










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkSerie correlación geológica
versión On-line ISSN 1666-9479
Ser. correl. geol. vol.25 no.1 San Miguel de Tucumán ene./jun. 2009
TEMAS DE PALEONTOLOGÍA I
Inner morphology of Palaeoanemone (Cnidaria: Actiniaria): a burrowing anemone of the Carboniferous of Argentina
Roberto Ricardo Lech1
1Centro Nacional Patagónico-CONICET, Brown 2915, (9120) Puerto Madryn, Chubut, Argentina. E-mail: rrlech@cenpat.edu.ar
Abstract: The rocks of marine carboniferous of Argentina present numerous associations of invertebrates, among which can be found burrowing anemones and also traces of its biological activity. Palaeoanemone is an exceptional case of fossilization of these organisms, preserving its morphology both external and internal. The anatomical knowledge that is known about the morphology of this genus is enlarged by observing details of the walls of their body, peristome and pharynx, as well as mesenteries and tentacles.
Key words: Cnidaria; Actiniaria; Inner Morphology; Carboniferous; Argentina.
Resumen: Morfología interna de Palaeoanemone (Cnidaria: Actinaria): anémona cavadora del Carbonífero de la Argentina. Las rocas del Carbonífero marino de Argentina presentes numerosas asociaciones de invertebrados entre los que se puede encontrar anémonas cavadoras como así también rastros de su actividad biológica. Palaeoanemone es un caso excepcional de fosilización de estos organismos en el que se preservo parte de su morfología tanto externa como interna. El conocimiento anatómico que se tiene de la morfología de este género es ampliado al describirse detalles de las paredes de su cuerpo, peristoma y faringe, así como mesenterio y tentáculos.
Palabras clave: Cnidaria; Actiniaria; Morfología Interna; Carbonífero; Argentina.
Introduction
A review of the actiniarian: Palaeoanemona marcusi, previously described by Lech (1986b) for carboniferous marine rocks of San Juan, Argentina, allowed for the recognition of part of the internal morphological characters, not observed before.
Palaeoanemone belongs to a rare case of fossilization of a solitary polyp of anthozoan cnidaria, in which it was possible to identify numerous anatomical features, as well as traces of its activity in the sediment.
The fossil specimens here studied are similar in its external and internal morphology to burrowing anemones. These belong to the actiniarians group, anthozoan cnidaria commonly called sea anemone. They are solitary organisms that lack hard parts, which present only the poly phase in their life cycle. They own a pedal disc with which they fix to hard objects, or their base is modified to drill the soft substrate of the sea floor. The tentacles can be organized in different rings around the mouth or they can round off the oral surface completely.
The body of the burrowing anemones can be divided in three sections (capitulum, scapus and physa). Internally, they show even mesenteries, either complete or incomplete, in hexamerous cycles and normally two siphonoglyphs.
Other anthozoans similar to the actiniarians are the ceriantharians. The latter, unlike the sea anemones, present various complete mesenteries and only one siphonoglyph; they have a stretched body and are adapted to living in tubes produced by them either on the sand or on the mud.
Throughout this work, the knowledge about this group of organisms is enlarged by identifying the internal morphological characteristics of the fossilized specimens found in the Leoncito Formation (late Lower Carboniferous early Upper Carboniferous) appearing on the southeast of the city of Barreal, San Juan province, Argentina (Figure 1.A).

Figure 1. A. Location of the outcrop of the Leoncito Formation with fossils, Leoncito Encima, San Juan, Argentina. B. Geological map of the Leoncito Formation in the hamlet of Leoncito Encima: 1. Recent sediments, 2. Paraconglomerate, 3. Orthoconglomerate, 4. Sandstone, 5. Intercalation of sandstone and lutyte, 6. Lutyte / Mudstone, 7. Outcrop with anemones fossils.
