










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkSerie correlación geológica
versión On-line ISSN 1666-9479
Ser. correl. geol. vol.25 no.1 San Miguel de Tucumán ene./jun. 2009
TEMAS DE PALEONTOLOGÍA I
Microfloras ordovícicas en Sierra de Zenta, Cordillera Oriental Argentina
Lucía Aráoz1
1Miguel Lillo 205, INSUGEO, Facultad de Ciencias Naturales e I.M.L., U.N.T. CONICET. E-mail: lucia_araoz@yahoo.com.ar
Resumen: En este trabajo se presenta un estudio integrado de las microfloras ordovícicas registradas en la Sierra de Zenta, NW Argentina (provincias de Jujuy y Salta), realizado a partir del análisis sistemático de palinomorfos, principalmente acritarcos y con el aporte de otros grupos como algas prasinofíceas, quitinozoos y criptosporas. La Sierra de Zenta está localizada en una zona transicional entre la Cordillera Oriental y las Sierras Subandinas, mostrando características comunes con ambas provincias geológicas. El sector norte de la sierra presenta características similares a la Cordillera Oriental, donde se identificaron las formaciones Santa Rosita y Acoite (Grupo Santa Victoria). Mientras que el sector centro y sur de la sierra se observó similitudes con las Sierras Subandinas, aunque las unidades se identificaron informalmente como Unidades Formacionales. Se describen dos perfiles geológicos, uno continuo en el sector norte, en Abra de Zenta Abra Límite, y otro representado por un perfil integrado, descripto mediante cuatro perfiles parciales y relevado en el sector central de la sierra, en Abra Buey Ojo Laguna Verde. En base al registro de acritarcos se definieron cinco asociaciones palinológicas informales (ASZ) cuyas edades fueron asignadas según la distribución temporal de los taxones componentes. Estas asociaciones fueron comparadas con las de otras secuencias ordovícicas de Argentina y diversas regiones del mundo, estableciendo correlaciones parciales bioestratigráficas y paleogeográficas.
Palabras clave: Acritarcos; Ordovícico; Bioestratigrafía; Zenta; Argentina.
Abstract: Ordovician microfloras of the Sierra de Zenta, Cordillera Oriental Argentina. This work presents the first integrated research on the Ordovician microfloras recorded in the Sierra de Zenta, NW Argentina (Jujuy and Salta provinces), and was carried out on the basis of systematic analysis of palynomorphs, mainly acritarchs. The Sierra de Zenta is located on a transitional zone between the Cordillera Oriental and the Sierras Subandinas, sharing common characters to with both geological provinces. The northern sector of the Sierra displays similar characters to the Cordillera Oriental, with the presence of the Santa Rosita and Acoite formations (Santa Victoria Group). Although the central and southern sector of the Sierra de Zenta presents a great resemblance to the Sierras Subandinas, there is not a perfect match on the stratigrahy of their Ordovician units. Two geological profiles are described, a continuous one in the northern sector of the Abra de Zenta Abra Límite, and another that is represented by an integrated section relevated in the central part of the sierra, between the Abra Buey Ojo Laguna Verde. The last stratigraphical sector is composed by four partial sections. In the palynological samples the acritarchs are dominant elements, while prasinophyte, chitinozoans and cryptospores represent minor escorting microfossils. Based on acritarchs records, five informal associations were defined whose ages were assigned according to the chronostratigraphical distribution of the taxones components (ASZ). These associations are compared to other Ordovician sequences of Argentina and diverse regions of the world, establishing some partial bioestratigraphical and paleogeographical correlations.
Key words: Acritarchs; Ordovician; Biostratigraphy; Zenta; Argentina.
Introducción
En el contexto de la biodiversificacion del Paleozoico Inferior el fitoplancton marino estuvo mayormente representado por acritarcos. Estos microfósiles de paredes orgánicas, heterogeneidad morfológica y relativamente fácil extracción, son una herramienta de gran utilidad en los estudios bioestratigráficos del Paleozoico Inferior a nivel mundial con un gran potencial para las reconstrucciones paleoecológicas y paleogeográficas.
Este trabajo constituye un resumen de la Tesis Doctoral "Estudios microflorísticos de la Sierra de Zenta, Cordillera Oriental Argentina" (Aráoz, 2009) y presenta un estudio integrado de las asociaciones microflorísticas provenientes de niveles ordovícicos de la Sierra de Zenta, realizado a partir del análisis sistemático de acritarcos que ocurren asociados a otros palinomorfos como quitinozoos, prasinofíceas y criptosporas.
La Sierra de Zenta es un importante núcleo orográfico del Noroeste Argentino, situado entre las provincias de Salta y Jujuy (Departamentos Iruya y San Ramón de la Nueva Orán) y Jujuy (Departamento de Humahuaca) en una zona geológica intermedia entre la Cordillera Oriental y las Sierras Subandinas (Figura N° 1). Se trata de una extensa región para la que se tiene un conocimiento geológico relativamente limitado, sobre todo en cuanto a su paleontología y microfloras, donde se destacan los extensos afloramientos de niveles ordovícicos que permitieron obtener importantes material para estudios palinológicos.
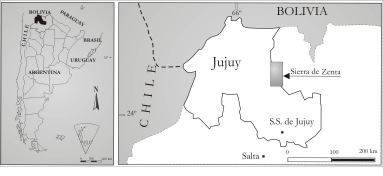
Figura 1. Mapa de ubicación de la Sierra de Zenta en el Noroeste Argentino, provincias de Salta y Jujuy.
Las microfloras analizadas fueron comparadas con las asociaciones descriptas para otras secuencias ordovícicas de Argentina y de diversas regiones del mundo, estableciendo relaciones paleogeográficas y bioestratigráficas. A partir de estos datos palinológicos se infieren condiciones paleoecológicas determinantes en la distribución de estos organismos en el medio, de modo que se logró un mejor entendimiento de las distribuciones microflorísticas en la región durante el Ordovícico.
Metodología
El trabajo se dividió en varias etapas que incluyeron recopilación de antecedentes bibliográficos y cartográficos, levantamiento de perfiles y recolección de muestras para análisis palinológicos, procesamiento y montaje del material palinológico, identificación y descripción de los palinomorfos, análisis de sus distribuciones espaciales y temporales, interpretación bioestratigráfica, paleoecológica y paleogeográfica.
En la revisión de la cartografía se observaron diferencias en las asignaciones de edades de algunos niveles aflorantes (Hoja Geológica 2366 IV, Ciudad de Libertador General San Martín de González et al., 2004; Cuenca del Noroeste, Mapa de Y.P.F.; Geología de la Región Noroeste, R.A. - provincias de Salta y Jujuy, Dirección de Fabricaciones Militares; Mapa Geológico de la Provincia de Salfity y Monaldi, 1998), en base a lo cual se consideró que la Hoja Geológica 2366 IV es el más adecuado a las observaciones de campo.
La sucesión fue relevada y descripta en secciones separadas como perfiles parciales y posteriormente fueron vinculados en un perfil integrado para el Ordovícico de la sierra. Se diferenciaron dos perfiles, uno en el sector norte de la sierra, entre Abra de Zenta y Abra Límite y el otro en el sector centro-sur de la sierra entre Abra de Buey Ojo y Laguna Verde.
De los niveles ordovícicos se extrajo un total de 68 muestras de las cuales 38 resultaron fértiles y 30 estériles, procesadas con técnicas estándar de ácidos fluorhídrico y clorhídrico, posteriormente montadas en gelatina-glicerina en el laboratorio micropaleonto-lógico del Instituto Superior de Correlación Geológica (INSUGEO-CONICET /UNT). Los preparados palinológicos fueron incorporados a la Colección Palinológica del Instituto Miguel Lillo (LIL PB pm 425, 427, 428, 429, 432, 433 y 444) y a la Colección de Paleopalinología del INSUGEO-CONICET/UNT (LIL PB pm / ICG 100-130). Se usaron Microscopio Optico de luz transmitida Carl Zeiss Jena Laboval 4, con aumentos 10x, 40x y 100x de la Cátedra de Paleontología de la Facultad de Ciencias Naturales e I.M.L. (UNT) y Fotomicroscopio Trinocular Nikon Eclipse 80i del Instituto Superior de Correlación Geológica con cámara digital Nikon Coolpix S10 (INSUGEO-CONICET / UNT).
Entre los palinomorfos se identificaron principalmente acritarcos con una menor proporción de algas prasinofíceas, quitinozoos y criptosporas asociadas. La identificación sistemática de los acritarcos se basó en sus caracteres morfológicos con un ordenamiento en una clasificación alfabética de géneros y especies (Tongiorgi et al., 2003). Se identificaron 32 géneros de acritarcos con 53 especies y otras formas que permancen indeterminadas hasta el momento; también 3 géneros y 1 especie de prasinofíceas; 4 géneros de quitinozoos y criptosporas indeterminadas. Se elaboraron cuadros de distribución temporal de especies seleccionadas en base a lo cual se diferenciaron cinco asociaciones palinológicas que fueron comparadas con las de otras localidades del Noroeste Argentino y del mundo para establecer correlaciones de edad.
La nomenclatura usada para la unidades cronoestratigráficas ordovícicas son las tradicionales series como también las nuevas unidades definidas por la International Commission on Stratigraphy, según corresponda su uso en los trabajos citados. Para asignar edades en las unidades formacionales y asociaciones palinológicas descriptas en este trabajo se usa la nomenclatura de Webby et al. (2004).
Antecedentes generales para la Sierra de Zenta
La geología del Noroeste Argentino es ampliamente conocida en sus diferentes aspectos: estructurales, estratigráficos, paleontológicos y bioestratigráficos entre otros. En contraposición, el conocimiento geológico de la Sierra de Zenta es parcial, de tipo regional y vinculado principalmente a la confección de mapas geológicos regionales (Turner, 1960; 1964; Amengual y Zanettini, 1973; Zanettini, 1973; González et al., 2004) y a las actividades mineras (Castillo et al., 1996; Castillo, 1999; Sureda, 1999; Boso y Monaldi, 2008) y otros trabajos más específicos como Nieva y Aceñolaza (2006) y Aceñolaza et al. (2008). Localidades fosilíferas del Ordovícico de la Sierra de Zenta son mencionadas en Aceñolaza y Aceñolaza (2002); Aceñolaza y Gutiérrez-Marco (2002); Aceñolaza y Milana (2005); Aceñolaza et al. (2006); Benedetto et al. (2007) y Tortello et al. (2008).
Los únicos trabajos estratigráficos de detalle para la sierra son de Starck (1995; 1996; 2008) y Starck et al. (1992 b; 1993 a, b) donde se analizan las secuencias de los ciclos sedimentarios siluro - devónicos y carbonífero - pérmicos aflorantes en diferentes sectores de la sierra, entre ellos el Abra de Zenta, cuya edad es reconsiderada en este trabajo a partir de nuevos datos palinológicos.
