods to the field of biological anthropology are shown and discussed.
Key words: Geometric morphometrics; Landmarks; Least squares and repeated-medians Procrustes Analysis
La variedad de formas encontradas en los organismos ha suscitado el interés de los naturalistas desde milenios atrás (Darwin, 1859; Thompson, 1917). En particular, los estudios antropológicos se han centrado en la descripción y comparación de la variación morfológica presente en los humanos modernos y las especies estrechamente emparentadas (e.g., primates no humanos y homínidos; Boas, 1912; Le Gros Clark, 1949; Beals et al., 1984; Relethford, 2004). Entre estos taxa, las diferencias morfológicas son principalmente cuantitativas: por ejemplo, el cráneo humano y el de especies primates relacionadas está conformado por el mismo conjunto de elementos óseos, pero difieren cuantitativamente en su tamaño y su forma (Fleagle, 1999). Por lo tanto, el estudio de la variación morfológica se ha basado fundamentalmente en el empleo de métodos cuantitativos que pretenden caracterizar el tamaño y la forma de los organismos: las llamadas técnicas morfométricas (Reyment, 1985; Bookstein, 1991). En consecuencia, la morfometría ha sido una de las principales aproximaciones para el estudio de la variación morfológica entre individuos, poblaciones y/o especies de primates y particularmente entre poblaciones de humanos modernos (Howells, 1969; Bookstein et al., 2004; Slice, 2005, 2007).
Los primeros estudios morfométricos combinaron el empleo de medidas lineales entre puntos anatómicos con técnicas estadísticas uni y multivariadas para el análisis de las diferencias de tamaño y de forma (Bookstein, 1996; Slice, 2007). Este enfoque, actualmente denominado morfometría tradicional (Marcus, 1990; Rohlf y Marcus, 1993), alcanzó gran desarrollo en la segunda mitad del siglo XX por la creciente disponibilidad de computadoras y de programas para análisis estadísticos multivariados. A finales del siglo XX se afianzó el enfoque cuantitativo que hoy denominamos morfometría geométrica o síntesis morfométrica (Bookstein, 1996; Adams et al., 2004; Mitteroecker y Gunz, 2009). Se definió como la fusión empírica de la geometría con la biología (Bookstein, 1982) y tuvo como propósito extraer información de la geometría de una forma biológica para propósitos comparativos particulares. Uno de los métodos más extendidos hasta la actualidad dentro de los enfoques geométricos es el que emplea el Análisis de Procrustes (AP) para obtener variables de tamaño y de forma a partir de las coordenadas cartesianas de puntos anatómicos (Gower, 1975; Siegel y Benson, 1982; Rohlf y Slice, 1990) y analizarlas posteriormente mediante el empleo de técnicas estadísticas multivariadas (Bookstein, 1996; Mitteroecker y Gunz, 2009).
Aunque el estudio de la variación morfométrica entre individuos, poblaciones y/o especies de primates ha sido uno de los objetivos centrales de la antropología biológica, hasta el presente no se cuenta con revisiones matemáticamente rigurosas y objetivas del AP y de las técnicas estadísticas multivariadas involucradas. En general, las revisiones se han polarizado entre aquellas que defienden la aproximación denominada Síntesis Morfométrica -en la que se utilizan el AP por cuadrados mínimos en combinación con técnicas estadísticas multivariadas que efectúan rotaciones rígidas del espacio euclidiano, tales como Componentes Principales y Regresiones Multivariadas (e.g., Bookstein, 1996; Mitteroecker y Gunz, 2009)- y las que critican cualquier técnica morfométrica basada en el AP (e.g., Richtsmeier et al., 2002). El objetivo central de este artículo es revisar los fundamentos de diferentes versiones del AP aplicado a coordenadas de puntos anatómicos para obtener las variables de tamaño y de forma utilizadas en la morfometría geométrica y discutir su aplicación a problemas bioantropológicos. El texto está ordenado como sigue: en primer lugar, revisamos diversas nociones básicas involucradas en los estudios morfométricos; entre ellos, los conceptos de tamaño y forma. A continuación discutimos los fundamentos del AP aplicado a coordenadas de puntos anatómicos y señalamos las diferencias entre sus dos versiones más importantes: la superposición por cuadrados mínimos y la superposición robusta por medianas repetidas. Finalmente, mostramos y discutimos algunos ejemplos de la aplicación de ambos métodos en el campo de la antropología biológica.