The paleontological and geological aspects of the Leoncito Formation have been studied mainly by Keidel and Harrington (1938), Baldis (1964), Lech (1986a-b, 1989), López Gamundi and Alonso (1982), and López Gamundi and Rosello (1995). An actualized condensed version about the bioestratigraphic aspects can be found in detail in Césari et al. (2007) and, Lech and Cisterna (2008).
Materials and Methods
Two fossilized specimens have been studied attributed to burrowing anemones, in which details of its external and internal morphology and its respective structures of bioturbation can be observed. These materials are stored at the collection of Invertebrate Paleontology of the Miguel Lillo Institute (PIL). The sample PIL: 13.283 would comprise the majority of the anemone's body, while the second sample (PIL: 12.946) is attributed to the anterior section of the organism.
The study of the internal morphology was done through two mirror-like polishes performed in both extremities of the sample PIL: 12.946, which were documented using the equipment macrophotography Nikon with KODALIT film (12 ASA speed), making positive copies on RC paper grade 4.
Systematic palaeontology
Starting from the morphological structures observed in the fossil specimens studied, we recognize the actinopharynx and the mesenteries, and these are two of the three diagnostic apomorphies of Anthozoa that include siphonoglyph presence (see Daly et al., 2007).
The internal and external morphological characteristics present in the body of the fossilized animal and its associated bioturbation structure (Figures 2.B, C: ec, ic and 5.A-C) suggest its relation with the group of actiniarians with tentacles that arise at the margin and/or from the disc, together with burrowing anemones.
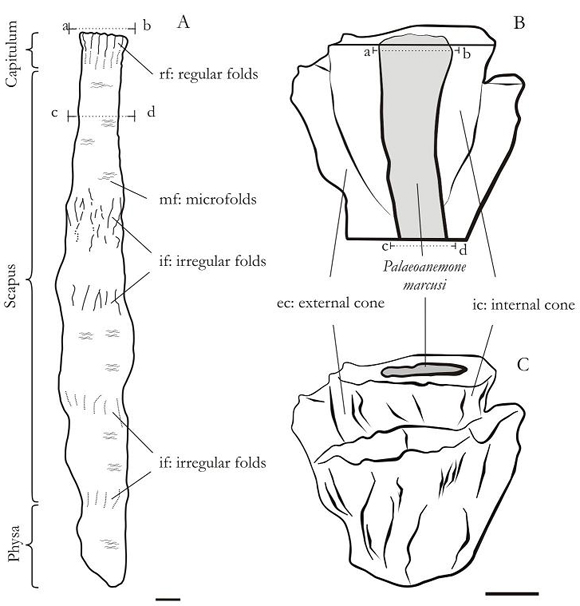
Figure 2. A. Reconstruction of Palaeoanemone marcusi based on PIL: 12.946 and PIL: 13.283. B. Transversal polished sections of P. marcusi (PIL: 12.946). B and C. Schematic of the structure of bioturbation track associated with the oral region of the Palaeoanemone marcusi (PIL: 12.946). B. Longitudinal section. a-b and c-d are the sections in which he notes the internal morphology described. Scale bar = 1 cm.
The systematic arrangement suggested by Daly et al., 2007 is adopted for the treated fossils specimens, although a lack of preservation of diagnostic characters cannot allow a taxonomic allocation to categories of lower hierarchy.
It dismisses allocation to ceriantarians because Palaeoanemone does not have the tube characteristic of this group, produced by discharge of specialized cnidae called ptychocysts and mucus that sheaths the column.
Phyllum Cnidaria
Class Anthozoa
Subclass Hexacorallia
Order Actiniaria
Family Incertae sedis
Forty five families were included in the Order Actiniaria by Daly et al. (2007), and only three of them would have burrowing anemone forms (Families: Andresiidae, Edwardsiidae and Iosactiidae). Unfortunately, in the studied fossil material, the morphological features that allow a specific diagnostic assignment to any of these families were not preserved.
Genus Palaeoanemone Lech, 1986b
Type species Palaeoanemone marcusi Lech.
Palaeoanemone marcusi Lech
Figures 2.A-C, 3.A-B, 4.A-B and 5.A-C
1986b. Palaeoanemone marcusi Lech: 186; Figures 6. A-D.