En los últimos años ha sido notable el avance en los estudios geológicos del Ordovícico de la sierra, especialmente en lo que se refiere a paleontología y bioestratigrafía, entre ellos Aceñolaza et al. (2001) y Aceñolaza et al. (2003) analizan en el Abra de Zenta un perfil estratigráfico que asignan a la Formación Santa Rosita (Grupo Santa Victoria), con una edad Tremadociano inferior alto en base a la presencia de trilobites de la Biozona de Kainella meridionalis (descriptos posteriormente por Tortello y Aceñolaza, 2008), dato confirmado por los palinomorfos asociados. Los trabajos referidos a los conodontes presentes en la sierra son de Heredia y Aceñolaza (2005); Albanesi et al. (2007 a); Ortega et al. (2007); Aceñolaza y Heredia (2008); Heredia et al. (2008) y Albanesi et al. (2008). Otros trabajos paleontológicos y bioestratigráficos son de Aceñolaza y Aceñolaza (2007) y Aráoz et al. (2008).
Los antecedentes palinológicos para la Sierra de Zenta son escasos, entre ellos Aráoz y Vergel (2001) describen la asociación microflorística del Abra de Zenta asociada a los trilobites de la Biozona de Kainella meridionalis que fue analizada en mayor profundidad por Aráoz (2002; 2009) y Aceñolaza et al. (2003). Otros trabajos son los de Aráoz y Aceñolaza (2004) que dan a conocer una microflora asignada al Tremadociano Tardío provenientes de niveles asignados a la parte inferior y media del Grupo Santa Victoria y Aráoz (2008) y Aráoz et al. (2008) que resumen los datos paleontológicos/palinológicos/bioestratigráficos del Paleozoico Inferior de la Sierra de Zenta, mientras que Rubinstein et al. (2008) analizan el límite Ordovícico - Silúrico en la sección de Caspalá.
Otros datos palinológicos de niveles paleozoicos son del Devónico, Carbonífero y Pérmico del norte de la sierra, en la localidad de Abra Límite, con trabajos de di Pasquo y Vergel (2008); Noetinger y Di Pasquo (2008) y Vergel et al. (2008 a, b).
Antecedentes palinológicos para el Noroeste Argentino
En cuanto al estudio de las microfloras eopaleozoicas del Noroeste Argentino, se destaca el notable incremento que tuvo en los últimos años, aunque todavía es necesaria mucha información de localidades clave y de áreas todavía inexploradas. Algunos trabajos palinológicos de localidades ordovícicas del Noroeste Argentino que fueron usados como referencia en el análisis de las microfloras de la Sierra de Zenta son: Volkheimer et al. (1980); Martin (1982); Bultynck y Martin (1982); Ottone et al. (1992); Ottone et al. (1995); Manca et al. (1995) y Rubinstein (1997), mientras que trabajos de estudios integrados de palinomorfos de cuencas argentinas y esquemas bioestratigráficos son los de Ottone (1999); Rubinstein et al. (1999); Rubinstein y Toro (1999; 2001; 2002); Rubinstein (2001); Vergel et al. (2002); Rubinstein (2003); Toro et al. (2003); Vecoli et al. (2006); Waisfeld et al. (2006); Rubinstein et al. (2007 a, b); Tortello et al. (2008) y de la Puente y Rubinstein (2009). La sucesión ordovícica-silúrica de Sierra de Zapla en Sierras Subandinas fue estudiada por Rubinstein (2003; 2005) y Rubinstein y de la Puente (2008) mientras que Rubinstein y Vaccari (2001; 2004) estudian microfloras de ese periodo en la Puna y Achab et al. (2006) estudian las microfloras ordovícicas en el Sistema de Famatina.
Aspectos Generales
La Sierra de Zenta, situada entre las provincias geológicas Cordillera Oriental y Sierras Subandinas y con un conocimiento geológico detallado relativamente escaso, fue incluida dentro del ámbito de la Cordillera Oriental en los primeros esquemas de ordenamiento de las unidades geológicas en Argentina (Brackebush, 1892; Keidel, 1925; Groeber, 1938). Posteriormente, Ramos (1999) adoptó para las Sierras Subandinas, un esquema que las divide en dos sectores bien diferenciados, el Sistema Subandino s.s. y el Sistema Interandino u Occidental, incluyendo a la Sierra de Zenta y tomada como una zona transicional entre la Cordillera Oriental y el Sistema Subandino s.s. Astini (2008) remarca un evidente desarrollo de secuencias ordovícicas en la Cuenca Andina Central donde define tres subregiones (faja oriental, central y occidental), con la Sierra de Zenta incluida en la faja central.
La problemática surgida en torno a la asignación de edades a algunos niveles aflorantes en la sierra, como por ejemplo las unidades de Abra de Zenta que Starck et al. (1993 a, b) y Starck (1996) asignan al siluro-devónico y que en este trabajo se reasignan como ordovícicas, fue analizada durante este trabajo a partir de datos palinológicos. En base al análisis de los datos y las observaciones de campo, se acepta para la Sierra de Zenta una ubicación en un sector de características geológicas transicionales, con niveles comparables con unidades de Cordillera Oriental y de Sierras Subandinas.
Marco estratigráfico regional - Estratigrafía de la Sierra de Zenta
La columna estratigráfica (Figura N° 2) muestra las unidades geológicas de Sierra de Zenta en comparación con las de Cordillera Oriental y Sierras Subandinas. Las unidades del sector norte de la Sierra de Zenta son equiparadas con las de Cordillera Oriental y se usa la misma nomenclatura, mientras que las unidades del sector sur son parcialmente comparables con las de Sierras Subandinas y denominadas informalmente como Unidades Formacionales I, II y III. La elaboración de trabajos sedimentológicos de detalle y de estratigrafía secuencial en toda la región, permitirán una mejor caracterización de las unidades aquí descriptas, sin descartar la posibilidad de que éstas puedan nominarse como nuevas unidades en el Ordovícico del Noroeste Argentino.
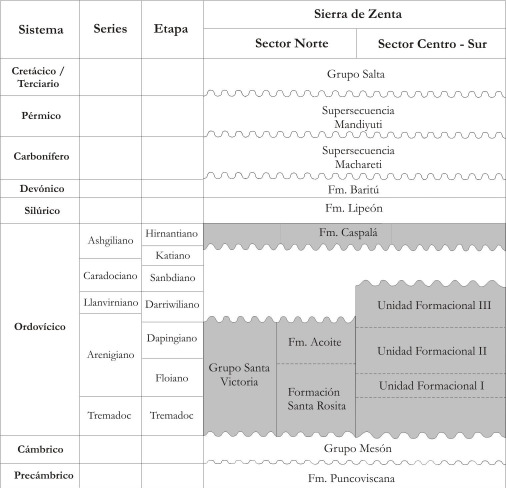
Figura 2. Columna Estratigráfica General para la Sierra de Zenta, con las unidades diferenciadas para el sector norte y centro sur de la misma.
A continuación se mencionan brevemente los datos de afloramientos de las unidades identificadas en la Sierra de Zenta.
Formación Puncoviscana (s.l.) (Turner, 1960). La Hoja Geológica 2366 IV (González et al, 2004) describe afloramientos en el sector norte del Abra de Zenta; Amengual y Zanettini (1973) mencionan afloramientos al Oeste, Sur y Norte de la región de Cianzo y Caspalá y Aceñolaza y Aceñolaza (2005; 2008) describen en el tramo superior de la Cuesta de Aparzo y camino a Zenta, niveles arenosos fuertemente plegados asignados a esta formación, donde identificaron Helminthoidichnites tenuis.
Grupo Mesón (Turner, 1960). González et al. (2004) describen afloramientos del grupo, generalmente asociados a la Formación Puncoviscana en la Sierra de Zenta y Sierra de Aparzo; Amengual y Zanettini (1973) los mencionan en el sector occidental y noroeste de la comarca de Cianzo y Caspalá diferenciando las formaciones Lizoite y Campanario y Sánchez y Salfity (1999) reconocen la Formación Lizoite en el Abra de Zenta, debajo de niveles tremadocianos.
Grupo Santa Victoria (Turner, 1960). González et al. (2004) mencionan afloramientos del Grupo Santa Victoria dispuestos en una franja con orientación NE - SW desde el norte de la sierra hasta el Abra de Zenta. En la Sierra de Zenta el Grupo Santa Victoria se superpone en discordancia a depósitos del Grupo Mesón o de la Formación Puncoviscana y es cubierto por depósitos del Paleozoico Medio y Superior. Si bien Starck et al. (1993 a, b) y Starck (1996) asignan los niveles aflorantes en el Abra de Zenta al Devónico, en este trabajo se reconocen en el Perfil Abra de Zenta Abra Límite las formaciones Santa Rosita y Acoite (Ordovícico) en Abra de Zenta, con fósiles que confirman edad tremadociana en el sector basal y floiana-dapingiana en el sector medio a superior y edades devónicas en Abra Límite. En base a esto, las edades devónicas mencionadas por Starck et al. (1993 a, b) y Starck (1996) para el Abra de Zenta corresponderían a la localidad de Abra Límite, donde los palinomorfos permiten asignar edades devónicas.
Formación Santa Rosita (Turner, 1960). Amengual y Zanettini (1973) registran afloramientos de la formación en el sector noreste de Cianzo y al sur de la localidad de Ocumazo mientras que Astini (2008) remarca que los miembros de la Formación Santa Rosita representativos de regiones próximas a la Quebrada de Humahuaca, pierden definición hacia el norte y este de la misma, como en el sector Zenta - Santa Ana, donde menciona afloramientos con espesores mayores a 3000 m pero sin una clara subdivisión de miembros. En este trabajo se identificó esta formación en el sector basal del Abra de Zenta en contacto por falla con el Grupo Mesón. Las características litológicas de la formación corresponden a las de su área tipo, aunque se observaron algunas diferencias en cuanto al espesor, siendo en esta sección de menos de 30 m, sin base determinada. En estos afloramientos Aceñolaza et al. (2003) identificaron una fauna que incluye trilobites, graptolites, braquiópodos, moluscos, equinodermos, con trilobites de la Biozona de Kainella en una asociación afín a las descriptas para el Tremadociano inferior alto y Tremadociano superior bajo de otras localidades de Cordillera Oriental. (Tortello y Aceñolaza, 2008).
Formación Acoite (Turner, 1960). En este trabajo fue identificada en Abra de Zenta, superpuesta a la Formación Santa Rosita y con una zona de discontinuidad estructural que la separa de los niveles superiores de la Formación Caspalá. En esta formación se identificaron fósiles que permitieron asignar a los niveles portadores una edad Arenigiano Inferior a Medio (Turner, 1960; Turner y Mon, 1979; Astini et al., 2004).
Formación Caspalá (Starck et al., 1992 a). Starck et al. (1992 a y Starck (1995) definen esta formación para los niveles que apoyan transgresivamente sobre sedimentitas arenigianas aflorantes en la región de Santa Ana y que son cubiertos de modo discordante por la Formación Lipeón. En este trabajo se la identificó en Abra Límite, al tope del Perfil N° 1 separada por una zona de fallas de los niveles devónicos suprayacentes (Vergel et al., 2008 a) y en Laguna Verde, en el tope del Perfil N° 5, superpuestos mediante discordancia a la Unidad Formacional III.
