











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLa provincia de Santiago del Estero ocupa la parte más árida del sector septentrional de la llanura Chaco-Pampeana. Su clima es subtropical, con escasas lluvias estivales cuyas aguas se evaporan o filtran en charcos y pantanos ante la falta de declive. El territorio santiagueño es atravesado por dos grandes cursos de agua de régimen torrencial, los ríos Dulce y Salado, que nacen en los cordones montañosos de Salta y Catamarca y cruzan la provincia en sentido noroeste a sudeste. El río Dulce desemboca en la laguna Mar Chiquita, en la provincia de Córdoba, en tanto que el río Salado finaliza su curso en el río Paraná, en la provincia de Santa Fe (Fig. 1). Para las poblaciones humanas, la ocupación del terreno siempre ha estado determinada por la necesidad imprescindible de agua de alcance fácil y disponibilidad permanente. En Santiago del Estero, estas rutas obligatorias fueron trazadas por los cursos fluviales y depresiones, habilitadas especialmente durante la estación de lluvias. En este sentido, los ríos Dulce y Salado han constituido, desde tiempos inmemoriales, una importante ruta de comunicación entre las sociedades valliserranas del noroeste argentino y aquellas de las planicies hacia el sur y el oriente (Drube, 2009).

Fig. 1 Mapa de Santiago del Estero con la ubicación aproximada de los sitios arqueológicos de procedencia de las muestras estudiadasMapa de Santiago del Estero con la ubicación aproximada de los sitios arqueológicos de procedencia de las muestras estudiadas. El recuadro marca el área de los Bañados de Añatuya.
Las fechas de las primeras ocupaciones de esta región no son certeras, dada la esporádica presencia de sitios precerámicos con tecnología lítica, localizados mayormente en las serranías al sur del territorio provincial (Gómez, 1975; Gramajo de Martínez Moreno, 1992; Togo, 2004), para los cuales tampoco se cuenta con fechados radiocarbónicos. Para el siglo IV, la información se hace más palpable a partir de la identificación de los primeros asentamientos de grupos sedentarios dentro de la provincia, que se describen en mayor profundidad a continuación.
Históricamente, la Arqueología organizó la evidencia sobre los desarrollos culturales en el territorio santiagueño en una secuencia cronológica que reconocía dos grandes “Tradiciones”, subdivididas a su vez en “Fases”. Si bien esta aproximación al registro arqueológico resulta tipologista y homogeneizante, constituye a los fines de este trabajo una herramienta para contextualizar las muestras analizadas. La primera de estas Tradiciones arqueológicas, llamada “Cultura Las Mercedes” (Gómez, 1966), está integrada por un complejo agroalfarero proveniente del NOA, considerado el más antiguo del territorio santiagueño. Por los fechados que se disponen, Las Mercedes se habría desarrollado entre el 350 y el 1100/1200 d.C. (Del Papa, 2012). Hacia el 800 d.C., Las Mercedes se introduce a la llanura y toma contacto con grupos sedentarios portadores de alfarería de estilo Sunchituyoj, transmitiéndoles algunos de sus elementos de ideología Aguada. Es en este momento que se perfilan claramente los rasgos culturales característicos de la denominada “Tradición Cultural Chaco-Santiagueña” (Lorandi, 1978), ubicada en toda la cuenca de los ríos Dulce y Salado, la Mesopotamia entre ambas, y una considerable zona aún no definida al este y oeste de ambos ríos.
A pesar de la uniformidad reportada en la cultura material de las poblaciones santiagueñas tardías, se observa la coexistencia, entre los años 1200 y 1600 d.C., de dos principales tradiciones estilísticas según la decoración de la alfarería, las cuales podrían haber tenido orígenes geográficos diferentes (Lorandi, 1974). Una de ellas, llamada Sunchituyoj, es una expresión estilística de desarrollo local previo, con escasa dispersión fuera del territorio. Entre los años 1200 y 1400 d.C., aparecen en el registro evidencias de contacto constante entre los portadores de esta cerámica y los de la otra tradición característica de la Arqueología santiagueña, denominada Averías. La secuencia cronológica continúa con un momento posterior (entre 1420 y 1610 d.C.), marcado por la integración de estas dos grandes tradiciones alfareras a través del pasaje de rasgos de una a otra. Correlacionando estilos cerámicos y grupos étnicos, la hipótesis planteada por Lorandi (2015) es que dos grupos, al menos, pudieron compartir el mismo territorio y un gran número de asentamientos en Santiago del Estero, conformando aldeas biétnicas con alto grado de integración.
Hacia el final del período prehispánico, se observa la expansión de la cerámica Averías por fuera de Santiago, tanto en áreas aledañas (norte de Córdoba, este de Catamarca y Tucumán), como aún más lejos, hacia la zona de Andalgalá, Campo del Arenal, Valle del Cajón, Valle de Santa María y el norte de La Rioja, muy posiblemente debido al traslado de habitantes de la llanura santiagueña como mitimaes al servicio de los incas (Lorandi, 1980). Para esta última etapa, cercana a la llegada de los conquistadores, algunos autores han señalado la vinculación de la cerámica Averías con un aumento de la densidad poblacional y una intensificación de la producción agrícola y la actividad textil, en contraste con los registros que hay para asociaciones con otros tipos de cerámica (Taboada y Angiorama, 2010). En este contexto, los Bañados de Añatuya (Departamento Avellaneda) se configuraron como el sector de mayor desarrollo y relevancia del río Salado medio (Fig. 1).