2008. Palaeoanemone marcusi Lech; Zamponi: 203, Figure 8.29.
Diagnosis (Lech, 1986b). Cylindrical body with a rounded aboral end and an oral region with 14 regular well delimited folds, which are wider rather than higher and separated by interfold spaces of same width as the folds, and of even bottom. The whole external surface of the body is covered by microfolds with an average density of 2,6 microfolds by millimetre.
Description extended. Cylindrical body with a rounded aboral end and a shortened oral end. The external surface of the oral region of the body presents 14 regular longitudinal folds (Figures 2.A: rf and 5.B), rounded, wider rather than higher with interfold spaces of same width than the folds, but of even background. A slight widening of the oral region can be observed owing to a greater development of the longitudinal folds. The rest of the body shows four narrowed areas originating irregular longitudinal folds, partly anastosomed (Figures 2.A: if and 5.C); those areas are expanded approximately at the same distance. The narrowed areas are highly marked towards the oral region producing a greater reduction in the diameter of the body in this region than in the aboral region. The entire outer surface is covered by microfolds that surround the body transversely with an average density of 2,6 microfolds by millimetre (Figures 2.A: mf and 5.B).
Diverse subcircular structures of different diameter and distribution have been observed by polishing the oral end. Three of those structures (Figures 3.A, 4.A: bw, sph, phw) are concentric to the longitudinal axis of the body and delimit three areas. The peripherical area, delimited by the bw and sph structures, presents a series of very small subcircular structures (Figures 3.A, 4.A: t) with a diameter that varies between 1 and 2,5 millimetres; some of them coincide with external longitudinal folds. The sph structure is very thin and sinusoidal. In the second area, delimited by the intermediate and central subcylindrical structure (Figures 3.A, 4.A: sph, phw), six or seven septal structures are observed (Figures 3.A, 4.A: m), delimiting other subcircular areas (Figures 3.A, 4.A: cc). The central area delimited by phw presents a homogeneous surface (Figures 3.A, 4.A: ph).
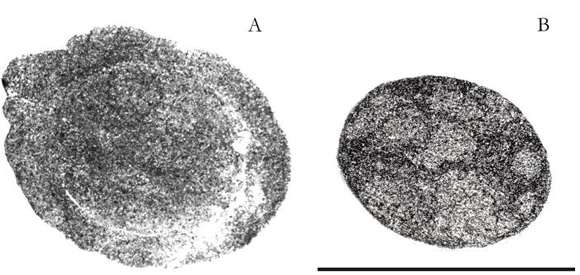
Figure 3. A,B. Transversal polished sections of Palaeoanemone marcusi (PIL: 12.946). A. Oral section, B. Distant section to 1,6 cm of the previous polished one. Scale bar = 1 cm.
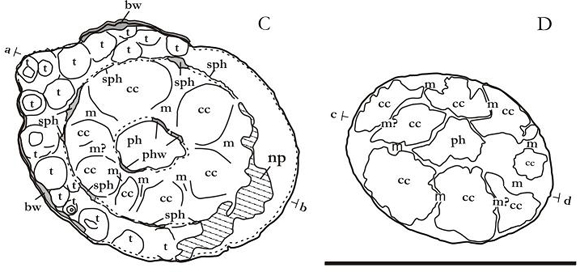
Figure 4. A,B. Interpretation of the internal morphology. cc: corporal cavity, ph: pharynx, np: not preserved, bw: body wall, phw: pharynx wall, m: mesentery, m?: dubious mesentery, sph: sphincter, t: tentacle. a-b and c-d are the sections in which he notes the internal morphology described and shown in Figures 2.A.B. Scale bar = 1 cm.
Subcircular structures, some with irregular contour and diverse diameter, have been observed in the second polished distant from the previous 16 millimetres (Figures 3.B, 4.B: cc, ph). These structures are distributed more or less homogeneously and they are not in contact with each other (Figures 3.B, 4.B: m).