Landmarks, datos morfométricos, tamaño y forma
Los landmarks pueden ser definidos como puntos anatómica o geométricamente homólogos entre estructuras. Bookstein (1991) distinguió tres tipos de landmarks (Fig. 1A):
- Tipo I: aquellos puntos que cuentan con mayor evidencia biológica de su homología; por ejemplo, un patrón local de yuxtaposición de tejidos o una pequeña sección de características histológicas inusuales;
- Tipo II: puntos cuya homología sólo se sostiene con evidencia geométrica y no histológica; por ejemplo, puntos de máxima curvatura;
- Tipo III: puntos con al menos una coordenada ambigua; por ejemplo, los extremos de un diámetro máximo o el punto inferior de una concavidad. Estos landmarks caracterizan más de una región de la estructura, condicionando cualquier interpretación geométrica o biológica que se haga sobre ellos. Debido a la naturaleza imprecisa de los landmarks de tipo III, Bookstein revisó en 1997 su clasificación de 1991 y denominó semilandmarks a éste último grupo (Bookstein, 1997).

Fig. 1. Representación de los landmarks tipo I, II y III (A) y distancias lineales (B; M1 y M2).
La forma de un objeto ha sido caracterizada y estudiada en la morfometría tradicional a través del registro de medidas lineales entre landmarks (Howells, 1973; Marcus, 1990; Fig. 1B). Un enfoque alternativo se basa en el registro y el análisis de las coordenadas cartesianas de los landmarks, en dos o tres dimensiones (Bookstein, 1991; Fig. 1A). El argumento principal para justificar el uso de las coordenadas de los landmarks en reemplazo de las tradicionales distancias lineales entre ellos lo proporciona el hecho de que en muchos casos éstas últimas no consiguen capturar la configuración espacial específica de la estructura subyacente (Adams et al., 2004; Slice, 2007).
Una vez registradas las coordenadas cartesianas de los landmarks de una estructura -o las distancias lineales entre ellos- es posible estudiar su forma y su tamaño (Fig. 2). Todas las técnicas morfométricas desarrolladas con posterioridad a la década de 1970 (Bookstein, 1989) analizan estos dos conceptos de manera independiente. La idea de separar la forma del tamaño ha sido ampliamente discutida en la antropología y en la biología. Muchos investigadores consideran que la forma es más relevante que el tamaño en los análisis taxonómicos y de anatomía funcional (Corruccini, 1987), tanto para niveles subespecíficos (Sneath y Sokal, 1973) como supraespecíficos (Relethford, 1984), señalando que aunque forma y tamaño son heredables en la mayoría de los grupos biológicos, el tamaño tiene una mayor variabilidad entre grupos (Corruccini, 1987) y ello enmascara la variación de forma en los análisis multivariados (Bookstein, 1989).

Fig. 2. Representación del cambio en forma y tamaño entre dos objetos geométricos, un cuadrado y un rectángulo. Se puede observar como el cambio en forma (form) es una combinación del cambio en tamaño (size) y "forma" (shape).
En el análisis basado en coordenadas cartesianas de landmarks (Bookstein, 1991), el tamaño se define a partir del tamaño del centroide (s): la raíz cuadrada de la suma de las distancias cuadradas de todos los puntos respecto a su centro de gravedad (Bookstein, 1989, 1991; Fig. 3).

Utilizando notación matricial, el tamaño del centroide se puede calcular como:

donde tr representa la traza de una matriz, Id es una matriz identidad y P una matriz con coeficientes ![]()
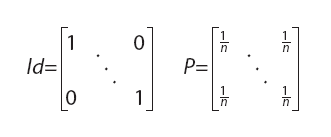
ambas de tamaño nxn (siendo n el número de landmarks) y donde X es la matriz de tamaño nxq (q=2 ó 3, en general) de coordenadas de los landmarks (Rohlf, 1990; Rohlf y Slice, 1990; Bookstein, 1991). La conveniencia de utilizar el tamaño del centroide como variable de tamaño en los análisis geométricos radica en que es la única alternativa que no se correlaciona con la forma cuando el modelo nulo de alometría es verdadero (Bookstein, 1991; ésta falta de correlación sólo es esperable cuando se efectúa una superposición por cuadrados mínimos).

Fig. 3. Representación geométrica de las distancias Euclideanas involucradas en el cálculo del tamaño del centroide de una configuración de 4 coordenadas cartesianas en 2 dimensiones.
Una vez definido el tamaño, la forma es el espacio de todas las proporciones de las variables medidas con respecto a una única variable de tamaño X (Mosimann y James, 1979; Bookstein, 1989). Es importante notar que el tamaño de una estructura puede expresarse como un valor absoluto; la forma en cambio, sólo puede expresarse en términos relativos.