Los grupos humanos asentados en los Bañados de Añatuya, asociados inicialmente al nombre de juríes y posteriormente denominados tonocotés por las crónicas coloniales, mantuvieron redes de relaciones con poblaciones de los valles intermontanos, los incas y los pueblos chaco-litoraleños al menos desde el año 1200 d.C. hasta tiempos coloniales (Taboada, 2011; Taboada y Angiorama, 2010; Taboada, Angiorama, Leiton y López Campeny, 2013). Estas relaciones se evidencian en el registro arqueológico por la presencia de objetos elaborados con materias primas alóctonas (metales, caracoles del Pacífico, tejidos), asignados al período prehispánico tardío e Inca, y que pueden asociarse con investiduras político-sociales andinas, interpretados como posibles dones y contradones en situación de alianza y reciprocidad (Taboada y Angiorama, 2010). Este tipo de bienes se ha encontrado en abundancia en el sitio Sequía Vieja (470±50 años C14 AP) y en menor medida y variedad en algunos de sus vecinos inmediatos, pero no en el resto de los cerca de 200 sitios conocidos para Santiago. Estas características hacen pensar en los Bañados de Añatuya como un núcleo sociocultural diferenciado de otros sitios más al norte o al este del Salado (Taboada, 2014). Lorandi (1980) plantea que, si bien el Tawantinsuyu no ocupó las tierras bajas propiamente dichas, muchos grupos de este origen, parcialmente transculturados a las pautas del altiplano y los valles colindantes, jugaron un rol importante en la defensa de la frontera oriental del imperio inca contra los ataques de los chiriguanos al norte y de los lules al sur. Estas áreas de frontera se habrían integrado al Tawantinsuyu a través de pactos políticos confirmados con alianzas matrimoniales y el intercambio de dones.
Con respecto a la relación entre los grupos sedentarios locales del Salado medio y los pueblos chaco-litoraleños, Lorandi (1978; 1980) y Castro Olañeta (2013) acuerdan en considerar que los lules, un grupo originario del Chaco, avanzaron hacia el oeste pocos años antes de la llegada de los españoles, mezclándose con los habitantes locales (tonocotés), sedentarizándose e incorporando nuevas prácticas de acceso y producción de recursos. Hay registros claros y presencia recurrente desde al menos el 1000 d.C. (Lorandi, 1978) de materiales y rasgos que se han asignado a tradiciones alfareras chaqueñas y del litoral, como las campanas y alfarerías gruesas y los apéndices en forma de loros en varios sectores de Santiago del Estero, y una importante cantidad de pipas cerámicas halladas en los sitios de Sequía Vieja y Averías en la zona de los Bañados de Añatuya. Estos elementos han sido interpretados como indicadores de una integración más bien pacífica y quizás bastante generalizada de los aportes poblacionales chaco-litoraleños (Taboada, 2017).
Estudios de deformación craneana en poblaciones agroalfareras de Santiago del Estero observaron la presencia minoritaria de individuos con un patrón deformatorio diferencial. Cráneos femeninos sin deformación, hallados en contextos con cerámica Averías, probablemente corresponderían a etnicidades chaqueñas, quienes no acostumbraban a realizar esa práctica. Por su parte, aquellos cráneos con deformación cuneiforme (del sitio Bocatoma) y tabular erecta planofrontal (provenientes de la región del río Salado medio) podrían constituir evidencias de contacto con poblaciones del área andina, en ese momento bajo dominio incaico, o bien con pueblos de las florestas de tierras bajas ubicadas al este de los Andes (Drube, 2010).
Teniendo en consideración el complejo escenario planteado por las diversas evidencias acerca de las interacciones entre las poblaciones santiagueñas y otros grupos prehispánicos de Argentina, es posible interrogarse si la composición genética de las poblaciones antiguas de Santiago del Estero refleja tales vínculos sociales y movimientos poblacionales con respecto a los habitantes del Noroeste argentino y a los grupos Chaco-Litoraleños. Por otra parte, resulta de interés científico indagar sobre la continuidad de los linajes maternos prehispánicos en la población santiagueña contemporánea, siendo esta última producto de los complejos y sucesivos eventos de mestizaje que caracterizaron la conformación de las poblaciones nacionales latinoamericanas. En ese sentido, el ADN antiguo (ADNa), es decir, aquel material genético que se extrae de restos arqueológicos, puede aportar información sobre los procesos evolutivos de las poblaciones humanas: permite abordar el estudio de eventos migratorios, patrones de residencia y relaciones genéticas entre poblaciones humanas, teniendo en cuenta su dimensión temporal (Hofreiter, Davis, Poinar, Kuch y Pääbo, 2001). Durante la última década se han incrementado los análisis de ADNa tanto en el actual territorio argentino como en el resto de Sudamérica (ver revisión en Llamas et al., 2016; Postillone, Crespo, Russo y Cardozo, 2014). La mayoría de los estudios genéticos de poblaciones antiguas utilizan marcadores uniparentales, principalmente del ADN mitocondrial (ADNmt) que, por su abundancia en la célula, incrementa las posibilidades de recuperar material genético de restos arqueológicos que han sufrido diferentes procesos que degradan el ADN (Hummel, 2003). Mediante técnicas moleculares se han identificado al menos 16 haplogrupos mitocondriales fundadores que ingresaron al continente americano en distintos momentos y utilizando rutas de poblamiento alternativas (Brandini et al., 2018; Malhi et al., 2010; Perego et al., 2010; Tamm et al., 2007). Dentro de estos haplogrupos, la definición de clados geográficamente restringidos permite inferir procesos de diferenciación local (Bodner et al., 2012; De Saint Pierre et al., 2012a y b; García, Pauro, Nores, Bravi y Demarchi, 2012; Gomez-Carballa et al., 2018; Motti et al., 2017).