Depósitos siluro-devónicos. González et al. (2004) registran niveles silúricos-devónicos al este y sur del pliegue del Hornocal y hacia el sur y sureste de Santa Ana rodeando los depósitos ordovícicos del Grupo Santa Victoria de la Sierra de Zenta. Amengual y Zanettini (1973) identifican las formaciones Lipeón y Baritú al este y sudeste de la región de Cianzo y Caspalá. En el Abra de Zenta, Starck et al. (1992 a, b, 1993 a, b) y Starck (1995, 1996, 2008) describen depósitos siluro devónicos de las formaciones Lipeón y Baritú en la región de Abra de Zenta Caspalá. Starck (1995, 1996) menciona la Supersecuencia Las Pavas en diferentes sectores de la sierra Vergel etal. (2008 a) y Noetinger y di Pasquo (2008) e identifican en el Abra Límite asociaciones palinológicas de edad devónica que confirman parcialmente la presencia de la Supersecuencia Las Pavas y por encima de ella niveles diamictíticos discordantes atribuidas al Pennsylvaniano por sus esporas y granos de polen, confirmando la presencia de la discordancia devónico carbonífera. Al respecto, en este trabajo se confirma que la localidad citada como Abra de Zenta incluye niveles exclusivamente ordovícicos inferior a medio, mientras que el Abra Límite incluye niveles ordovícicos tardíos, devónicos y carboníferos, por lo tanto la localidad que Starck et al. (1993 a, b) y Starck (1996; 2008) denominan Abra de Zenta, sería lo que aquí se denomina Abra Límite.
Depósitos del Paleozoico Superior. Amengual y Zanettini (1973) y González et al. (2004). señalan afloramientos de los grupos Macharetí y Mandiyutí en una franja al sur de la Sierra de Hornocal, discordantes sobre niveles siluro devónicos. Starck et al.(1993 a, b) y Starck (2008) describen afloramientos de las formaciones Tupambi y Tarija en una franja de dirección aproximada N - S, que desde el Abra de Zenta continúa hasta la latitud de Caspalá, donde se adelgaza hacia el oeste. Vergel et al. (2008 a, b) y di Pasquo y Vergel (2008) reconocieron niveles atribuidos al Pennsylvaniano en las localidades de Abra Límite, Abra Rocío y Abra Blanca.
Grupo Salta (Turner, 1959). Amengual y Zanetini (1973) reconocen afloramientos del Grupo Salta en el área de Cianzo y Caspalá, al sur de la Sierra de Zenta, cerca de la localidad de Santa Ana y en la Serranía de Hornocal. Starck et al. (1993 a) mencionan la Formación Yacoraite en el Norte del Abra de Zenta y el Subgrupo Pirgua en el Sur, remarcando que el Abra de Zenta habría conformado un depocentro originado por fallas normales.
Nuevas unidades formacionales identificadas en Sierra de Zenta.
Unidad Formacional I. Esta unidad fue identificada en el sector central de la Sierra de Zenta, en la localidad de Abra Buey Ojo, en el Perfil N° 2 que pasa transicionalmente a la Unidad Formacional II. La Unidad Formacional I es una secuencia heterolítica con areniscas finas fangolíticas y estratificación mediana a gruesa, con frecuentes intercalaciones de areniscas finas con macrofósiles (Lingula sp.) e icnitas (Cruziana, Skolithos) sin valor bioestratigráfico. La unidad fue comparada con la Formación Zanjón aunque se diferencia principalmente en el desarrollo de mayores espesores, aproximadamente 150 m en este sector. La edad floiana de esta Unidad Formacional es determinada en base a la microflora incluida.
Unidad Formacional II. Con afloramientos identificados al norte de Abra Buey Ojo, en el tope del Perfil N° 2 y en la base del Perfil N° 3, la unidad es una secuencia de areniscas fangolíticas intercaladas con potentes bancos de areniscas finas a medianas portadoras de las trazas Lockeia isp., Protovirgularia dichotoma, Skolithos linearis y elementos del grupo Cruziana rugosa (Aceñolaza y Aceñolaza, 2007). La unidad se comparó con la Formación Labrado aunque tiene mayores espesores, aproximadamente 200 m, y un arreglo litológico que no permitió diferenciar unidades menores. La edad determinada en base a la microflora es Dapingiano a probable Darriwiliano.
Unidad Formacional III. Esta unidad se identificó en gran parte del sector central de la Sierra de Zenta, en la parte superior del Perfil N° 3 y en los Perfiles N° 4 y 5. En este último perfil en Laguna Verde, la unidad (citada como Formación Capillas por Aráoz et al., 2008) está en contacto discordante con la Formación Caspalá. La unidad se describe como una sucesión de areniscas finas fangolíticas, con estratificación y frecuentes intercalaciones de capas o lentes de areniscas finas muy duras, calizas y niveles de coquinas. La unidad fue comparada con la Formación Capillas, aunque también con mayores espesores de alrededor de 380 m. En Laguna Verde, Heredia y Aceñolaza (2005) y Aceñolaza y Heredia (2008) registraron conodontes como Erismodus aff. quadridactylus que asigna una edad Darriwiliano Sanbdiano para los niveles portadores mientras que la microflora incluida permite asignar a la unidad una edad darriwiliana. Esta unidad es comparable con la parte superior del perfil Ordovícico de Los Colorados, sobre el flanco oeste de la Cordillera Oriental Jujueña (Astini et al., 2004).
Perfiles de Sierra de Zenta
Los niveles ordovícicos aflorantes en la Sierra de Zenta se estudiaron en dos sectores, uno al norte descripto en el Perfil N° 1 donde se describen los niveles ordovícicos más antiguos identificados y otro en el sector central de la sierra, confeccionado en base a cuatro perfiles parciales, Perfiles N° 2, 3, 4 y 5.
El Perfil N° 1 se inicia en Abra de Zenta y se desarrolla hacia el sur hasta Abra Límite donde se observa un contacto tectónico con areniscas y conglomerados de la Formación Caspalá (Ordovícico tardío) y por encima se encuentra en discordancia niveles interpretados como Devónico Temprano a Medio y Pennsylvaniano (Vergel et al., 2008 a, b; di Pasquo y Vergel, 2008).
El perfil integrado se desarrolla entre Abra Buey Ojo y Laguna Verde y se observó una correspondencia parcial con las unidades descriptas para el Ordovícico de la Sierra de Zapla (Nieniewiski y Wleklinski, 1950; Monaldi et al., 1986; Astini y Marengo, 2006; Astini, 2008). En el perfil se identificaron tres unidades que se definieron informalmente como Unidades Formacionales I, II y III. En el tope del perfil, en Laguna Verde, se identificaron las Formaciones Caspalá y Lipeón superpuestas a la Unidad Formacional III.
Perfil N° 1 (Figura N° 3). Este perfil se encuentra ubicado entre el Abra de Zenta y el Abra Límite. Los niveles ordovícicos de este perfil se asignaron a las formaciones Santa Rosita y Acoite (Ordovícico inferior) y a la Formación Caspalá (Ordovícico superior) y son superpuestos por niveles de edades devónica y pennsylvaniana.
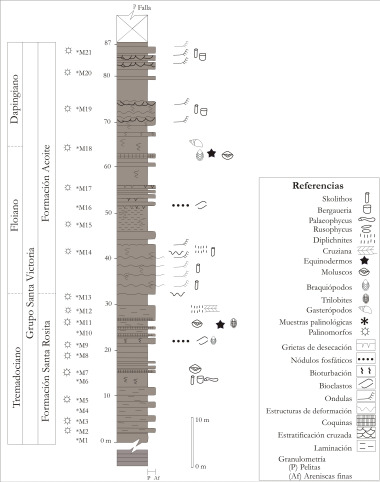
Figura 3. Perfil N° 1, descripto en el sector norte de la sierra, entre Abra de Zenta y Abra Límite.
En el sector inferior del perfil, la Formación Santa Rosita incluye trilobites de la Biozona de Kainella meridionalis del Tremadociano Inferior tardío a Superior (Aceñolaza et al., 2003; Tortello y Aceñolaza, 2008), moluscos rostroconchos, gasterópodos, braquiópodos y equinodermos asociados a trazas fósiles que son comunes en la mayoría de los niveles y que fueron referidas a la "icnofacies de Cruziana" (Aceñolaza et al., 2003). Entre los palinomorfos se recuperaron acritarcos, algas prasinofíceas, criptosporas y un ejemplar de quitinozoo, con una importante variación en la cantidad y diversidad.
El sector inferior del perfil fue interpretado como una zona de plataforma, con un arreglo de shoreface, cercano a la base de máximo de olas y somerización a partir de los bancos de areniscas finas que aumenta su potencia hacia arriba hasta los primeros niveles de coquinas, indicando una lower offshore a lower shoreface y en el sector medio y superior del perfil se interpreta una somerización con exposición subaérea y por encima una nueva profundización (Aceñolaza et al., 2003).
Perfil N° 2 (Figura N° 4).Este perfil se inicia en Abra Buey Ojo y es el primero de los perfiles parciales del sector central de la sierra, en el se identificaron las Unidades Formacionales I y II. Son escasos los macrofósiles recuperados entre los que se incluyen gasterópodos, graptolites y braquiópodos linguliformes indeterminados (Aráoz, 2009). Entre los palinomorfos recuperados se identificaron acritarcos asociados a prasinofíceas, criptosporas y quitinozoos indeterminados. La microflora de la Unidad Formacional I permite asignarle una edad floiana mientras que la Unidad Formacional II no contiene elementos palinológicos suficientes para asignar su edad.
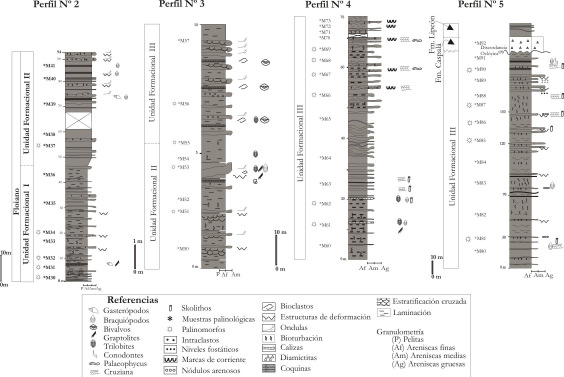
Figura 4. Perfiles N° 2-3-4 y 5, descriptos en el sector centro sur de la sierra, entre Abra de Buey Ojo y Laguna Verde.
El sector inferior a medio del perfil representa un paleoambiente más profundo que el resto de la secuencia con un incremento de granulometría a la mitad del perfil que indica una somerización. La mayor participación pelítica en algunos sectores refleja una menor energía y deposición por procesos de decantación mientras que los niveles arenosos y bases deformadas y erosivas con óndulas en el techo, son indicadores de una somerización de la secuencia.
Perfil N° 3 (Figura N° 4). En este perfil se identificaron las Unidades Formacionales II y III separadas por un sector transicional, con graptolites como únicos fósiles identificados (Didymograptus sp., Expansograptus sp., Tetragraptus sp. y Baltograptus sp.) que permiten considerar una edad arenigiana (Aráoz y Aceñolaza, 2004). Los palinomorfos son relativamente abundantes pero con una diversidad baja a media, entre los que se incluyen acritarcos, prasinofíceas, quitinozoos y criptosporas.
El aumento de los bancos arenosos de mayor granulometría en la parte media a superior del perfil indicaría una somerización con respecto al sector inferior mientras que las coquinas y los nódulos arenosos redondeados asociados a los bancos de areniscas más gruesas indican eventos de alta energía.
Perfil N° 4 (Figura N° 4).La Unidad Formacional III fue descripta en toda la extensión del perfil y se destaca que los palinomorfos identificados son más abundantes y variados que en las microfloras de los perfiles anteriores e incluyen acritarcos, quitinozoos y escasas criptosporas.