La noción de forma utilizada en la morfometría geométrica es la propuesta por Mosimann, que hace referencia a la invariabilidad en diferentes escalas físicas y que fue reformulada en coordenadas cartesianas por David Kendall (1984). A diferencia de las variables lineales, un conjunto de coordenadas cartesianas no sólo tiene información sobre el tamaño y la forma de los objetos, sino también sobre la posición y la orientación relativa de las coordenadas (Fig. 4). Así, la forma puede ser definida como la información presente en un conjunto de coordenadas después que la localización, la escala y la orientación son excluidas (Bookstein, 1991, 1996), generalmente por medio de un Análisis de Procrustes (AP; Gower, 1975; Rohlf, 1990; Rohlf y Slice, 1990).
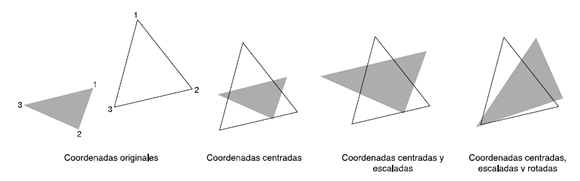
Fig. 4. Representación gráfica de los tres pasos del Análisis de Procrustes (el caso particular del ajuste por cuadrados mínimos) usando dos triángulos. Las coordenadas resultantes después del análisis son denominadas coordenadas de forma Procrustes.
Análisis de Procrustes
En la mitología griega, Procrustes era un posadero muy particular que obligaba a sus huéspedes a acostarse en uno de los dos lechos de los que disponía forzando a que se ajustaran perfectamente a ellos: a los que eran bajos, los acomodaba en una cama larga y los estiraba cruelmente; a los que eran altos, los acomodaba en una cama corta y les cortaba las piernas. Inspirado en esta leyenda, el AP describe un conjunto de herramientas matemáticas que permiten comparar dos configuraciones de puntos homólogos provenientes de dos variantes de la misma entidad: dos individuos dentro de una población, dos especies, etc.
Específicamente, el objetivo del AP es determinar si representaciones alternativas de los mismos n puntos exhiben diferentes relaciones internas entre ellos. Cabe aquí recordar que existen operaciones que transforman un ordenamiento sin alterar las relaciones internas entre los puntos representados (Fig. 4), a saber:
- desplazar todos los puntos a una distancia fija, a lo largo de una determinada dirección. Esto constituye una traslación de la configuración;
- una dilatación o una contracción fija de todos los puntos a lo largo de las direcciones que unen a cada uno de ellos con el centroide de la configuración. Se trata de un cambio de escala uniforme; - desplazar todos los puntos usando un ángulo fijo alrededor de un eje. Esto se realiza a través de una rotación/reflexión de la configuración.
El AP busca determinar entonces qué combinación de un cambio de escala y de movimientos rígidos (la rotación/reflexión y la traslación lo son) permite superponer de manera óptima ambas configuraciones, dejando a la vista las verdaderas diferencias entre ambas representaciones (Fig. 4).
El AP resulta particularmente apropiado para estudiar las diferencias de forma entre dos estructuras representadas por configuraciones de landmarks (Rohlf, 1990; Rohlf y Slice, 1990; Dryden y Mardia, 1998; Adams et al., 2004; Mitteroecker y Gunz, 2009) y existen diferentes versiones del método. El AP por cuadrados mínimos -también conocido como superposición Procrustes clásica- y el AP robusto por medianas repetidas son las dos estrategias más utilizadas y están basadas en criterios de alguna manera contrastantes (Rohlf, 1990; Rohlf y Slice, 1990; Zelditch et al., 2004). Es claro que dos configuraciones diferentes rara vez podrán ser superpuestas "perfectamente", de modo que distintos métodos de superposición arrojarán diferentes resultados, en general.
La superposición Procrustes clásica busca minimizar la suma de las diferencias al cuadrado entre todos los pares de landmarks. Geométricamente, esto se consigue promediando la falta de ajuste de todos los puntos. Las soluciones analíticas para este enfoque fueron obtenidas por Gower (1971, 1975) y Sibson (1978). Debido a que los cálculos del tipo "promedio" suelen ser inestables, las verdaderas diferencias de forma entre ambas configuraciones pueden no quedar representadas adecuadamente por el resultado de esta superposición. A pesar de ello, la relativa simplicidad matemática del método hace que su uso sea ampliamente difundido.
Cuando dos configuraciones de landmarks exhiben diferencias de forma localizadas, la comparación más efectiva consiste en superponer perfectamente los puntos o partes para los que no existen cambios. Ello permite que las diferencias se detecten y queden evidenciadas a través de la falta de ajuste y los grandes residuales de los restantes puntos ó partes. En el ámbito de la estadística, el adjetivo robusto o resistente alude a la capacidad de un método para identificar qué patrón de comportamiento en los datos es minoritario o atípico y proceder en consecuencia.