En este trabajo se presentan los primeros resultados del secuenciamiento de la Región Hipervariable I del ADN mitocondrial de restos óseos humanos de la provincia de Santiago del Estero, con el objeto de contribuir al conocimiento sobre la diversidad genética de las poblaciones nativas de la llanura santiagueña y los procesos de interacción con grupos de otras regiones de Argentina en tiempos tardíos.
MATERIAL Y MÉTODOS
Muestra
Se seleccionaron 10 muestras dentarias procedentes de distintos sitios arqueológicos de la provincia de Santiago del Estero (Tabla 1, Fig. 1). Ocho de ellas fueron obtenidas de la colección de restos humanos del Museo de Ciencias Antropológicas y Naturales “Emilio y Duncan Wagner” de la ciudad de Santiago del Estero, entre las cuales 7 provienen de sitios ubicados en los Bañados de Añatuya, en tanto que una no posee datos de procedencia (pero puede presumirse un origen cercano al núcleo del Salado medio donde los hermanos Wagner realizaron sus excavaciones). El contexto arqueológico de los sitios de procedencia sugiere que estas muestras corresponden a la última fase de la secuencia cronológica de Santiago del Estero, entre los 1400 y 1600 d.C. (Fase Oloma Bajada-Icaño, sensuLorandi, 1974). Finalmente, 2 de las muestras seleccionadas fueron tomadas de la colección Von Hauenschild del Museo de Antropología de Córdoba (atribuibles a las excavaciones efectuadas por éste al norte del Río Dulce), una de ellas originaria del sitio Bocatoma y la otra sin datos de procedencia. No se han realizado dataciones en Bocatoma, no obstante, la cerámica en este sitio adscribe casi en su totalidad al complejo Sunchituyoj, con muy escasa presencia de tiestos Averías en superficie (Drube, 2010; Gómez, 2009).
TABLA 1 Muestras analizadas y resultados obtenidosn.d.:no determinado.
| Muestra | Procedencia | Polimorfismos | Linaje | ||
| RHV I (16023-16410) (+16000) | RHV II (128-256 y 368-585) | ||||
| Colección Wagner (región del Salado medio) | 9SE | Laguna Muyoj | 223T 298C 325C 327T | - | C1 |
| 11SE | Averías | n.d. | - | n.d. | |
| 12SE | Chilca Pozo | n.d. | - | n.d. | |
| 16SE | Laguna Muyoj | 223T 298C 325C 327T 344T | 146C 249d | C1b | |
| 489C 493G 523d 524d | |||||
| 22SE | Sequía Vieja | n.d. | - | n.d. | |
| 27SE | Tulip Loman | 223T 325C 327T | 489C 493G 523d 524d | C1b | |
| 34SE | Cañitas | n.d. | - | n.d. | |
| 46SE | Sin procedencia | 051G 223T 259T 271C 298C 311C 325C 327T | - | C1d1b | |
| Colección Von Hauenschild (Norte del Río Dulce) | 47SE | Sin procedencia | n.d. | - | n.d. |
| 54SE | Bocatoma | 223T 242T 311C 325C 362C | - | D1j | |
Análisis de ADN antiguo
Precauciones para evitar la contaminación
Diversas precauciones fueron tomadas para minimizar el riesgo de contaminación de las muestras a lo largo de todo el estudio, desde la recolección hasta el trabajo en el laboratorio de extracción y tipificación del ADN. En general, se siguieron las recomendaciones presentadas por Pääbo et al. (2004) y descritas por Nores y Demarchi (2011).