El sector inferior del perfil se interpreta como un ambiente marino de plataforma interna, con capas de tormenta, con un gradual incremento de la fracción arenosa hacia la parte superior y un cambio de trazas de Cruziana a tubos verticales como Skolithos, que indica una somerización gradual en estos niveles. La presencia de acritarcos asociados a quitinozoos y ausencia de criptosporas indican en conjunto paleoambientes alejados del continente.
Perfil N° 5 (Figura N° 4). Este perfil es el último de los perfiles parciales y se encuentra en Laguna Verde incluye los niveles ordovícicos más jóvenes descriptos. Los niveles pertenecen a la Unidad Formacional III sobre los cuales suprayace en forma discordante las formaciones Caspalá y Lipeón. En la parte superior de esta potente secuencia arenosa hay una importante cantidad y variedad de trazas fósiles entre los que se destacan pavimentos de Protovirgularia dichotoma, Cruziana (del grupo rugosa), Lockeia y Monomorphichnus, asociados a niveles con abundantes Skolithos (Aceñolaza y Milana, 2005; Heredia y Aceñolaza, 2005; Aceñolaza et al., 2006; Aceñolaza y Aceñolaza, 2007; Aceñolaza y Heredia; 2008 y Heredia et al., 2008). En el sector superior del perfil Heredia y Aceñolaza (2005), Aceñolaza y Heredia (2008) y Heredia et al. (2008) registran el conodonte Erismodus aff. quadridactylus que ratifica una edad darriwiliana alta/sandbiano baja para los niveles portadores. Los palinomorfos muestran una marcada disminución en cuanto a la cantidad y diversidad taxonómica con respecto a los niveles anteriores.
Para este perfil se puede interpretar un depósito en ambientes de plataforma externa que someriza en la parte superior donde hay un aumento de niveles de areniscas y granulometrías más gruesas. La presencia de trazas penetrativas como Skolithos en las fracciones más gruesas de areniscas sugiere una somerización y mayores energías.
Bioestratigrafía - Discusiones
El análisis de la distribución estratigráfica de los palinomorfos de los niveles ordovícicos de la Sierra de Zenta, permitió diferenciar asociaciones microflorísticas basadas en la presencia (aparición/ desaparición) y abundancia de especies de acritarcos. Aunque fueron identificados otros palinomorfos como algas prasinofíceas, quitinozoos y criptosporas, su presencia es menos representativa por lo que la diferenciación de las asociaciones se basa en los acritarcos. Las asociaciones definidas son ASZ1, ASZ2, ASZ3, ASZ4 y ASZ5, de las cuales ASZ1, ASZ2 y ASZ3 se identifican en el Perfil N° 1 mientras que las ASZ4 y ASZ5 en los Perfiles 2, 3, 4 y 5.
El significado cronoestratigráfico de las asociaciones se establece en base a las comparaciones y correlaciones con las zonaciones palinológicas previamente establecidas a nivel regional y mundial y que disponen de controles a partir de otras evidencias paleontológicas o geológicas asociadas. La comparación de las asociaciones con las de localidades del Noroeste Argentino permitió establecer solamente correspondencias parciales.
ASZ1 (M2 M12)
La ASZ1 se define en el sector inferior a medio del Perfil N° 1, en niveles de la Formación Santa Rosita, para la cual los trilobites de la Biozona de Kainella meridionalis asignan una edad Tremadociano Inferior Tardío (Figura N° 5, 6 y 9). La ASZ1 se define por la presencia exclusiva de los acritarcos Acanthodiacrodium angustum, A. spinum, A. ubuii, Balthisphaeridium sp. 1, Cristallinium cambriense, C. ovillense, Cymatiogalea cuvillieri, C. velifera, Saharidia downieii, S. fragilis, Vulcanisphaera africana y V. britannica. Otros acritarcos presentes son: Acanthodiacrodium spp., Cymatiogalea membranispina, C. spp., Dasydiacrodium spp., Eliasum llaniscum, Eupoikilofusa sp.1, Ferromia pelita, Micrhystridium spp., Leiofusa spp, Polygonium spp., V. tuberata, V. turbata y fusiformes indeterminados. Además hay prasinofíceas (Cymatiosphaera spp., Leiosphaeridia spp., Pterospermella sp. 1 y P. spp.) y criptosporas indeterminadas.
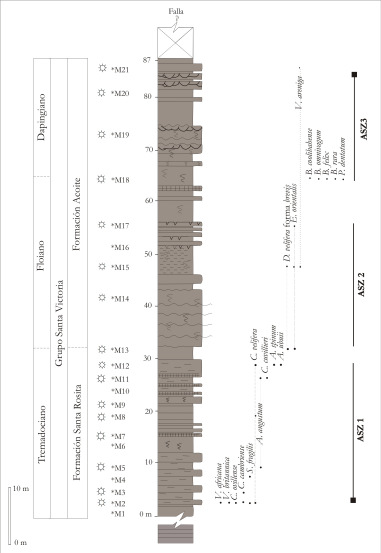
Figura 5. Distribución de las asociaciones palinológicas ASZ1, ASZ2 y ASZ3, definidas en el Perfil No. 1 y distribución de las especies más representativas incluidas en las mismas.
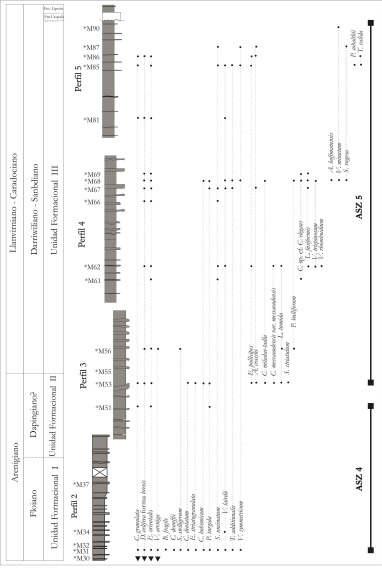
Figura 6. Distribución de las asociaciones palinológicas ASZ4 y ASZ5 definidas en los Perfiles No. 2, 3, 4 y 5 y distribución de las especies de acritarcos más representativas incluidas en las mismas.
La ASZ1 tiene una baja diversidad taxonómica, con un claro dominio de acritarcos y muy baja proporción de prasinofíceas y criptosporas. Entre los acritarcos hay una proporción relativamente alta de formas complejas como Acanthodiacrodium, Dasydiacrodium y Polygonium típica de asociaciones del Ordovícico más temprano, al igual que la aparición de diferentes especies de Vulcanisphaera (Servais et al., 2004). Se considera que Eliasum es producto del redepósito por su escasez y mala preservación y por encontrarse en una asociación con características que representan una edad tremadociana más que cámbrica.
La ASZ1 fue comparada con las microfloras de otras localidades del Noroeste Argentino de edades cámbricas y ordovícicas y la mejor equivalencia se estableció con la Asociación 3 de los niveles tremadocianos de la Quebrada de Moya (Aráoz y Vergel, 2006). Si bien las formas en común son escasas (A.angustum y C. membranispina) ambas asociaciones muestran una similar diversificación genérica y específica, con abundancia de acritarcos diacromorfos y poligonomorfos y aparición de formas típicamente tremadocianas. La microflora descripta por Manca et al. (1995) para el Tremadociano inferior de Nazareno también muestra una diversidad de géneros y especies de acritarcos similar a ASZ1, con algunos taxones en común como: Acanthodiacrodium spinum (en Nazareno, A. sp. cf. A. spinum), Cymatiogalea velifera (en Nazareno, C. sp. cf. C. velifera), especies indeterminadas de Leiofusa, Micrhystridium (en Nazareno, varias especies identificadas), Polygonium spp. (en Sierra de Zenta, varias especies identificadas) y especies de Vulcanisphaera (en Nazareno, Vulcanisphaera sp.). La Asociación Acanthodiacrodium angustumVulcanisphaerabritannica del Tremadociano temprano del Norte de Africa (Vecoli, 1999) incluye entre otras formas Acanthodiacrodium angustum, A. ubuii, Polygonium gracile y Vulcanisphaera britannica comunes con la ASZ1. Otra equivalencia parcial se establece con la Asociación de acritarcos Zona 5, del Tremadociano de Irán (Ghavidel-syooki y Vecoli, 2008) con la cual comparte las formas S. fragilis, A. angustum, V. africana y C. membranispina y ambas carentes de formas típicamente cámbricas, excepto E. llaniscum.
Edad de la asociación. Acanthodiacrodium angustum es un taxón diagnóstico del Tremadociano más bajo a nivel mundial y es tomado como un indicador de la posición del límite Cámbrico - Ordovícico. Además Saharidia fragilis, Vulcanisphaera africana y Cymatiogalea tienen registros desde el Cámbrico aunque son típicamente abundantes y comunes en asociaciones del Tremadociano a nivel mundial. Acanthodiacrodium spinum, A. ubuii, Cristallinium ovillense y Saharidia fragilis tienen sus registros más altos en el Tremadociano y son diagnósticas del Tremadociano Acanthodiacrodium ubuii y Vulcanisphaera britannica. La presencia de A. angustum, A. ubuii, Cymatiogalea velifera, C. cuvillieri, C. membranispina y Cristallinium ovillense asociado a la abundancia de especies de Vulcanisphaera no permiten considerar una edad más joven que Tremadociano Inferior, dato aportado también por los trilobites de la Biozona de Kainella meridionalis.
Observaciones. Martin (1993) y Vecoli (2000) consideran que el dominio de formas simples como Saharidia en una asociación de baja diversidad como en M2, es un indicador de paleoambientes relativamente someros. Mientras que Vecoli (1999) considera que Acanthodiacrodium angustum es una forma sensible a las condiciones paleoambientales y sugiere ambientes proximales y someros. El dominio de formas pequeñas como Micrhystridium (M7 y M9) es también un reflejo de asociaciones proximales. La relativa abundancia de formas complejas como Polygonium, Vulcanisphaera o Cymatiogalea (M2) se interpreta como de ambientes abiertos y a cierta distancia de la playa (Vecoli, 2000). Las criptosporas permiten inferir una proximidad relativa a zonas costeras o paleoambientes continentales. Los datos obtenidos a partir del análisis palinológico de la ASZ1 son coincidentes con los datos aportados por la sedimentología del perfil, indicando la presencia de paleoambientes someros y una relativa cercanía al continente.
ASZ2 (M13 M17)
La ASZ2 fue definida en el sector medio del Perfil N° 1, en la Formación Acoite (Figura N° 5, 6 y 9). La ASZ2 se define por la primera aparición de los acritarcos Cymatiogalea granulata, Dactylofusa velifera forma brevis, Eisenackidium orientalis, Eisenackidium sp. 1, Eupoikilofusa spp., Polygonium gracile y Vavrdovella areniga y el primer registro de quitinozoos (Conochitina sp.) Otros acritarcos identificados y registrados en la asociación anteriormente descripta son: Acanthodiacrodium spp., Micrhystridium sp., Polygonium sp. y fusiformes indeterminados. Además hay una muy baja cantidad de prasinofíceas (Pterospermella sp. 1) y criptosporas indeterminadas.