Resumiendo: cuando las diferencias de forma tienen lugar en un conjunto pequeño de landmarks, la superposición resultante debería mostrar que en la mayoría de los puntos no existen cambios.
Existen diversas técnicas robustas de AP (Dryden y Walker, 1999; Larsen, 2008); entre ellas, el AP por medianas repetidas (Siegel y Benson, 1982) goza del mayor breakdown value: 50% (cf. Siegel, 1982), indicando que el método es capaz de obtener el ajuste óptimo aún si sólo el 50% de los landmarks pueden superponerse perfectamente. Se basa en cálculo de medianas repetidas para obtener la superposición que mejor ajusta a la mayoría de los puntos, con el objetivo de que grandes faltas de ajuste en sólo unos pocos landmarks tengan un alto impacto en el resultado. Así, el resultado de una superposición por medianas repetidas es más robusto que el de una superposición clásica, porque refleja con mayor precisión las eventuales diferencias de forma localizadas entre dos estructuras.
En lo que sigue, abordaremos el problema de comparar dos configuraciones de n landmarks en p=2 dimensiones empleando un mínimo de notación matemática; las fórmulas que describen el funcionamiento de los métodos sólo requieren un conocimiento básico de álgebra lineal. Existen herramientas análogas para p=3 dimensiones (Slice, 1996), pero nos limitaremos principalmente a 2 dimensiones por simplicidad.
Superposición Procrustes por cuadrados mínimos (clásica)
Consideremos dos matrices X e Y de tamaño nx2 cuyas filas corresponden a las coordenadas cartesianas de las mismas n entidades (landmarks) en p=2 dimensiones:
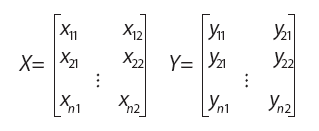
En lo que sigue se asumirá que ambas matrices tienen su centroide en el origen; esto se consigue premultiplicando a cada una de ellas por la matriz Id–P, donde Id es la matriz identidad y P una matriz de coeficientes ![]() , ambas de tamaño nxn:
, ambas de tamaño nxn:
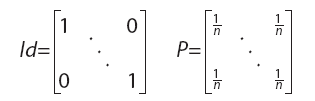
De acuerdo con el espíritu de la leyenda mitológica y las consideraciones previas, se pretende hallar el vector traslación t=[t1 t2], el factor de escala ρ>0 y la matriz de rotaciónreflexión R (matriz ortogonal) que minimicen la suma de las diferencias cuadradas entre los puntos de una configuración y la versión rotada, escalada y trasladada de la configuración restante:
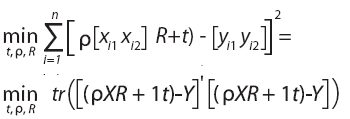
donde "tr" denota la traza, 1 es un vector columna de n unos y X´ es la traspuesta de la matriz X.
Si USV'=Y'X es la descomposición en valores singulares de la matriz Y'X, donde U,V son matrices ortogonales de tamaño pxp y S=diag (s11, s22, … , spp) una matriz diagonal de tamaño pxp con elementos diagonales no negativos y decrecientes (s11 ≥ s22 ≥ … ≥ spp ≥ 0), se puede probar que los valores de t, r y R que minimizan la expresión anterior son:

Aplicando estas transformaciones sobre la matriz X se obtiene su superposición Procrustes por cuadrados mínimos respecto a Y. Las coordenadas transformadas de los landmarks en X tienen suma mínima de desviaciones cuadradas con respecto a las correspondientes coordenadas en Y.
Superposición Procrustes por medianas repetidas (robusta)
Una diferencia importante entre ésta estrategia y la anterior es que, debido a su formulación matemática, el AP por medianas repetidas sólo considera rotaciones para minimizar las diferencias de orientación entre ambas configuraciones. Por este motivo suele llevarse a cabo previamente un AP clásico, que efectuará una reflexión cuando resulte necesario y permitirá al mismo tiempo una estimación adecuada del ángulo de la rotación robusta (Siegel y Benson, 1982).
Se considera en primer lugar la rotación. Cada elemento de una matriz de rotación en dos dimensiones queda determinada por un único parámetro: el ángulo de rotación θ, medido en radianes. Así, la matriz

efectúa una rotación de ángulo θ antihoraria y alrededor del origen de coordenadas en el plano cartesiano.