Extracción de ADN
El procesamiento de las piezas dentales y la extracción de ADN se realizaron por duplicado para cada muestra en el Laboratorio de ADN Antiguo, ubicado en el Centro de Investigación del Patrimonio Arqueológico de Córdoba (CIPAC). Se siguió un protocolo de extracción basado en la descalcificación y digestión de piezas dentarias. En primer término, las muestras fueron sumergidas en hipoclorito de sodio al 6% durante 5-15 minutos, luego lavadas repetidamente con agua destilada y secadas en estufa. A continuación, fueron pulverizadas con torno dental a baja velocidad. Aproximadamente 0,3g de polvo fueron incubados en presencia de 1ml de una solución de 0,5M EDTA (pH 8,0) y 1mg de proteinasa K a 56ºC con agitación durante toda una noche. Finalmente, se centrifugó a 5000g por 5 minutos y se purificó el sobrenadante obtenido usando el kit Wizard SV PCR Clean-Up System (Promega) que se basa en la afinidad del ADN por la sílice en presencia de altas concentraciones salinas. En cada extracción realizada se incorporó un blanco que consiste en un tubo sin muestra sometido al mismo protocolo, y que posteriormente es analizado en paralelo al ADN extraído, a fin de corroborar que no hubo contaminación durante el procedimiento.
Secuenciamiento de la Región Hipervariable I del ADN mitocondrial
Debido a la fragmentación del ADN en las muestras antiguas, se amplificaron por PCR tres fragmentos solapantes menores a 221 pares de bases (pb) de la Región Hipervariable I (RHV I), entre las posiciones 16023 y 16410, utilizando los iniciadores: HVR1_P1F y HVR1_P1Rc (Raff et al., 2010), M13-L16120, M13R-H16239 y M13-L16208 (Adachi et al., 2009), y R16410 (Gabriel et al., 2001) (Tabla 2). A fin de refinar la caracterización de los subhaplogrupos de C1, se secuenció un fragmento de 284 pb perteneciente a la Región Hipervariable II (RHV II), en busca del polimorfismo 493G, diagnóstico de C1b, utilizando los iniciadores F314 y R597 (Motti, 2012) (Tabla 2). Las reacciones de PCR se llevaron a cabo en un volumen total de 55μl conteniendo 1X de GoTaq Buffer (Promega), 0,20mM de cada dNTP (Promega), 0,28μM de cada iniciador, 2,75U de GoTaq DNA Polymerase (Promega), 0,1 mg/ml de BSA y 5-6μl de la muestra de ADNa. En las reacciones de amplificación se incluyeron controles positivos y negativos. Las condiciones de ciclado fueron: un ciclo inicial de desnaturalización durante 6 minutos a 94ºC; 50 ciclos de desnaturalización por 30 segundos a 94ºC, hibridación por 45 segundos a 53ºC y elongación por 45 segundos a 72ºC; y un ciclo de elongación final de 5 minutos a 72ºC. Los productos de PCR se enviaron a Macrogen (Seúl, Corea) para su purificación y secuenciamiento de ambas cadenas de ADN. Las secuencias obtenidas fueron corregidas manualmente y alineadas con la Secuencia de Referencia revisada de Cambridge (rCRS) (Andrews et al., 1999), usando el programa Sequencher versión 5.3 (Gene Codes Corporation). Posteriormente, fueron asignadas a los subhaplogrupos correspondientes utilizando el programa HaploGrep 2 (Kloss-Brandstaetter et al., 2011), que se basa en la base de datos Phylotree (van Oven, 2015), y mediante búsqueda bibliográfica.
TABLA 2 Información y referencia bibliográfica de los iniciadores utilizados
| Nombre | Posición | Secuencia | Largo del amplicón generado | Referencia bibliográfica |
| HVR1_P1F | 16023-16043 | 5' GTTCTTTCATGGGGAAGCAG 3' | 160 pb | Raff et al., 2010 |
| HVR1_P1Rc | 16182-16160 | 5' TTGATGTGGATTGGGTTTTT 3' | ||
| M13-L16120 | 16120-16139 | 5' TTACTGCCAGCCACCATGAA 3' | 118 pb | Adachi et al., 2009 |
| M13R-H16239 | 16220-16239 | 5' TGGCTTTGGAGTTGCAGTTG 3' | ||
| M13-L16208 | 16209-16227 | 5' CCCCATGCTTACAAGCAAG 3' | 165 pb | |
| R16410 | 16391-16410 | 5' GAGGATGGTGGTCAAGGGAC 3' | Gabriel et al., 2001 | |
| F314 | 314-333 | 5' CCGCTTCTGGCCACAGCACT 3' | 284 pb | Motti, 2012 |
| R597 | 576-597 | 5' GAGGAGGTAAGCTACATAAACT 3' |
Comparación de los haplotipos obtenidos
Se contrastaron los haplotipos determinados en la muestra prehispánica de Santiago del Estero con una base de datos de secuencias de la RHV I de poblaciones prehispánicas sudamericanas (Fehren-Schmitz et al., 2015; Figueiro et al., 2017; Gómez-Carballa et al., 2015; Llamas et al., 2016; Nores, Fabra, García y Demarchi, 2017; Postillone et al., 2017; Russo, Gheggi, Avena, Dejean y Cremonte, 2017 y referencias allí citadas), con el objeto de indagar sobre las interacciones entre las poblaciones santiagueñas y otros grupos prehispánicos. Además, se extendió la búsqueda a poblaciones contemporáneas del Cono Sur (Bobillo et al., 2010; Cabana, Merriwether, Hunley y Demarchi, 2006; Cardoso et al., 2013; Catelli et al., 2011; Figueiro, Hidalgo y Sans, 2011; García, Pauro, Bailliet, Bravi y Demarchi, 2018; Gómez-Carballa et al., 2018; Motti, 2012; Salas, Jaime, Álvarez-Iglesias y Carracedo, 2008; Sevini et al., 2013). Esta selección no pretende ser una síntesis completa de los estudios publicados de ADN mitocondrial en Sudamérica, sino servir como marco para estudiar a la población de Santiago del Estero en un ámbito geográfico mayor, incluyendo poblaciones que constituyan puntos de comparación coherentes con las hipótesis planteadas. Se construyó un mapa de isofrecuencias de la distribución actual de uno de los linajes de la muestra arqueológica discutidos en el texto, mediante el algoritmo de Kriging utilizado en el programa Surfer 10 (Golden Software 2002). Para ello se tuvieron en cuenta sólo los trabajos que presentaran frecuencias de la población total analizada.