La ASZ2 se caracteriza por la escasa cantidadde ejemplares y una baja diversidad de especimenes, con pocas de las formas típicas tremadocianas descriptas en la ASZ1, pero con taxones características del lapso Floiano Dapingiano. Se destaca la aparición de Dactylofusa velifera forma brevis, para la cual se ha considerado el registro más bajo en el Arenigiano inferior, en la Zona de Tetragraptus akzharensis (Rubinstein et al., 1999). Eisenackidium orientalis está ampliamente distribuida en el Floiano de la Cuenca Andina Central y es una especie guía de esta edad en el NOA con el registro más antiguo en el Tremadociano Tardío de la region de Pascha-Incamayo (de la Puente et al. 2009). Vavrdovella areniga es otra forma cuyos registros controlados por otros fósiles son a partir del Arenigiano, aunque hay registros en el Tremadociano. Cymatiogalea granulata aparece también en el Arenigiano.
La escasa cantidad de formas identificadas en la ASZ2 no permite una buena correlación con otras asociaciones, aunque la presencia de elementos comunes en toda la región como Cymatiogalea granulata, Dactylofusavelifera forma brevis, Eisenackidium orientalis y Vavrdovella areniga permiten compararlas. Estos taxones fueron identificados en otras secciones del Noroeste Argentino, como en la Formación Zanjón de la Sierra de Zapla, con una edad floiana (Rubinstein y de la Puente, 2008) aunque se diferencia de la ASZ2 porque en ésta no se registra la mayoría de los taxones presentes en la Formación Zanjón. Achab et al. (2006) mencionan la presencia de estas especies en las formaciones Suri y Los Molles del Sistema de Famatina, a las que atribuyen edades Stage 2 y Stage 3 (Floiano y Dapingiano). Otro ejemplo son las "Asociaciones Inferior y Superior" (AL y AU) reconocidas por Ottone etal. (1992) en la Formación Acoite de las secciones de Los Colorados y Sierra de Cajas, que tienen en común con la ASZ2 los acritarcos: Polygonium gracile, Cymatiogalea granulata y Eisenackidium sp. A (posteriormente E. orientalis por Rubinstein en Rubinstein et al., 1999). Sin embargo, en la ASZ2 no se identificaron otras formas presentes en la AU, claramente arenigianas.
En el sector entre las ASZ1 y ASZ2 se encontraría la transición tremadociana floiana que no puede ser determinada con mayor detalle por la ausencia de formas típicas de este lapso.
Edad de la asociación. La presencia de Dactyolfusa velifera forma brevis y Cymatiogalagranulata indicarían una edad floiana a los niveles que contienen la ASZ2, sin embargo no se puede descartar un Tremadociano tardío por la presencia de Eisenackidium orientalis. La presencia del quitinozoo también indica una edad no más antigua que tremadociana superior.
Observaciones. La somerización observada en el sector medio del perfil se refleja en una menor diversidad de la microflora con respecto a los niveles inferiores. El registro de Vavrdovella areniga aún en ausencia de otras formas típicas de peri-Gondwana (Arbusculidium, Striatotheca y Coryphidium) permite sugerir una afinidad con esta provincia de agua fría. Se remarca también la ausencia de formas típicas que se correspondería con la edad propuesta. En ambos casos se podrían considerar causas paleoambientales o paleogeográficas de carácter local.
ASZ3 (M18 M21)
La ASZ3 fue definida en el sector superior del Perfil N° 1, en niveles de la Formación Acoite (Figura N° 5, 6 y 9) y se define por la presencia exclusiva de los acritarcos Balthisphaeridium coolibahense, B. omnivagum, B. sp. cf. B. crinitum, Barakella rara, B. felix y Polygonium dentatum. La ASZ3 se caracteriza por la primera aparición de Stelliferidium sp. y Tectitheca sp. cf T. additionalis. Otros taxones de acritarcos presentes pero registrados en las anteriores asociaciones son Cymatiogaleamembranispina, C. spp., Eupoikilofusa sp. 1, Micrhystridium spp., Polygonium sp., Vavrdovella areniga y Vulcanisphaera turbata. También se identificaron restos de prasinofíceas (Pterospermella sp. 1) y criptosporas indeterminadas.
La ASZ3 es pobre cuantitativamente y de muy baja diversidad palinológica, compuesta únicamente por acritarcos. Se destaca la aparición de formas típicamente arenigianas como Balthisphaeridium coolibahense, cuyo registro más bajo es en el Yushaniano Medio, en la Zona del graptolite A. suecicus (Yin, 1995); Balthisphaeridium omnivagum para la cual sólo se conocen registros en el Arenigiano, Barakella cuyo FAD (First Appearance Data) es en la base del Dapingiano (Li et al., 2002; Servais et al., 2008) aunque Rubinstein et al. (2007 b) lo registran en el Floiano tardío de Cordillera Oriental.
La escasa presencia de formas en la ASZ3 sólo permite una ligera comparación con la asociación identificada en la Formación Molles del Sistema de Famatina (Achab et al., 2006) y que tiene en común las formas Balthisphaeridium coolibahense y Balthisphaeridium sp. cf. B. omnivagum.
Edad de la asociación. Los taxones de la ASZ3 permiten considerar para la misma una edad dapingiana, principalmente por la presencia del género Barakella, aunque no se descarta una edad floiana tardía. Se puede inferir que la transición floiana - dapingiana está incluida en este sector del perfil, entre los niveles que incluyen las ASZ2 y ASZ3.
ASZ4 (M30 M51)
La ASZ4 fue definida en el Perfil N° 2 en niveles de las Unidades Formacionales I y II y en el sector basal del Perfil N° 3 en la Unidad Formacional II (Figura N° 7, 8 y 9). La ASZ4 se define por la primera aparición de los acritarcos Balthisphaeridium fragile, Coryphidiumbohemicum, Cristallinium dentatum, Cymatiogalea deunffii, Dasydiacrodium sp., Eupoikilofusastriatogranulata, Multiplicisphaeridium? spp., Picostellaturgida, Pirea sp. cf. P. colliformis, Polygonium spp., Stellechinatumuncinatum, Stellechinatum sp. 1, Stelliferidiumstelligerum, Striatotheca principalis, Tectithecaadditionalis, T. sp. cf. T.additionalis, Veryhachiumlairdii y V. symmetricum. Otros acritarcos presentes registrados en las asociaciones ya descriptas son: Acanthodiacrodium spp., Cymatiogaleagranulata, Cymatiogalea sp., Dactylofusa velifera forma brevis, Eisenackidium orientalis, Eisenackidium sp. 1, Leiofusa sp., Micrhystridium spp., Polygonium gracile, Stelliferidium sp., Vavrdovellaareniga y Vulcanisphaeratuberata. Además se encontraron prasinofíceas (Cymatiosphaera spp., Leiosphaeridia spp. y Pterospermella sp. 1), criptosporas y quitinozoos (Euconochitina sp. y Eremochitina sp. y otros indeterminados).

Figura 7. Perfiles No. 1, 2, 3, 4 y 5 con la distribución de las asociaciones palinológicas ASZ1, ASZ2, ASZ3, ASZ4 y ASZ5.
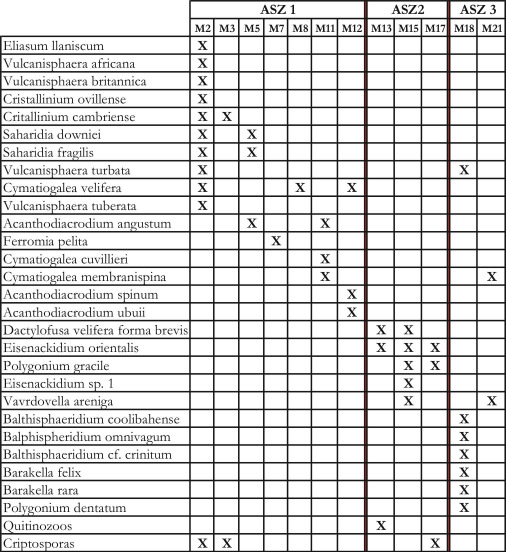
Figura 8. Cuadro de distribución de especies de acritarcos seleccionados, quitinozoos y criptosporas que componen las asociaciones palinológicas ASZ1, ASZ2 y ASZ3.
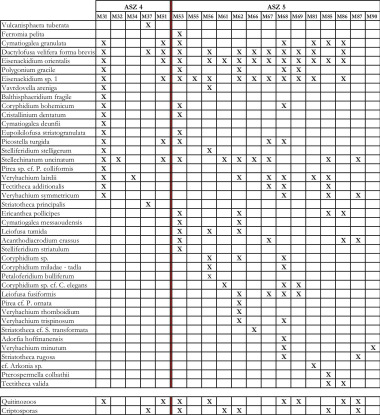
Figura 9. Cuadro con la distribución de especies de acritarcos seleccionados, quitonozoos y criptosporas que componen las asociaciones palinológicas ASZ4 y ASZ5.
La ASZ4 se caracteriza por la mayor abundancia y diversidad de palinomorfos, especialmente acritarcos y quitinozoos y por mostrar un marcado cambio en los componentes con respecto a las asociaciones ASZ1, ASZ2 y ASZ3 descriptas. Entre los componentes de la ASZ4 Cymatiogalea deunffii, Vavrdovella areniga, Veryhachium lairdii y los géneros Pirea, Striatotheca y Coryphidium son componentes de la asociación messasoudensis trifidum, por lo que podría considerarse parcialmente equivalentes a la misma. La asociación messasoudensis trifidum (Molyneux y Rushton, 1988; Fortey et al. 1991; Servais y Molyneux, 1997) caracteristica del Tremadociano Superior Floiano en localidades de Peri-Gondwanafue fue subdividida por Cooper et al. (2004) en cinco subasociaciones.
La ASZ4 se comparó con las Asociaciones 2, 3 y 4 definidas por Rubinstein et al. (1999) y Rubinstein y Toro (2001) para el Arenigiano temprano de Cordillera Oriental, en niveles de las Biozonas de Tetragraptus akzharensis y Baltograptus deflexus con los cuales tiene en común: Cymatiogala granulata, Dactylofusavelifera forma brevis, Eisenackidium orientalis, Vavrdovellaareniga además de Coryphidium y Striatotheca. La Asociación 5 definida para el Arenigiano medio en la Biozona de Didymograptus bifidus tiene en común con la ASZ4 los acritarcos Coryphidium bohemicum, Cristallinium dentatum, Cymatiogalagranulata, Picostella turgida, Striatotheca sp. cf. S. principalis, T. additionalis y V. lairdii. En trabajos recientes para la Formación Acoite (Rubinstein et al., 2007 b) se mencionan acritarcos que sugieren una correlación con la sub-asociación 5 de la asociación messasoudensis-trifidum (Arbusculidium filamentosum y Aureotesta clathrata var. simplex) aunque no registradas en la ASZ4. En la parte superior de la Formación Parcha, en la sección Pascha-Incamayo (Rubinstein et al., 2007 b) se describen formas de la asociación messaoudensis-trifidum, de las cuales están presentes en la ASZ4 Cymatiogalea deunffii y taxones acompañantes como Coryphidium, Vavrdovella y Acanthodiacrodium. Rubinstein et al. (2007 b) reconocen en la sección de Los Colorados, en niveles de la Biozona de Baltograptus deflexus, formas que sugieren una correlación con la sub-asociación 5 (base del Floiano) de la microflora messaoudensis-trifium, de las cuales solo Cymatiogalea granulata y Striatotheca principalis se encuentran en la ASZ4.