La construcción de la rotación por medianas repetidas se desarrolla en etapas: inicialmente, para cada par de landmarks distintos en una de las configuraciones se determina el rayo (segmento dirigido) que los une. Se calcula entonces el ángulo θij∈[-π,π] que debe rotarse el rayo que parte del landmark Li =[xi1 xi2] y llega al landmark Lj =[xj1 xj2] (1≤i≠j≤n) para que resulte perfectamente alineado con el rayo homólogo que parte del landmark LYi =[yi1 yi2] y llega al landmark LYj =[yj1 yj2]:

donde arc cos es la función arco-coseno, · denota el producto escalar de vectores y ║║representa la norma de un vector. Por cada par de rayos homólogos se define así una estimación inicial del ángulo (y por lo tanto de la matriz) de la rotación por medianas repetidas. El rango seleccionado para medir los ángulos θ
Una vez obtenidos los 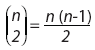 ángulos θij (1≤i≠j≤n), se calcula su mediana repetida:
ángulos θij (1≤i≠j≤n), se calcula su mediana repetida:
![]()
a partir de la cual se define finalmente la matriz de rotación por medianas repetidas
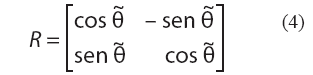
De manera similar, por cada par de landmarks homólogos se define un estimador inicial del factor de escala ρij como el factor que hace que el segmento que conecta los landmarks i y j en la estructura X resulte de la misma longitud que el segmento homólogo en la estructura Y:
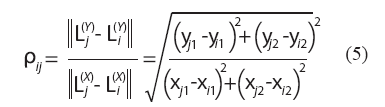
El factor de escala por medianas repetidas se consigue aplicando las medianas repetidas sobre estas![]() cantidades:
cantidades:
![]()
Es interesante notar que la rotación y el cambio de escala del método de medianas repetidas se obtienen de forma independiente uno del otro. Sólo después que ambos han sido determinados, el vector traslación robusto puede estimarse inicialmente con cada fila de la matriz residual

A lo largo de cada columna de la matriz T se calcula la mediana (simple) para arribar al vector traslación robusto

En resumen, la superposición por medianas repetidas de las configuraciones X e Y se consigue transformando la matriz X en
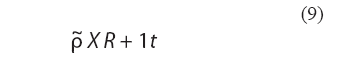
donde 1 es un vector columna de n unos.
Generalización de la superposición Procrustes para más de dos configuraciones en 2D y 3D
Existe una generalización directa del AP clásico para el ajuste simultáneo de más de dos configuraciones de landmarks en 2D y 3D, que consiste en la superposición iterativa de cada configuración contra la configuración media o consenso (Gower, 1975; Rohlf y Slice, 1990). El algoritmo se detiene cuando ya no existen cambios en los valores de la suma de cuadrados residual (i.e., la distancia Procrustes), la medida de ajuste natural que resulta minimizada.
Rohlf y Slice (1990) propusieron una extensión del AP por medianas repetidas para permitir el ajuste de más de dos configuraciones de puntos en 2D de manera análoga, superponiendo iterativamente cada configuración contra la configuración mediana. Sin embargo, el criterio utilizado para detener el algoritmo en este caso es exclusivamente la falta de cambio en la configuración mediana, ya que el método no cuenta con una medida de ajuste asociada naturalmente.
Finalmente, cabe mencionar que la extensión del AP por medianas repetidas para dos configuraciones en 3D es relativamente reciente (Slice, 1996) y la ausencia de un criterio de optimización natural para el ajuste simultáneo de más de dos configuraciones -en 2D y 3D- hace que este problema constituya todavía materia de debate, a la espera de nuevos y superadores resultados.
Ejemplos
Se presentan a continuación un ejemplo geométrico artificial y un ejemplo biológico de aplicación morfométrica del AP por cuadrados mínimos y por medidas repetidas para ilustrar sus diferencias.
Ejemplo 1
En el ejemplo artificial, dos configuraciones de landmarks con cuatro coordenadas cartesianas fueron generadas: un rectángulo (configuración a; Fig. 5A) y un polígono similar que resulta al desplazar hacia abajo el landmark 4 (configuración b; configuración a; Fig. 5A). La superposición por cuadrados mínimos de estas dos configuraciones produce, como es de esperar, un ajuste global; las coordenadas resultantes sugieren que las mayores diferencias de forma están asociadas a los landmarks 2 y 4 (Fig. 5B). La superposición por medianas repetidas en cambio, produce un ajuste en el que los landmarks 1, 2 y 3 se superponen perfectamente y las diferencias de forma se concentran únicamente en el landmark 4 (Fig. 5C). El resultado del ajuste por medianas repetidas refleja así las diferencias en forma existentes entre las dos configuraciones generadas artificialmente y coincide con nuestras nociones elementales e intuitivas de geometría.