Por otra parte, se compararon las secuencias obtenidas en este estudio con las reportadas para la población contemporánea de la provincia de Santiago del Estero (Catelli et al., 2011; Pauro, García, Nores y Demarchi, 2013) con el fin de rastrear la continuidad de los linajes mitocondriales prehispánicos en este territorio.
RESULTADOS
Las precauciones tomadas durante el procesamiento y análisis de las muestras en el laboratorio fueron exitosas, ya que no se observaron productos de amplificación en los controles negativos de PCR y blancos de extracción. Se obtuvieron secuencias confiables de la RHV I en cinco de las diez muestras procesadas, mientras que en las restantes no se obtuvieron amplificaciones o fueron espurias.
La región secuenciada (16023-16410 pb) permitió la identificación de haplotipos diferentes en las cinco muestras (Tabla 1). La muestra 54SE, proveniente del norte del Río Dulce, presenta las mutaciones 16242T y 16311C, características del subhaplogrupo D1j. Los demás individuos fueron asignados al haplogrupo C1. La presencia del polimorfismo 16051G en la secuencia del individuo 46SE indica su pertenencia al subhaplogrupo C1d, mientras que la mutación 493G observada en las secuencias de 16SE y 27SE permitió asignarlas a C1b. La primera de ellas exhibe, a su vez, una transición en la posición 16344; en tanto que la segunda, la reversión de la mutación 16298C, esperada para los linajes derivados de M8. Esta reversión se puede observar en paralelo en varias secuencias C1b, tanto antiguas como contemporáneas, con diversos orígenes geográficos y con polimorfismos diferentes en la RVH II (Bobillo et al., 2010; Kemp, Tung y Summar, 2009; Rieux et al., 2014; Tito, Polo y Lewis Jr, 2012). Por esta razón, la lectura realizada en el rango 368-585 de la RHV II no es lo suficientemente informativa como para inferir el vínculo filogenético entre la secuencia de 27SE y alguno de los individuos C1b+@16298 de la literatura.
Por último, la muestra 9SE sólo pudo ser secuenciada dentro del rango de lectura analizado para la RHV I, por lo que su tipificación quedó limitada al haplogrupo C1. Esto podría deberse al estado de conservación de la muestra, considerando que el fragmento de la RHV II seleccionado (284 pb) tiene un tamaño mayor al recomendado para análisis de ADN antiguo.
Al realizar una búsqueda en la literatura de secuencias que presentaran el haplotipo observado en el individuo 16SE, se observó que todas las muestras contemporáneas C1b+16344 de Argentina se agrupan en un único linaje, caracterizado a su vez por las mutaciones 16519C de la RHV I (aunque cae fuera del rango de lectura del presente trabajo) y 146C de la RHV II. El secuenciamiento de un fragmento de 128 pb entre las posiciones 128 y 256, utilizando los iniciadores M13-L127 y M13R-H257 (Adachi et al., 2009) permitió confirmar la pertenencia de la muestra 16SE a este linaje C1b+146.
Por otro lado, en la secuencia de 46SE se observa, además del polimorfismo 16051G, la coocurrencia de tres mutaciones en la RHV I (16259T 16271C 16311C), idénticas a las detectadas en un individuo kolla de Salta cuyo mitogenoma fue adscripto al clado C1d1b por Perego y colaboradores (2010). Por esta razón, si bien no se cuenta con la secuencia mitocondrial completa de 46SE, se asume su pertenencia a este linaje, definido por los polimorfismos 195C y 928G (Fig. 2).
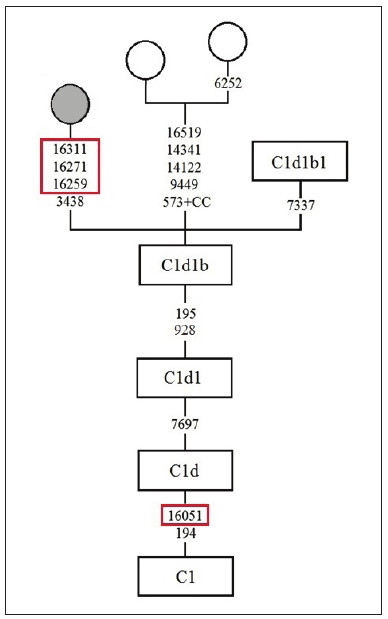
Fig. 2 Sección del árbol filogenético de C1Sección del árbol filogenético de C1, modificado de Perego et al. (2010). El círculo gris representa la muestra de un individuo kolla de Salta. Los recuadros rojos señalan las mutaciones identificadas en la secuencia del individuo 46SE de Santiago del Estero que permiten asumir su pertenencia al linaje C1d1b.