La Formación Zanjón, cuya edad floiana tardía - dapingiana temprana (Astini y Marengo, 2006) es restringida al Floiano por su microflora (Rubinstein, 2005; Rubinstein y de la Puente, 2008) tiene en común con la ASZ4 los acritarcos: Cymatiogalea granulata, Dactylofusa velifera forma brevis, Eisenackidium orientalis, Striatotheca principalis y Coryphidium, aunque en la ASZ4 no se identificaron formas diagnósticas como Cymatiogalea messaoudensis ni los géneros Vogtlandia o Rhopaliophora.
Las formaciones Suri (Floiano - Arenigiano inferior a medio) y Molles (Dapingiano / Darriwiliano - Arenigiano superior - Llanvirniano) del Sistema de Famatina (Achab et al., 2006) contienen microfloras ricas y diversas con pocas formas en común con la ASZ4, aunque se destaca la presencia de algunas con valor estratigráfico como Dactylofusa velifera forma brevis, Eupoikilofusa striatogranulata (como Dactylofusa striatogranulata), Striatotheca principalis y Vavrdovella areniga de la Formación Suri, Veryhachium lairdii de la Formación Molles y Eisenackidiumorientalis en ambas. En base a esto, se considera una similitud parcial con la microflora de la Formación Suri y una edad equivalente. La "Upper Acoite Palynological Assemblage" (AU) establecida por Ottone et al. (1992) para el Arenigiano medio de la Formación Acoite, tiene pocas formas en común (P. gracile, C. granulata, V. lairdii) y con escaso valor bioestratigráfico.
Edad de la asociación. Cymatiogalea deunffii es un taxón característico del límite Tremadociano Floiano; Dactylofusa velifera forma brevis se registra a partir del Arenigiano inferior; Eisenackidium orientalis es tipicamente floiano pero se registra desde el Tremadociano tardío y Picostella turgida tiene sus registros limitados al Arenigiano temprano. Otros taxones que aparecen por primera vez en el Arenigiano son Balthisphaeridium fragile, Cristallinium dentatum, Cymatiogalea granulata, Eupoikilofusa striatogranulata, Tectitheca additionalis y Striatothecaprincipalis. Coryphidium bohemicum, Veryhachium lairdii y V. symmetricum tienen su registro más bajo en el Yushanian temprano, aunque las dos primeras son citadas desde el Tremadociano.
La composición de la ASZ4 permite asignarle una edad floiana y considerar una cercanía límite Tremadociano Floiano por la presencia de Cymatiogalea deunffii. Esta especie junto con Coryphidium, Vavrdovella y Acanthodiacrodium permiten una correlación parcial con la sub-asociación 5 de la microflora essaoudensis-trifidum.
En comparación, tanto la ASZ4 como la ASZ2 se encuentran cercanas al límite Tremadociano Floiano, sin embargo, las composiciones son notablemente diferents, en base a esto se puede inferir para la ASZ4 una edad floiana y para la ASZ2 una probable edad floiana a tremadociana tardía.
Observaciones. En el sector inferior a medio del perfil la asociación palinológica es diversa y heterogénea e incluye una relativamente alta cantidad y diversidad de acritarcos y quitinozoos que indicarían condiciones marinas abiertas y alejadas de la costa. En la parte alta de la secuencia, en la cual se observa un incremento de la granulometría de los sedimentos, se registraron criptosporas y una ausencia de quitinozoos.que indicaría paleoambientes cercanos al continente.
La presencia de Cymatiogalea deunffii, Vavrdovella areniga, Coryphidium, Pirea, Striatotheca y Veryhachium permiten establecer afinidades con la Provincia Mediterránea o peri-Gondwana, que representan una asociación de aguas frías para el Ordovícico temprano (Vavrdova, 1974; Li Jun; 1987) aunque otras formas típicas de la provincia como Arbusculidium, Dicrodiacrodium y Frankea no fueron registradas en la asociación.
Si bien la ASZ4 tiene formas en común con la Formación Acoite (Cordillera Oriental), Formación Zanjón (Sierras Subandinas) y las formaciones Suri y Molles (Sistema de Famatina) se remarca que hay en la ASZ4 y en la Sierra de Zenta en general, una ausencia de las formas que son típicas del Arenigiano a nivel regional y mundial.
ASZ5 (M53 M90)
La ASZ5 fue identificada en el sector medio a superior del Perfil N° 3 y en los Perfiles N° 4 y 5 en niveles de la Unidad Formacional 3 (Figura N° 7, 8 y 9).
La ASZ5 se define por la primera aparición de los acritarcos Acanthodiacrodium crassus, Adorfiahoffmanensis, cf. Arkonia sp., Aureotesta? sp., C. miladae-tadla, Coryphidium sp. cf. C. elegans, C. spp., Cymatiogalea messaoudensis var. messaoudensis, Ericantheapollicipes, Leiofusafusiformis, Leiofusatumida, Petaloferidium bulliferum,cf. Picostella sp. 1, Pirea sp. cf. P. ornata, Poikilofusa spp., Priscogalea sp., Stelliferidium striatulum, Striatothecarugosa, Striatotheca sp. cf. S. transformata, Striatotheca spp., Tectitheca valida, Veryhachium minutum, V. rhomboidium y V. trispinosum. También se registran por primera vez los quitinozoos Euconochitina sp., Eremochitina sp. y Lagenochitina sp. y la prasinofícea Pterospermella colbathii.
Otros acritarcos registrados en las asociaciones anteriores son: Acanthodiacrodium spp., Barakella sp., Coryphidiumbohemicum, Cristalliniumdentatum, Cymatiogalea granulata, C. spp., Dactylofusavelifera forma brevis, Dasydiacrodium spp., Eisenackidiumorientalis, Eisenackidium sp. 1, Eupoikilofusa striatogranulata, Eupoikilofusa sp. 1, Fer romia pelita, Leiofusa spp., Micrhystridium spp., Multiplicisphaeridium? spp., Picostella turgida, Polygoniumgracile, Polygonium spp., Stellechinatum uncinatum, Stellechinatum sp. 1, Stelliferidium stelligerum, Stelliferidium sp., Tectitheca additionalis, T. sp. cf. T. additionalis, Vavrdovella areniga, Veryhachium lairdii y V. symmetricum. También se registran prasinofíceas (Cymatiosphaera sp., Leiosphaeridia spp., Pterospermella sp. 1 y Pterospermella spp.), quitinozoos (Conochitina sp. e indeterminados) y crisptosporas indeterminadas.
La ASZ5 se caracteriza por tener una gran cantidad y diversidad de acritarcos y una menor proporción de quitinozoos y prasinofíceas, aunque mayor con respecto a las otras asociaciones.
La ASZ5 se identificó en una amplia extensión de los perfiles y presenta una notable heterogeneidad a lo largo de los niveles portadores, donde se destacan muestras como la M68 o M62 que contienen la más alta diversidad y abundancia de formas, especialmente de acritarcos y quitinozoos y otras en las cuales los palinomorfos son muy escasos y poco variados. A pesar de estas diferencias, se remarca la amplia representación, en la mayoría de los niveles, de taxones como Dactylofusa velifera forma brevis, Eisenackidiumorientalis, Eisenackidium sp. 1, Vavrdovella areniga y Stellechinatum uncinatum.
Los taxones registrados en la ASZ5 tienen rangos de distribución amplios, desde el Floiano/ Dapingiano hasta el Darriwiliano o incluso algunos de ellos fueron registrados en edades tan jóvenes como Caradociano (Sanbdiano/Katiano inferior). También es notable en la ASZ5 la aparición de numerosas formas nuevas aunque no hay una desaparición notable de formas que permitan alguna consideración bioestratigráfica relevante.
Las asociaciones microflorísticas descriptas por Rubinstein et al. (1999) y Rubinstein y Toro (1999; 2001) para diferentes secciones de la Cordillera Oriental tienen en común con la ASZ5 varias formas de acritarcos: Adorfia hoffmanensis, Barakella sp., Cymatiogalea granulata, C. messaoudensis var. messaoudensis, Dactylofusa velifera forma brevis, Eisenackidium orientalis, Coryphidium miladae-tadla, Coryphidium spp., Picostella turgida, Stellechinatum uncinatum, Stelliferidium spp., Striatotheca spp., Tectitheca additionalis, Vavrdovella areniga, Veryhachium lairdii, Veryhachium trispinosum, V. rhomboidium. Estos taxones tienen una amplia distribución temporal que permiten asignar a la ASZ5 edades desde floianas a darriwilianas.
La ASZ5 fue comparada con las microfloras de niveles ordovícicos aflorantes en Sierra de Zapla, Sierras Subandinas analizadas por Rubinstein (2005) y Rubinstein y de la Puente (2008). Describen en la Formación Zanjón una microflora para la que consideran una edad floiana y que tiene acritarcos en común con la ASZ5: Cymatiogalea messaoudensis var. messaoudensis, Dactylofusavelifera, Eisenackidium orientalis, Striatotheca spp. y Tectitheca additionalis - valida. Para la microflora de la Formación Capillas consideran una edad máxima darriwiliana, con los acritarcos en común con la ASZ5: Ericanthea pollicipes, Cymatiogaleagranulata, Striatothecarugosa, Striatotheca sp., Tectithecaadditionalisvalida y Veryhachium trispinosum y Barakella. En la ASZ5 se registraron taxones con edades más jóvenes y no registrados en la Formación Capillas (Acanthodiacrodium crassus, Leiofusa tumida y Veryhachium rhomboidium) pero no incluye otras de importancia bioestratigráfica como Liliosphaeridium, Arbusculidium o Rophaliophora. Esto permite marcar que la mejor correspondencia de la ASZ5 es con la Formación Capillas aunque no es completa.
Achab et al. (2006) registran en las formaciones Suri y Molles de Famatina (Floiano, Dapingiano/ Darriwiliano) algunos taxones comunes con la ASZ5: Eupoikilofusa striatogranulata (como Dactylofusa striatogranulata), Veryhachium trispinosum, Eisenackidiumorientalis, Vavrdovella areniga, Dactylofusa velifera forma brevis y Veryhachium lairdii.
Un trabajo reciente de Rubinstein et al. (2008) menciona escasos acritarcos diagnósticos del Ordovícico Superior (Katiano) en Caspalá, aunque ninguno de ellos fue registrado en la ASZ5.
Edad de la asociación. La mayoría de los taxones registrados tienen rangos de distribución entre el Floiano - Dapingiano, como Coryphidium, Cymatiogalea messaoudensis var. messaoudensis, Dactylofusa velifera, Eisenackidium orientalis, Petaloferidium bulliferum y Picostella turgida. Otras formas que se encuentran en el lapso Arenigiano - Llanvirniano (Floiano, Dapingiano, Darriwiliano) son Adorfia hoffmanensis, Aureotesta, Ericantheapollicipes, Vavrdovella areniga y el género Striatotheca El género Arkonia tiene su FAD en la base del Llanvirniano (Darriwiliano), Pterospermella colbathii tiene registros comprobados en el Llanvirniano temprano, Veryhachium rhomboidium es conocido a partir del Caradociano (Sanbdiano) y Leiofusatumida tiene registros a partir del Ashgiliano (Katiano medio - Hirnantiano). Acanthodiacrodium crassus tiene registros confirmados por otros fósiles a partir del Ashgiliano aunque fue descripta en estratos más antiguos (Jachowicz, 1995).