Stratigraphic position and age. Leoncito Formation (Figures 1.B), stratigraphic level of pelite with anemones (31°47´49´´S, 69°20´W); from to Late Early Carboniferous.
Comparisons. The fossil record of sea anemones is restricted to the Palaeozoic and most of the known species are attributed to actiniarians. However, traces of its biological activity can be found in the rocks dating from several geological ages.
Mackenzia costalis Walcott of the Middle Cambrian of Burgess Shale, Canada, is characterized by having an elongated body with prominent longitudinal folds which end in a disc-shape like structure (Conway Morris, 1993). Because of its external morphological characteristics, it is believed that these folds belonged to the internal partitions and that it owned a large gastrovascular cavity (Briggs et al., 1994), but as no tentacles were observed, its inclusion in the group of sea anemones would be problematic (Hou et al., 2005).
Another taxon of doubtful relationship to sea anemones is Xianguangia sinica Chen and Erdtmann of the Lower Cambrian of China. It has a cylindrical spindle-shape with 16 elongated tentacles, each bearing numerous fine arms or branches with setae (Chen and Erdtmann, 1991; Hou et al., 2004). This fossil is considered both as a ctenophore and/or as an actiniarian (Hou et al., 1999), including doubts about its allocation within sea anemones (Hou et al., 2005).
Archisaccophyllia kunmingensis Hou et al. (2005) of the Cambrian of China presents an elongated body, distinguished by a pedal disc, a column and an oral disc, probably with 12 simple tentacles.
The scarce paleontological register is probably owed to the characteristic organic nature of these organisms that lack hard parts. However, there are numerous tracks attributed to burrowing anemones throughout geological times, like the Conostichus Lesquereux (1876) and Bergaueria Prantl (1946). From an interpretative approach, Conostichus is considered as rests of an organism related to burrowing anemones, while Bergaueria could be considered as a casing? (refuge?) structure or as rest of themselves (Pemberton et al., 1988).
Discussion and conclusions
The longitudinal folds (Figures 2.A: rf and 5.B) as well as the small subcircular structures (Figures 3.A, 4.A: t) in the oral region would give evidence of the longitudinal epidermic muscles of the tentacles that the present burrowing anemones possess. The peripherical subcircular structures mentioned (Figures 3.A, 4.A: t) in the oral region would correspond to tentacles that arise at the margin and/or from the oral disc.

Figure 5. A. Structure of bioturbation track. B. Oral region of Palaeoanemone marcusi (Paratipo PIL: 12.946) and C. Body and structure of bioturbation track. (Holotipo PIL: 13.283). Scale bar = 1 cm.
The external subcircular structure (Figures 3.A, 4.A: bw) could be interpreted as the body wall, while the central subcircular structure (Figures 3.A, 4.A: wph) would belong to the pharynx wall.
The intermediate circular structure (Figures 3.A, 4.A: sph) which is presented as an internal limit to the tentacles of oral disk, could be interpreted as remnants of the circular muscle that correspond to the marginal sphincter musculature in the current anemones.
The septal and subcircular structures (Figures 3.A.B, 4.A.B: m, cc) that surround the pharynx would be the equivalent to some of the different types of mesenteries and to the body cavities present in the anemones (Hyman, 1940; Brusca and Brusca, 1990).
The presence of complete mesenteries helps in the function of the internal hydraulic skeleton by limiting the maximum expansion of the diameter of the column when the muscles retract. The transversal microfolds (Figures 2.A: mf and 5.B) could belong to vestiges on the body of the peristaltic movements of the body's animal, occurred during the landslip of the sediment. The movement or progression through general contractions of the body is given by the presence of a hydrostatic skeleton that allows the progressive movement of that body as a whole, through changes in its own shape. That hydrostate or hydrostatic skeleton is the combination of the structure and functional character of the organism, which allow the use of water that some division of its body possesses as a support and the transmition of muscular force.
The hydrostatic skeleton are of constant volume and any local increase in pressure, caused by muscular contraction, will be transmitted equally through the hydrostatic skeleton.