Con respecto a la comparación de los haplotipos obtenidos en la muestra de Santiago del Estero con las secuencias prehispánicas compiladas, se destaca que las únicas coincidencias encontradas, a excepción del haplotipo nodal C1 de 9SE, de amplia distribución en América, fueron con individuos de la región central de Argentina (provincia de Córdoba). El linaje D1j de 54SE fue observado también en dos individuos de 4525±20 (UCI AMS 22288) y 1192±40 (MTC 13247) años AP de las costas de la Laguna Mar Chiquita, provincia de Córdoba (García et al., 2012; Nores et al., 2017). Por su parte, el haplotipo C1d1b+16259 16271 16311 de 46SE sólo es compartido con la secuencia de un individuo de 2331±46 años C14 AP (AA93745) del sitio El Diquecito, ubicado en esa misma región. Este hecho resulta particularmente significativo, teniendo en cuenta que la base de datos de comparación está constituida por 496 secuencias antiguas, de las cuales sólo 31 pertenecen a restos arqueológicos de Córdoba. Por su parte, no se observaron los haplotipos de 16SE y 27SE en otras muestras arqueológicas.
DISCUSIÓN
La muestra arqueológica de Santiago del Estero analizada en este trabajo presenta varios linajes característicos de las poblaciones contemporáneas del centro de Argentina. De acuerdo con Motti (2012), la evidencia a nivel mitocondrial en poblaciones actuales permite postular la existencia de un núcleo de diferenciación poblacional en la región central de Argentina, desde los primeros momentos de ocupación, y un vínculo por origen común y/o extenso flujo génico entre grupos del centro (Córdoba y Santiago del Estero) y el oeste argentino (Catamarca, La Rioja y los valles cuyanos). El caso mejor caracterizado es el del subhaplogrupo D1j (Bodner et al., 2012), al cual pertenece la muestra 54SE del sitio Bocatoma, ubicado al norte del Río Dulce. Actualmente, la mayor incidencia de D1j se encuentra en la ciudad de La Rioja (45%), seguido de Santiago del Estero (28%), Chepes (27%), y las provincias de Catamarca (22%) y Córdoba (18%) (Motti, 2012; Pauro, 2015). Considerando su patrón de distribución geográfica, con mayor frecuencia y variabilidad en las Sierras Pampeanas, se ha postulado un origen local para este linaje, en el seno de una metapoblación de gran profundidad temporal, que habría ocupado esta región al menos desde hace 4525 años (García et al., 2012). Si bien también se han reportado secuencias D1j en individuos del Golfo San Matías, Río Negro, datados en 3100 y 2200-2300 años AP, se ha hipotetizado que este linaje podría haber arribado al norte de la Patagonia portado por grupos que siguieron una ruta interna a través del centro de Argentina (Crespo, Favier Dubois, Russo, Lanata y Dejean, 2017).
Otro de los linajes característicos del núcleo de diferenciación poblacional constituido por las regiones del centro y oeste de Argentina es C1d1b (Motti et al., 2017), al cual fue adscripto el haplotipo de la muestra 46SE de este estudio. Por su distribución geográfica y edad estimada, Perego y colaboradores (2010) asumen un origen sudamericano para este linaje, durante un estadio intermedio de la diferenciación in situ de los grupos nativo-americanos locales. En Argentina, C1d1b presenta una distribución continua con foco en la región central, alcanzando una frecuencia máxima del 19,5% en Santiago del Estero, seguido por los wichí de Formosa (9%), la localidad de Santa María en Catamarca (8%) y las provincias de Córdoba (7%) y San Juan (6,6%), para luego disminuir gradualmente su frecuencia hacia el norte y el sur (García et al., 2018; Pauro, 2015). Por su parte, el motivo derivado C1d1b+16259 16271 16311, observado en la muestra 46SE, ha sido reportado en poblaciones contemporáneas del NOA (Motti 2012; Perego et al., 2010), en Montevideo (Figueiro et al., 2011), en el Gran Chaco (Cabana et al., 2006; Sevini et al., 2013) y en el centro de Argentina (García et al., 2018). A partir de datos poblacionales, se construyó un mapa de isofrecuencias para representar la distribución actual de este linaje, indicando además la localización de los sitios arqueológicos donde fue identificado (Fig. 3). Su mayor frecuencia se encuentra actualmente en la región del Gran Chaco, alcanzando el 4,5% entre los wichí de Formosa y el 7% en la población de “criollos” de Misión Nueva Pompeya, provincia de Chaco, aunque está ausente entre los wichí de la misma localidad (Sevini et al., 2013). En el sur de Santiago del Estero, la frecuencia de este haplotipo es del 2,5% (Pauro et al., 2013) en tanto que, en la provincia de Córdoba, es del 1,2% (García et al., 2018).