Aunque la ASZ5 tiene en común varias formas con las microfloras de las formaciones Zanjón y Acoite, se observó una mejor correspondencia con la de Formación Capillas, cuya edad máxima es darriwiliana. Sin embargo, la presencia de Acanthodiacrodium crassus, Leiofusa tumida y Veryhachium rhomboidium admitirían considerar edades más jóvenes para la asociación. La presencia del conodonte Erismodus aff. quadridactylus en los niveles más altos del Perfil N° 5 ratifica para los mismos una edad darriwiliana alta/sandbiana baja (op. cit.).
En base a la composición palinológica y las comparaciones con otras asociaciones se asigna para la ASZ5 una edad extendida entre el Darriwiliano hasta el Sanbdiano o Katiano/Hirnantiano.
Observaciones. Una característica destacable de la ASZ5 es la proporción relativamente alta de formas complejas y con procesos como Polygonium, Cymatiogalea, Stellechinatum, Veryhachium y Tectitheca, presentes en cantidad variable en la mayoría de los niveles y que permiten interpretar paleoambientes abiertos con cierta distancia de la costa.
Por otros lado, y como ocurre en las otras asociaciones definidas en Sierra de Zenta, la ocurrencia de formas como Striatotheca y Coryphidium en la ASZ5 permite considerar afinidades paleogeográficas con la provincia fría peri-Gondwana.
Descripciones sistemáticas
LISTA DE PALINOMORFOS
Acritarcos
Acanthodiacrodium angustum (Downie 1958) Combaz 1967 (Fig. 10-A)
Acanthodiacrodium crassus (Loeblich y Tappan 1978) emend. Vecoli 1999 (Fig. 12-I)
Acanthodiacrodium spinum Rasul 1979 (Fig. 10-B)
Acanthodiacrodium ubuii Martin 1969 (Fig. 10-C)
Acanthodiacrodium spp.
Adorfia hoffmanensis (Cramer, Allam, Kanes y Díez 1974) Ottone en Ottone et al.,1992(Fig. 15-I)
cf. Arkonia sp. (Fig. 12-H)
Aureotesta? sp. (Fig. 15-J)
Balthisphaeridium coolibahense Playford y Wicander 1988 (Fig. 11-E)
Balthisphaeridiumfragile Tongiorgi, Yin, Di Milia 1995 (Fig. 13-C)
Baltisphaeridiumomnivagum Playford y Wicander 1988 (Fig. 11-F)
Baltisphaeridium sp. cf. B. crinitum Martin 1978 (Fig. 11-D)
Balthisphaeridium sp. 1 (Fig. 10-D)
Barakellafelix Cramer y Díez 1977 (Fig. 11-G)
Barakellarara (Lu 1987) emend. Tongiorgi, Yin y Di Milia 1995 (Fig. 11-H)
Barakella sp. (Fig. 12-L)
Coryphidiumbohemicum Vavrdova 1972 (Fig. 15-K)
Coryphidium sp. cf. C. elegans Cramer, Allam, Kanes y Diez 1974 (Fig. 15-L)
Grupo CoryphidiummiladaeCoryphidium tadla Rubinstein en Rubinstein et al. 1999 (Fig. 12-A)
Coryphidium spp.
Cristalliniumcambriense (Slavikova) Vanguestaine 1978 (Fig. 10-E)
Cristalliniumdentatum (Vavrdova) Martin, Li Jun 1987 (Fig. 13-K)
Cristallliniumovillense (Cramer y Diez) Martin 1981 (Fig. 10-F)
Cymatiogalea cuvillieri (Deunff) Deunff 1964 (Fig. 10-G)
Cymatiogaleadeunffii Jardine, Combaz, Magloire, Peniguel y Vachey 1974 (Fig. 11-J)
Cymatiogaleagranulata Vavrdova 1966 (Fig. 13-L)
Cymatiogalea membranispina Deunff 1961 (Fig. 10-H)
Cymatiogaleamessaoudensis Jardine, Combaz, Magloire, Peniguel y Vachey 1974 var. messaoudensis (Fig. 14-G)
Cymatiogaleavelifera (Downie) Martin 1969 (Fig. 10-J)
Cymatiogalea spp. (Fig. 14-H)
Dactylofusavelifera Cocchio 1982 emend. Albani 1989 var. brevis sensu Albani 1989 (Fig. 12-B)
Dasydiacrodium spp. (Fig. 12-C)
Eisenackidiumorientalis Rubinstein 1999 en Rubinstein, Toro y Waisfeld 1999 (Fig. 12-D)
Eisenackidium sp. 1 (Fig. 14-B)
Eliasumllaniscum Fombella 1977 (Fig. 10-K)
Ericantheapollicipes Cramer y Diez 1977 (Fig. 12-F)
Eupoikilofusastriatogranulata Vavrdová 1989 (Fig. 14-C)
Eupoikilofusa sp. 1
Eupoikilofusa spp. (Fig. 11-L)
Ferromia pellita (Martin) Martin 1996
Leiofusafusiformis (Eisenack) Eisenack 1938 (Fig. 14-E)
Leiofusa tumida Downie 1959 (Fig. 14-F)
Leiofusa spp. (Fig. 14-D)
Micrhystridium spp.
Multiplicisphaeridium? spp. (Fig. 14-J)
Petaloferidium bulliferum Yin, Di Milia y Tongiorgi 1998
Picostella turgida Yin, Di Milia y Tongiorgi 1998 (Fig. 13-A)
cf. Picostella sp.
Pirea sp. cf. P.colliformis (Burmann) Eisenack, Cramer y Díez 1976 (Fig. 13-B)
Pirea sp. cf. P. ornata (Burmann) Eisenack, Cramer y Díez 1976 (Fig. 14-L)
Poikilofusa sp.
Polygonium dentatum (Timofeev ex Konzalová-Mazancová) Albani 1989
Polygoniumgracile Vavrdova 1966 (Fig. 15-A)
Polygonium spp. (Fig. 13-D)
Priscogalea sp.
Saharidiadownieii Combaz 1967
Saharidiafragilis (Downie 1958) Combaz 1967 (Fig. 10-L)
Stellechinatumuncinatum (Downie) Molyneux, 1987 (Fig. 15-C)
Stellechinatum sp. 1 (Fig. 15-B)
Stelliferidium stelligerum (Górka) Deunff, Górka y Rauscher 1974 (Fig. 15-D)< br /> Stelliferidium striatulum (Vavrdova) Deunff, Górka y Rauscher 1974 (Fig. 15-E)
Stelliferidium spp. (Fig. 13-E)
Striatothecaprincipalis Burmann 1970 (Fig. 13-F)
Striatotheca rugosa Tongiorgi, Yin, Di Milia 1995 (Fig. 12-K)
Striatotheca sp. cf. S. transformata Burmann 1970 (Fig. 11-B)
Striatotheca spp.
Tectithecaadditionalis Burmann 1968 (Fig. 13-G)
Tectithecavalida Burmann 1968 (Fig. 11-I)
Tectitheca sp. cf. T. additionalis
Vavrdovella areniga (Vavrdová) Loeblich y Tappan 1976 (Fig. 13-H)
Veryhachiumlairdii (Deflandre) Deunff 1959 ex Downie 1959 (Fig. 12-E)
Veryhachium minutum Downie 1959
Veryhachiumrhomboidium (Downie 1959) emend. Turner 1984 (Fig. 15-G)
Veryhachiumsymmetricum Lu 1987 emend. Tongiorgi, Yin y Di Milia 1995 (Fig. 13-I)
Veryhachiumtrispinosum (Eisenack 1938) Deunff 1954 ex Downie 1959 (Fig. 15-H)
Vulcanisphaeraafricana Deunff 1961 emend. Rasul 1976 (Fig. 10-I)
Vulcanisphaerabritannica Rasul 1976 (Fig- 11-A)
Vulcanisphaeratuberata (Downie 1958) Eisenack et al., 1973 (Fig. 13-J)
Vulcanisphaeraturbata Martin 1981 (Fig. 11-C)
Fusiformes indeterminados
Algas prasinofíceas
Cymatiosphaera spp. (Fig. 14-I)
Pterospermella colbathii Vavrdová 1990a (Fig. 11-K)
Pterospermella sp. 1 (Fig. 12-G)
Pterospermella spp.
Leiosphaeridia spp.
Quitinozoos
Conochitina sp. (Fig. 15-F)
Euconochitina sp.
Eremochitina sp. (Fig. 12-J)
Lagenochitina sp. (Fig. 14-K)
Criptosporas indeterminadas (Fig. 14-A)
 f
f
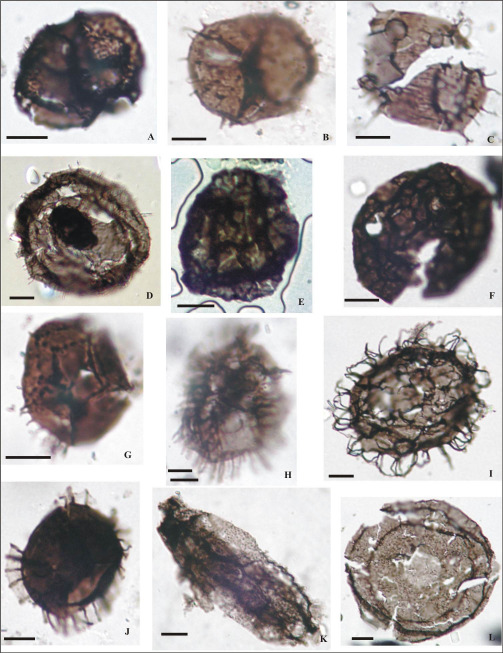
Figura 10. (Escala 10 µm): A-Acanthodiacrodium angustum LIL PB pm 425 (M2-3 55Y); B-Acanthodiacrodium spinum LIL PB pm/ICG 101 (M11-1 53H); C- Acanthodiacrodium ubuii LIL PB pm /ICG 101 (M11-2 51X1); D-Balthisphaeridium sp. 1 LIL PB pm 425 (M2-C 40J); E-Cristallinium cambriense LIL PB pm 425 (M2-A 39U4); F-Cristallinium ovillense LIL PB pm 425 (M2-F 50J3); G- Cymatiogalea cuvillieri LIL PB pm/ICG 101 (M11-5 24T); H-Cymatiogalea membranispina LIL PB pm/ICG 101 (M11-6 46E); I-Vulcanisphaera africana LIL PB pm 425 (M2-C 29H3); J-Cymatiogalea velifera LIL PB pm/ICG 102 (M12-1 41L); K-Eliasum llaniscum LIL PB pm 425 (M2-C 27P2); L-Saharidia fragilis LIL PB pm 425 (M2-F 34Y4).

Figura 11. (Escala 10 µm): A-Vulcanisphaera britannica LIL PB pm 425 (M2-E 38H); B-Striatotheca sp. cf. S. transformata LIL PB pm/ICG 121 (M66-2 49U); C-Vulcanisphaeraturbata LIL PB pm 425 (M2-A20T); D-Balthisphaeridium sp. cf. B. crinitum LIL PB pm 428 (M8-4 30T3); E-Balthisphaeridium coolibahense LIL PB pm/ICG 106 (M18-1 25B); F- Balthisphaeridium omnivagum LIL PB pm/ICG 106 (M18-4 41X4); G-Barakella felix LIL PB pm/ICG 106 (M18-2 22F); H-Barakella rara LIL PB pm/ICG 106 (M18-4 33U); I-Tectitheca valida LIL PB pm/ICG 127 (M86-5 42T); J- Cymatiogalea deunffii LIL PB pm/ICG 110 (M31-6 29C4); K-Pterospermella colbathii LIL PB pm/ICG 126 (M85-1 35Y2); L-Eupoikilofusa sp. LIL PB pm 428 (M18-1 55U3).