Fig. 5. Dos configuraciones de landmarks con cuatro coordenadas cartesianas generadas artificialmente (A), un rectángulo (configuración a) y un polígono similar logrado al desplazar hacia abajo el landmark 4 (configuración b), y sus superposiciones obtenidas empleando los métodos de cuadrados mínimos (B) y medianas repetidas (C).
Ejemplo 2
En nuestro segundo ejemplo, estudiamos los cambios de forma del cráneo entre dos individuos correspondientes a diferentes estadios ontogenéticos de una misma especie (Alouatta caraya), un subadulto (configuración a) y un adulto mayor (configuración b). Con tal fin se registraron 38 puntos anatómicos o landmarks (Fig. 6A). La superposición por cuadrados mínimos, graficada en la Figura 6B, muestra la falta de ajuste global de los 38 landmarks, aunque existe mayor variación en los landmarks de la región de la calota. La superposición por medianas repetidas es graficada en la Figura 6C, donde puede apreciarse que existe un mayor ajuste en los landmarks de la base del cráneo y una falta de ajuste más marcado en la región facial (Fig. 7). En términos generales, ambas superposiciones resultan consistentes; sin embargo, un análisis detallado permite detectar que las diferencias de forma identificadas por la superposición por medianas repetidas en la región facial son mucho más pronunciadas (Fig. 7), algo esperable por el crecimiento de esta región durante la ontogenia de la especie.

Fig. 6. Superposición de dos configuraciones landmarks del cráneo de dos individuos correspondientes a diferentes estadios ontogenéticos de una misma especie (A), un subadulto (configuración a) y un adulto mayor (configuración b), empleando los métodos de cuadrados mínimos (B) y medianas repetidas (C).
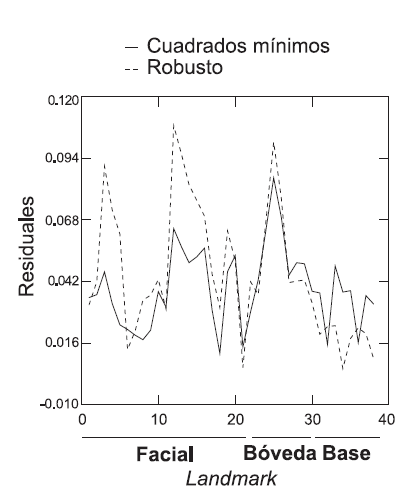
Fig. 7. Residuos Procrustes resultantes de la superposición de las configuraciones mostradas en la Figura 6 empleando los métodos de cuadrados mínimos y medianas repetidas.
Consideraciones finales
Las dos versiones del AP presentadas aquí para ajustar dos configuraciones de landmarks, así como las que permiten comparar n configuraciones (Rohlf y Slice, 1990), integran un conjunto de herramientas destinadas al estudio cuantitativo de la variación morfológica que ha sido denominado morfometría geométrica (Rohlf y Marcus, 1993). Al describir las variaciones morfológicas empleando las coordenadas cartesianas de landmarks homólogos, la morfometría geométrica permite comprender los cambios en las relaciones espaciales/estructurales entre los landmarks. Es importante señalar, además, que el método de superposición elegido determina las variables de forma que se generarán y que se emplearán posteriormente en los análisis multivariados.
El AP clásico es el método generalmente preferido para obtener una primera superposición óptima de dos configuraciones de landmarks después de la digitalización y generar de ese modo un conjunto de variables de forma que pueden ser utilizadas para los subsecuentes análisis estadísticos multivariados (Zelditch et al., 2004; Mitteroecker y Gunz, 2009). Una de las razones esgrimidas para su uso habitual es que el AP por cuadrados mínimos tiene la ventaja de estar enmarcado en la teoría de Kendall de la forma geométrica (Kendall, 1984; Zelditch et al., 2004) y por lo tanto es típicamente el preferido en los análisis estadísticos multivariados (Slice, 2005).