Fig. 3 Mapa de isofrecuencias del linaje C1d1b+16259 16271 16311 en poblaciones contemporáneas del cono sur de SudaméricaMapa de isofrecuencias del linaje C1d1b+16259 16271 16311 en poblaciones contemporáneas del cono sur de Sudamérica -con excepción de las secuencias reportadas en Montevideo (Figueiro et al., 2011) y en Salta (Perego et al., 2010), por no presentar frecuencias poblacionales-. LQ: La Quiaca, Jujuy; TA: Tartagal, Salta; MA: Maimará, Jujuy; SM: Santa María, Catamarca; BE: Belén, Catamarca; SJ: Provincia de San Juan; VT: Villa Tulumaya, Mendoza; LR: Provincia de La Rioja (Motti, 2012); CBA: Provincia de Córdoba; SL: Provincia de San Luis (García et al., 2018); SGO: Sur de la provincia de Santiago del Estero (Pauro et al., 2013); FO: Wichí de Formosa (Cabana et al., 2006); MNP: Misión Nueva Pompeya, Chaco (Sevini et al., 2013); MI: Misiones; CO: Corrientes; BA: Buenos Aires; RN: Río Negro (Bobillo et al., 2010). Los rombos rojos indican la procedencia de los individuos arqueológicos que presentan este haplotipo.
Sin embargo, la antigüedad de la muestra arqueológica cordobesa portadora del motivo C1d1b+16259 16271 16311, datada en 2331±46 años C14 AP, puede interpretarse como evidencia de un origen local de este haplotipo derivado en el centro de Argentina. Bajo este escenario, la presencia de C1d1b+16259 16271 16311 observada actualmente en el Gran Chaco se debería a eventos migratorios post-contacto. Sevini y colaboradores (2013) realizaron un exhaustivo muestreo de la localidad chaqueña Misión Nueva Pompeya, observando evidencias de orígenes diferentes de los linajes mitocondriales nativos y de bajo flujo génico entre los habitantes que autoadscriben como wichí y los que se reconocen como “criollos”. Según estos autores, los criollos de Chaco descienden de familias provenientes de Salta y Santiago del Estero que, entre los siglos XIX y XX, se trasladaron al bosque chaqueño junto con su ganado. La alta frecuencia de este linaje observada en este grupo podría explicarse por efecto fundador o por acción de la deriva génica en una población de pequeño tamaño. Por otro lado, evidencias etnohistóricas extienden los movimientos poblacionales entre Santiago del Estero y el Gran Chaco hasta varios siglos hacia atrás, identificando el límite entre Salta y Formosa como un “refugio” para los indígenas tonocotés y lules que huyeron de la Gobernación del Tucumán ante la llegada de los primeros conquistadores, donde se habrían establecido y entablado buenas relaciones con grupos indígenas chaqueños (Farberman, 2016). Tales procesos de interacción en tiempos pericontacto darían cuenta de la presencia del linaje C1d1b+16259 16271 16311 entre los wichí de Formosa.
El hecho de que los linajes D1j y C1d1b+16259 16271 16311 sean compartidos por las poblaciones prehispánicas de Santiago del Estero y del norte de la provincia de Córdoba (Laguna Mar Chiquita), apoya la hipótesis de García y colaboradores (2012), basada en análisis de poblaciones contemporáneas, sobre una antigua metapoblación que habría habitado el área de las Sierras Pampeanas.
Por su parte, la muestra 16SE fue asignada al linaje C1b+146. De acuerdo con Motti (2012), a pesar de que la posición 146 es un sitio altamente variable, la distribución geográfica que presenta en el territorio argentino permite sustentar la hipótesis de la existencia de un linaje monofilético derivado de C1b, identificado en la Región Control únicamente por una transición en esta posición. Los mitogenomas completos disponibles para Argentina (Salta, Tucumán y Catamarca) pertenecientes al linaje C1b+146 (Tamm et al., 2007) presentan una transición en la posición 12813, lo cual refuerza la hipótesis de monofilia, que se extiende a las muestras que sólo han sido secuenciadas a nivel de la Región Control. Sin embargo, dado el nivel de resolución disponible para la mayoría de las muestras de Argentina, no es posible afirmar conexiones a nivel extra-regional (Motti, 2012).
A partir del análisis filogeográfico de C1b+146, Motti (2012) propone un modelo de amplia distribución inicial de un linaje ancestral que luego se habría diferenciado en forma local, con la aparición de linajes derivados que adquirieron mayor frecuencia en determinadas regiones. A pesar de que las mayores frecuencias se concentran en el centro-oeste del territorio argentino, existe gran variedad de haplotipos en la ciudad de Tartagal, Provincia de Salta. Dentro de la muestra poblacional de esa localidad, se reportó un haplotipo derivado de C1b+146 con la mutación 16344T, la cual está presente en una de las muestras de este trabajo. Al compilar las secuencias de Argentina con el haplotipo C1b+146 16344 y representar sus relaciones filogenéticas, se evidenció un patrón de diferenciación dentro de este sub-linaje que no estaba plasmado en la red filogenética elaborada por Motti (2012). En el esquema construido, se observa un estado ancestral de C1b+146 16519 194 16344, representado por una secuencia proveniente de la provincia de Córdoba, a partir del cual derivan dos haplotipos presentes en poblaciones del Gran Chaco, Tartagal y San Salvador de Jujuy (Fig. 4). Considerando el rango de lectura obtenido para la RHV II, no es posible definir la posición de la secuencia de 16SE en este esquema. Para ello, será necesaria la tipificación de los polimorfismos de la Región Control completa.