Sea anemones are animals that have cylindrical bodies and the body's wall is frequently strengthened by unflexible fibre nets. These fibrous nets harden the body's wall, avoiding aneurism when the hydrostate is pressed and allowing changes in length and diameter on the body. The circulation of water in the internal gastrovascular cavity generates a flow that helps to maintain the hydrostate, which at the same time allows muscular contraction (Ruppert and Barnes, 1996).
When water circulates in an area, it generates a force that allows a specific job in another place. The displaced fluids allow parts of the body or tentacles to enlarge and extend in order to anchor in holes or sediments by introducing part of its body, and allowing the production of peristaltic waves all through the body surface to build galleries or tubes.
The simplest means of movement is stretching and retracting, which usually implies alternative changes in the body's diameter, which allows the movement on or in the substrate. Such movement is witnessed by the structure of bioturbation associated with Palaeoanemone (Lech, 1986b).
Acknowledgements
I am greatly indebted to Dr. Néstor Ciocco from the IADIZA-CCT-CONICET, Mendoza for their constructive comments to the manuscript, as well as with to Lic. Alicia Marcus for his invaluable assistance in the development of this work. A special thanks to Mr Franco Pertini from the Photographic Laboratory of Centro Nacional Patagónico for their dedication and pacience in the preparation of the photographs that illustrate the internal morphology of the fossils specimens.
References
1. Baldis, B. 1964. Estratigrafía y estructura del Paleozoico al Sud del Arroyo de las Cabeceras, Ea. Leoncito (San Juan). Boletín de Informaciones Petroleras 365: 28-33. [ Links ]
2. Briggs, D.E.G., Erwin, D.H. and Collier, F.J. 1994. The fossils of the Burgess Shale. Smithsonian Institutions Press, Washington DC, 238 pp. [ Links ]
3. Brusca, R.C. and Brusca, G.J. 1990. The Invertebrates. Sinauer Associates, Sunderland, MA (USA), 922 pp. [ Links ]
4. Césari, S.N., Gutiérrez, P.R., Sabattini, N., Archangelsky, A., Azcuy C.L., Carrizo, H.A., Cisterna, G., Crisafulli, A., Cúneo, R.N., Díaz Saravia, P., di Pasquo, M., González, C.R., Lech, R.R., Pagani, M.A., Sterren, A., Taboada, A.C. and Vergel, M.M. 2007. Paleozoico Superior de Argentina: un registro fosilífero integral en el Gondwana occidental. Asociación Paleontológica Argentina, Publicación Especial 11, Ameghiniana 50o. Aniversario: 35-54. [ Links ]
5. Chen, J.-Y. and Erdtmann, B.D. 1991. Lower Cambrian Lagerstätte from Chengjiang, Yunnan, China: insights for reconstructing early metazoan life. In: Simonetta, A.M. and Conway Morris, S. (eds.), The early evolution of Metazoa and the significance of problematic taxa. Cambridge University Press, Cambridge, pp. 57-76. [ Links ]
6. Conway Morris, S. 1993. The fossil record and early evolution of the Metazoa. Nature 361: 219-225. [ Links ]
7. Daly, M., Brugler, M.R., Cartwright, P., Collins, A.G., Dawson, M.N., Fautin, D.G., Scott, C.F., Mc Fadden, C.S., Opresko, D.M., Rodriguez, E., Romano, S.L. and Stake, J.L. 2007. The phylum Cnidaria: A review of phylogenetic patterns and diversity 300 years after Linnaeus. In: Zhang, Z.-Q. and Shear, W.A. (eds.), Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa 1668: 127-182. [ Links ]