Los autores discrepamos con esa preeminencia automática, que tiene consenso mayoritario entre los especialistas en técnicas de morfometría geométrica, por varias razones. En primer lugar, Slice (2001) mostró que el ajuste por cuadrados mínimos utilizado en estudios biológicos -descrito en esta revisión- es sólo una aproximación al espacio de formas de Kendall. En segundo lugar, los análisis estadísticos multivariados son realizados sobre las coordenadas de landmarks después que las mismas han sido superpuestas por cuadrados mínimos y proyectadas en un espacio Euclidiano, por lo que las coordenadas de landmarks en el espacio de Kendall en realidad, nunca son directamente empleadas en los análisis morfométricos comparativos. Por último, las coordenadas de landmarks superpuestas por medianas repetidas pueden ser después proyectadas sobre un espacio Euclidiano y/o ser analizadas por técnicas estadísticas robustas (e.g., Escalamiento Multidimensional No-Métrico), de manera análoga a lo que se hace en el AP por cuadrados mínimos. ¿En qué medida el AP por medianas repetidas genera un espacio de forma significativamente diferente del generado por el AP de cuadrados mínimos? Responder sólidamente a este interrogante requiere mayor investigación. Por lo tanto, la falta de argumentos teóricos contundentes e independientes para considerar al AP por cuadrados mínimos mejor que el AP por medianas repetidas nos permite pensar que puede ser útil emplear criterios empíricos para evaluar la elección de la mejor alternativa en cada caso particular.
En ese contexto y tal como pudo apreciarse en los ejemplos, un aspecto relevante del AP por cuadrados mínimos es que distribuye las diferencias de forma de manera homogénea entre todos los landmarks (Figs. 5B, 6B y 7). Esto podría parecer razonable si a priori se sabe que tal comportamiento es aplicable a un conjunto de datos (Bookstein, 1991). Sin embargo, en muchos estudios morfométricos se espera que la variación de forma entre individuos o especies se concentre en regiones específicas de las estructuras estudiadas (e.g., en estudios biomecánicos y ontogenéticos; Slice, 1996; Zelditch et al., 2004). En tales casos, el ajuste por medianas repetidas sería el método más adecuado (Siegel y Benson, 1982; Slice, 1996) ya que el AP clásico podría "enmascarar" o "diluir" las verdaderas diferencias de forma.
En conclusión, para decidir qué versión del AP es más conveniente utilizar con nuestros datos y en análisis estadísticos multivariados posteriores, es útil contar con información tanto de la biología de la estructura como del problema que será estudiado (Slice, 1996). En la práctica, un AP por medianas repetidas se aplica habitualmente luego de un AP clásico por razones técnicas (que hemos señalado), de modo que en el peor caso ambos métodos no diferirán demasiado. Sin embargo, en muchas situaciones el AP por medianas repetidas termina actuando como un "calibre más fino", que deja en evidencia -en caso de existir- la variación localizada de forma. En general, un análisis exploratorio de los resultados proporcionados por ambos métodos suele ser de gran utilidad.
Agradecimientos
Queremos agradecer a Paula Gonzalez y a dos revisores anónimos por sus valiosos comentarios que permitieron mejorar el artículo.
Literatura citada
1. Adams DC, Rohlf FJ, Slice DE. 2004. Geometric morphometrics: ten years of progress following the ‘revolution'. Ital J Zool 71:5-16.
2. Beals KL, Smith CL, Dodd SM. 1984. Brain size, cranial morphology, climate, and time machines. Curr Anthropol 25:301-330. [ Links ]
3. Boas F. 1912. Changes in bodily form of descendants of immigrants. Am Anthropol 14:530-563.
5. Bookstein FL. 1982. Foundations of morphometrics. Annu Rev Ecol Syst 13:451-470.
6. Bookstein FL. 1989. "Size and shape": a comment on semantic. Syst Zool 38:173-180.
7. Bookstein FL. 1991. Morphometric tools for landmark data: geometry and biology. Cambridge: Cambridge University Press.
8. Bookstein FL. 1996. Biometrics, biomathematics and the morphometric synthesis. Bull Math Biol 58:313-365.
9. Bookstein FL. 1997. Landmark methods for forms without landmarks: Localizing group differences in outline shape. Med Image Anal 1:225-243.
10. Bookstein FL, Slice DE, Gunz P, Mitteroecker P. 2004. Anthropology takes control of morphometrics. Coll Antropol 28:121-132.
11. Corruccini RS. 1987. Shapes in morphometrics: comparative analyses. Am J Phys Anthropol 73:289-303.
12. Darroch JN, Mosimann JE. 1985. Canonical and principal component of shape. Biometrika 72:241-252. [ Links ]
13. Darwin C. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: John Murray.
14. Dryden IL, Mardia KV. 1998. Statistical shape analysis. New York: John Wiley & Sons.
15. Dryden IL, Walker G. 1999. Highly resistant regression and object matching. Biometrics 55:820-825. [ Links ]
16. Fleagle JG. 1999. Primate adaptation and evolution. New York: Academic Press.
17. Gower JC. 1971. Statistical methods of comparing different multivariate analyses of the same data. En: Hodson FR, Kendall DG, Tautu P, editores. Mathematics in the archaeological and historical sciences. Edinburgh: Edinburgh University Press. p 138-149.