Fig. 4 Esquema filogenético de la diversidad del linaje C1b+146 16344 en ArgentinaEsquema filogenético de la diversidad del linaje C1b+146 16344 en Argentina. Los números representan la frecuencia absoluta de cada haplotipo en esa población. CBA: Provincia de Córdoba (García et al., 2018); TF: Toba de Formosa; PF: Pilagá de Formosa (Cabana et al., 2006); CC: Criollos de Chaco; WC: Wichí de Chaco (Sevini et al., 2013); TA: Tartagal, Salta (Motti, 2012); JU: San Salvador de Jujuy (Cardoso et al., 2013).
Retomando las hipótesis planteadas en la introducción con base en la evidencia arqueológica, los resultados producidos hasta el momento mediante el análisis de la Región Control del ADNmt de la muestra arqueológica de Santiago del Estero no permiten establecer fehacientemente una conexión entre estas poblaciones y aquellas que habitaron el NOA en tiempos tardíos e incaico, ni tampoco con los grupos chaco-litoraleños. En cambio, las secuencias mitocondriales prehispánicas obtenidas reflejan claramente un vínculo biológico entre las poblaciones de la llanura santiagueña y los grupos de la región central de Argentina (fundamentalmente de Córdoba) y de los valles del centro-oeste. A pesar de su reducido número, los haplotipos detectados dan cuenta de este fenómeno: de las cinco secuencias mitocondriales antiguas obtenidas, tres fueron asignadas a los linajes D1j, C1d1b y C1b+146 que han sido señalados como indicadores de un estrecho vínculo entre las poblaciones del centro y el oeste argentino, y del mantenimiento de flujo génico entre ellas (Motti, 2012; Motti et al., 2017).
Los resultados presentados en este trabajo se encuentran en línea con evidencias provenientes de otros abordajes de investigación bioantropológica. Estudios craneométricos realizados sobre la colección del Museo de Ciencias Antropológicas y Naturales “Emilio y Duncan Wagner” sugieren afinidad biológica entre las poblaciones tardías de Santiago del Estero y los grupos que habitaban el territorio de las actuales provincias de Córdoba, Catamarca, San Juan y Mendoza (Fabra y Demarchi, 2012). Por su parte, Cocilovo y Di Rienzo (1984-1985) encuentran mayor similitud entre las poblaciones de la Región Central (Córdoba y Santiago del Estero) y la de Belén (Catamarca), la cual, a su vez, se asemeja a las de San Juan y Mendoza.
Los estudios genéticos publicados hasta la fecha sobre poblaciones contemporáneas de Santiago del Estero tampoco hallaron evidencia de flujo génico significativo con grupos andinos, en tanto que reportaron varios linajes mitocondriales compartidos con poblaciones de las Sierras Pampeanas (en particular con Córdoba) y del Gran Chaco (Pauro et al., 2013). Teniendo en consideración estos antecedentes, resulta interesante destacar la continuidad de algunos linajes maternos observados en la muestra arqueológica aquí analizada. En la población de las localidades de Sumampa y Villa Atamisqui, ubicadas en el sur de la provincia de Santiago del Estero, los linajes D1j y C1d1b+16259 16271 16311 se encuentran presentes en una frecuencia del 13,8% y del 2,5%, respectivamente (Pauro et al., 2013). Sin embargo, no se ha detectado el haplotipo C1b+146 16344 de la muestra arqueológica 16SE en estas localidades. Esto podría deberse, bien a un error de muestreo, bien a la pérdida de linajes por procesos demográficos del pasado, como la contracción y/o el reemplazo poblacional.
Este trabajo constituye el primer análisis genético de individuos de sitios arqueológicos provenientes del actual territorio de la provincia de Santiago del Estero, lo cual le otorga especial relevancia a la hora de interpretar los patrones de poblamiento y movilidad en la región central de Argentina.
Aunque los resultados son preliminares, evidencian la importancia de los estudios paleogenéticos para contrastar hipótesis acerca de relaciones filogenéticas basadas en la distribución actual de linajes maternos, teniendo en cuenta que esta es producto de sucesivos eventos ocurridos en tiempos prehispánicos, así como durante los procesos de conquista y colonización española, y de construcción del estado-nación argentino. En ese sentido, creemos necesario continuar los análisis del ADNmt a un nivel resolutivo mayor, incrementando además el tamaño muestral y el número de sitios relevados.
Por otro lado, futuros estudios de ADNa en otras regiones de interés para entender los procesos sociales de las poblaciones prehispánicas tardías de las tierras bajas, tales como el Gran Chaco, podrían aportar valiosa información a la discusión acerca de la existencia de flujo génico con los grupos que habitaron el actual territorio de Santiago del Estero.