8. Hou, X.-G., Bergström, J., Wang, H.-F., Fang, X.-H. and Chen, A.-L. 1999. Fauna. Exceptionally Well-Preserved Animals from 530 Millions Years Ago. Yunnan Science and Technology Press. Kunming, 170 pp. [ Links ]
9. Hou, X.-G., Aldridge, R.J., Bergström, J., Siveter, D.J., Siveter, D.J. and Feng, X.-H. 2004. The Cambrian Fossils of Chengjiang, China: the flowering of animal life. Blackwell Publishing, Malden. Massachusetts, 233 pp. [ Links ]
10. Hou, X.-G., Stanley, G.D., Jie Zhao, JR. and Ma, A.-Y. 2005. Cambrian anemones with preserved soft tissue from the Chengjiang biota, China. Lethaia 38: 193-203. [ Links ]
11. Hyman, L. H. 1940. The Invertebrates. Volume 1, Protozoa through Ctenophora. McGraw-Hill Book Company, Inc. New York and London, 726 pp. [ Links ]
12. Keidel, J. and Harrington, H.J. 1938. On discovery of Lower Carboniferous tillites in the Precordillera of San Juan, Western Argentina. Geological Magazine 75: 885, 103-129. [ Links ]
13. Lech, R.R. 1986a. Estratigrafía del Yacimiento fosilífero de la Formación Leoncito (Carbónico), Depto. Calingasta, San Juan. Primeras Jornadas de Geología de Precordillera (San Juan, 1985). Asociación Geológica Argentina, Serie "A" Monografías y Reuniones N° 2: 163-167. [ Links ]
14. Lech, R.R. 1986b. Anémonas cavadoras fósiles y su rastra de bioturbación en el Carbonífero inferior marino de San Juan, Argentina. Ameghiniana 23: 185-190. [ Links ]
15. Lech, R.R. 1989. Algunos braquiópodos de la Formación Leoncito, Carbónico inferior de la provincia de San Juan, Argentina. 4o. Congreso Argentino de Paleontología y Bioestratigrafía (Mendoza, 1986), 4: 5-10. [ Links ]
16. Lech, R.R. and Cisterna, G. 2008. Leoncito (Formación...). In: Gutiérrez P.R. (ed.), Léxico Estratigráfico de la Argentina, Vol. VI, Carbonífero. Asociación Geológica Argentina, Serie "B" (Didáctica y Complementaría) N° 30; Servicio Geológico Minero Argentino, Publicación N° 169, pp. 106-107. [ Links ]
17. Lesquereux, L. 1876. Species of fossils marine plants from the Carboniferous measures. Indiana Geological Surey,Annual Report 7: 134-145. [ Links ]
18. López Gamundí, O. and Alonso, M.S. 1982. Áreas de Proveniencia de las Sedimentitas Carbónicas (F. Majaditas, F. Leoncito y F. Hoyada Verde) Barreal, San Juan, Argentina. 5o. Congreso Latinoamericano de Geología (Buenos Aries) 2: 481-490. [ Links ]
19. López-Gamundí, O.R. and Rossello, E.A. 1995. Pavimento glacial en la Formación Leoncito (Carbonífero), Precordillera Occidental, San Juan. Revista Asociación Geológica Argentina 50: 35-39. [ Links ]
20. Pemberton, S.G., Frey, R.W. and Bromley, R.G. 1988. The ichnotaxonomy of Conostichus and other plug-shaped ichnofossils. Canadian Journal Earth Sciences 25: 866-892. [ Links ]
21. Prantl, F. 1946. Two new problematic trails from the Ordovician of Bohemia. Académie Tchèque des Sciences, Bulletin International, Classe de Sciences Mathématiques, Naturelles et de la Médecine 46: 49-59. [ Links ]
22. Ruppert, E.E. and R.D. Barnes. 1996. Zoología de los Invertebrados. McGraw-Hill Interamericana Editores. Quinta edición. 1114 pp. [ Links ]
23. Zamponi, M.O., 2008. Cnidaria. In: Camacho, H.H. and Longobucco, M.I. (eds.) Los Invertebrados Fósiles. Fundación Historia Natural Félix de Azara, Universidad Maimónides: 171-219. [ Links ]
Recibido: 16 de Septiembre de 2009
Aceptado: 02 de Diciembre de 2009