18. Gower JC. 1975. Generalized Procrustes analysis. Psychometrika 40:33-51.
19. Howells W. 1969. The use of multivariate techniques in the study of skeletal populations. Am J Phys Anthropol 31:311-314.
20. Howells W. 1973. Cranial variation in man. A study by multivariate analysis of patterns of difference among recent human populations. Papers of Peabody Museum of Archaeology and Ethnology. Cambridge MA: Harvard University.
21. Kendall DG. 1984. Shape-manifolds, Procrustean metrics and complex projective spaces. Bull Lond Math Soc 16:81-121.
22. Larsen R. 2008. L1 Generalized Procrustes 2D Shape Alignment. Journal of Mathematical Imaging and Vision 31(2-3):189-194.
23. Le Gros Clark WE. 1949. History of the primates. London: British Museum (Natural History).
24. Marcus LF. 1990. Traditional morphmetrics. En: Rohlf FJ, Bookstein FL, editores. Proceedings of the Michigan morphometrics Workshop. Special Publication Number 2. Ann Arbor. Michigan: The University of Michigan Museum of Zoology. p 77-122.
25. Mitteroecker P, Gunz P. 2009. Advances in geometric morphometrics. Evol Biol 36:235-247.
26. Mosimann JE. 1970. Size allometry: size and shape variables with characterizations of the lognormal and generalized gamma distributions. J Am Stat Assoc 65:930-945.
27. Mosimann JE. 1988. Size and shape analysis. En: Kotz S, Johnson NL, editores. Encyclopedia of statistical science. New York: Wiley. p 497-508.
28. Mosimann JE, James FC. 1979. New statistical methods for allometry with application to Florida red-winged blackbirds. Evolution 33:444-459. [ Links ]
29. Relethford JH. 1984. Morphological size and shape variation among local human populations. J Hum Evol 13:191-194.
30. Relethford JH. 2004. Boas and beyond: migration and craniometric variation. Am J Hum Biol 16:379-386.
31. Reyment RA. 1985. Multivariate morphometrics and analysis of shape. Math Geol 17:591-609.
32. Richtsmeier JT, Burke Deleon V, Lele SR. 2002. The promise of geometric morphometrics. Yearb Phys Anthropol 45:63-91.
33. Rohlf FJ. 1990. Rotational fit (Procrustes) methods. En: Rohlf FJ, Bookstein FL, editores. Proceedings Michigan morphometrics Workshop. Special publication n° 2. Museum of Zoology. Michigan: University of Michigan. p 227-236.
34. Rohlf FJ, Marcus LF. 1993. A revolution in morphometrics. Trends Ecol Evol 8:129-132. [ Links ]
35. Rohlf FJ, Slice DE. 1990. Extensions of the Procrustes Method for the optimal superimposition of landmarks. Syst Zool 39:40‑59. [ Links ]
36. Sibson R. 1978. Studies in the robustness of Multidimensional Scaling: Procrustes statistics. J R Statist Soc B 40:234-238. [ Links ]
37. Siegel AF. 1982. Robust regression using repeated medians. Biometrika 69(1):242-244. [ Links ]
38. Siegel AF, Benson RH. 1982. A robust comparison of biological shapes. Biometrics 38:341-350. [ Links ]
39. Slice DE. 1996. Three-dimensional generalized resistant fitting and the comparison of least-squares and resistant fit residuals. En: Marcus LF, Corti M, Loy A, Naylor G, Slice DE, editores. Advances in morphometrics. New York: Plenum Press. p 179-199.
40. Slice DE. 2001. Landmark coordinates aligned by Procrustes analysis do not lie in Kendall's shape space. Syst Biol 50:141-149. [ Links ]
41. Slice DE. 2005. Modern morphometrics. En: Slice DE, editor. Modern morphometrics in Physical Anthropology. New York: Kliwer Academic Publishers. p 1-45.
42. Slice DE. 2007. Geometric morphometrics. Annu Rev Anthropol 36:261-81. [ Links ]
43. Sneath PHA, Sokal RR. 1973. Numerical taxonomy. San Francisco: Freeman.
44. Thompson DW. 1917. On growth and form. London: Cambridge. Zelditch ML, Swiderski DL, Sheets HD, Fink WL. 2004. Geometric morphometric for biologists: a Primer. London: Academic Press.