Figura 12. (Escala 10 µm; Quitinozoo con escala 100 µm): A-Coryphidium miladae-tadla LIL PB pm/ICG 123 (M68- 3 54M); B-Dactylofusa velifera forma brevis LIL PB pm/ICG 123 (M68-1 37B); C-Dasydiacrodium sp. LILPB pm/ICG 123 (M68-5 47Z); D-Eisenackidiumorientalis LIL PB pm/ICG 127 (M67-4 30X; E-Veryhachium lairdii LIL PB pm/ICG 123 (M68-3 56P); F-Ericanthea pollicipes LIL PB pm/ICG 126 (M85-2 31Y); G-Pterospermella sp. 1 LIL PB pm/ICG 123 (M68-1 56D); H- cf. Arkonia sp. LIL PB pm/ICG 125 (M81-1 53L3); I- Acanthodiacrodium crassus LIL PB pm/ICG 127 (M86-7 38L3); J-Eremochitina sp. LIL PB pm/ICG 123 (M68-4 31P); K-Striatotheca rugosa LIL PB pm/ICG 123 (M68- 3 49D); L-Barakella sp. LIL PB pm/ICG 128 (M87-2 39V2).
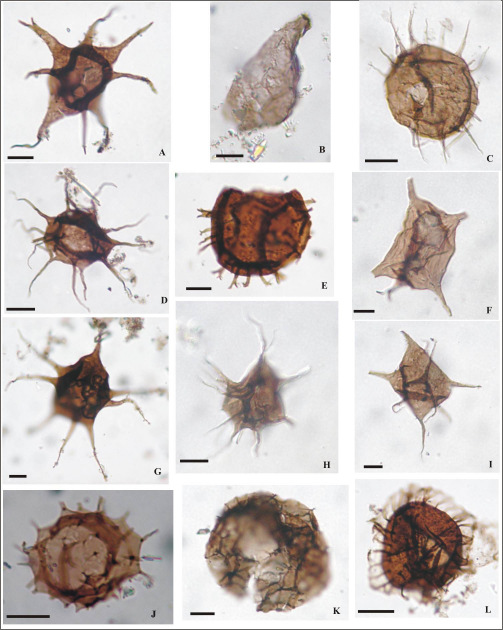
Figura 13. (Escala 10 µm): A-Picostella turgida LIL PB pm/ICG 110 (M31-6 36F); B- Pirea sp. cf. P. colliformis LIL PB pm/ICG 110 (M31-1 57D4); C-Balthisphaeridium fragile LIL PB pm/ICG 110 (M31-6 36E2); D- Polygonium sp. LIL PB pm/ICG 110 (M31-4 50M4); E- Stelliferidium sp. LIL PB pm/ICG 113 (M37-1 54E3); F-Striatotheca principalis LIL PB pm/ICG 113 (M37-1 50S1); G-Tectitheca additionalis LIL PB pm /ICG 123 (M68-3 35H); H-Vavrdovella areniga LIL PB pm/ICG 110 (M31-6 41K2); I- Veryhachium symmetricum LIL PB pm/ICG 110 (M31-6 42L4); J-Vulcanisphaera tuberata LIL PB pm/ICG 113 (M37-136H3); K-Cristallinium dentatum LIL PB pm/ICG 110 (M31-6 50V2); L-Cymatiogalea granulata LIL PB pm/ICG 115 (M51-3 44E4).
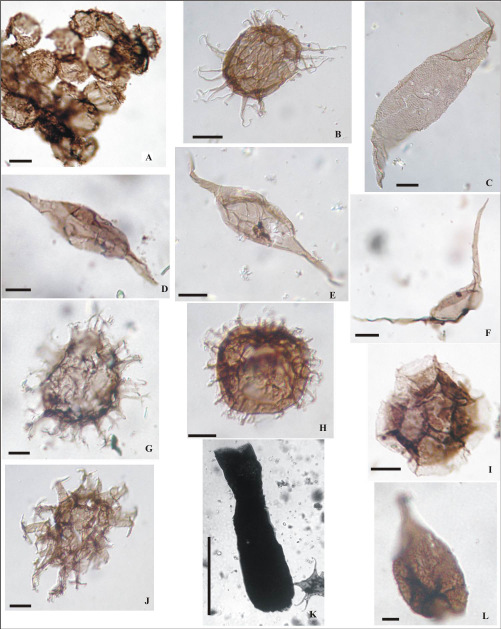
Figura 14. (Escala 10 µm): A- Criptosporas indeterminadas LIL PB pm/ICG 116 (M53-8 46X4). B-Eisenackidium sp. 1 LIL PB pm/ICG 116 (M53-9 55B4); C-Eupoikilofusastriatogranulata LIL PB pm/ICG 116 (M53-9 24V); D- Leiofusa sp. LIL PB pm/ICG 116 (M53-2 47U); E-Leiofusa fusiformis LIL PB pm/ICG 120 (M62-5 43D2); F-Leiofusa tumida LIL PB pm/ICG 120 (M62-5 54J2); G-Cymatiogalea messaoudensis var. messaoudensis LIL PB pm/ICG 120 (M62- 5 49L); H-Cymatiogalea sp. LIL PB pm/ICG 116 (M53-9 33B4; I-Cymatiosphaera sp. 1 LIL PB pm/ICG 116 (M53-8 36Y4); J- Multiplicisphaeridium? sp. LIL PB pm/ICG 116 (M53-9 41C4); K-Lagenochitina sp. LIL PB pm/ICG 123 (M68- 3 44Q); L-Pirea sp. cf. P. ornata LIL PB pm/ICG 120 (M62-5 52X2).

Figura 15. (Escala 10 µm, Quitinozoo: Escala 100 µm): A- Polygonium gracile LIL PB pm/ICG 116 (M53-8 32M2); B- Stellechinatum sp. 1 LIL PB pm/ICG 116 (M53-7 28J); C-Stellechinatum uncinatum LIL PB pm/ICG 116 (M53-9 33B4); D-Stelliferidium stelligerum LIL PB pm /ICG 118 (M56-4 36X); E-Stelliferidium striatulum LIL PB pm/ICG 116 (M53- 5 54T); F-Conochitina sp. LIL PB pm 433 (M13-3 48J1); G-Veryhachium rhomboidium LIL PB pm/ICG 120 (M62-5 36O1); H- Veryhachium trispinosum LIL PB pm/ICG 120 (M62-5 42G); I- Adorfia hoffmanensis LIL PB pm/ICG 123 (M68-1 27Y3); J- Aureotesta? sp. LIL PB pm/ICG 123 (M68-2 57X3); K-Coryphidium bohemicum LIL PB pm/ICG 123 (M68-1 46L3); L-Coryphidium sp. cf. C. elegans LIL PB pm/ICG 123 (M68-5 32Z).
NCERTAE SEDIS
GRUPOACRITARCHAEVITT 1963
GÉNERO ACANTHODIACRODIUM(TIMOFEEV 1958) DEFLANDRE Y DEFLANDRE-RIGAUD 1962
Especie tipo Acanthodiacrodium dentiferumTimofeev 1958
Observaciones. Se sigue el criterio de Tongiorgi (en Bagnoli et al., 1988) y Servais y Molyneux (1997) que Actinotodissus Loeblich y Tappan (1978) es sinónimo junior de Acanthodiacrodium (Timofeev 1958) emend. Deflandre y Deflandre-Rigaud (1962).
- Acanthodiacrodium angustum(Downie 1958) Combaz 1967.
1958 Diornatosphaeraangustum Downie, pp. 345-346, pl. 17, figs. 7, 8, text- fig. 3e.
1967 Acanthodiacrodium angustum (Downie) Combaz, p. 15.
Dimensiones (5 ejemplares): Long. Vesícula: 32-35 ìm; ancho vesícula: 23-28 ìm; cant. de procesos en cada polo: 70-80 ìm. Fig. 10-A.
Registros en Argentina: Cámbrico - Tremadociano: Quebrada de Moya (Aráoz y Vergel, 2006). Tremadociano Inferior: La et al., 2008 b; Aráoz, 2009). Este trabajo: M5 (R), M11 (RR).
- Acanthodiacrodium crassus (Loeblich y Tappan 1978) Vecoli 1999.
1978 Actinotodissus crassus Loeblich y Tappan 1978, pp. 1238, 1241, pl. 1, figs. 5-7.
1999 Acanthodiacrodium crassus (Loeblich y Tappan 1978) Vecoli, p. 28, pl. 1, figs. 8-10.
Dimensiones (15 ejemplares): Long. Vesícula: 21-29 ìm; ancho vesicula: 12-25 ìm; cant. de procesos: 12-16; long. de los procesos: 9-22 ìm. Fig. 12-I.
Registros con control independiente. Ashgiliano: Zona Dicellograptus complanatus var. ornatus Canadá (Jacobson y Achab, 1985). En Argentina: Tremadociano:Sierra de Zenta (Aceñolaza et al., 2003) y posteriormente asignado a Acanthodiacrodium sp. (Aráoz, 2009). Este trabajo: M53 (R), M67 (RR), M86(C),M87 (R).
- Acanthodiacrodium ubuii Martin 1969.
1969 Acanthodiacrodium ubuii Martin, pp. 127-128; pl. 1, fig. 51; text-fig. 81.
Dimensiones (3 ejemplares): Ancho vesícula: 26-28 ìm; long. Vesícula: 30 ìm; long. de los procesos: 4-6 ìm. Fig. 10-C.
Registros con control independiente. Registro más bajo: Tremadociano Inferior: en niveles de Dictyonema flabelliforme, Inglaterra (Rasul y Downie, 1974). Registro más alto: Tremadociano Superior, Zona Paltodus deltifer, Suecia (Bagnoli et al., 1988). En Argentina: Tremadociano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M12 (RR).
- Acanthodiacrodium spp.
Dimensiones (35 ejemplares): Long. Vesícula: 22-30 µm; ancho vesícula: 15-20 µm; procesos: 5-7 en cada polo; long. de los procesos: 9-25 ìm.
Descripción. Vesículas bipolares, con un estrechamiento en el sector ecuatorial más o menos marcado y contorno aproximadamente rectangular. En cada polo se levanta un número variable de procesos simples, huecos, homomórficos, cónicos a cilíndricos, acuminados y comunicados con la cavidad de la vesícula. Las bases de los procesos están ensanchadas y son ligeramente cónicas, dando un contorno poligonal a los polos. Algunos procesos pueden surgir en la parte media de la vesícula. La pared de la vesícula y de los procesos es delgada y de superficie lisa.
Observaciones. EnAcanthodiacrodium spp. se agruparon especimenes relativamente similares con una alta variabilidad en tamaño, forma y número y distribución de procesos. Se destaca la presencia, en diferentes niveles de ejemplares con morfologías intermedias entre Acanthodiacrodium, olygoniun, Tectitheca, Vavrdovella y Dasydiacrodium.
Registros en Argentina: Tremadociano: Sierra de Zenta (Aráoz, 2009). Este trabajo. M2(C), M14(R), M31 (RR), M69 (C), M85(C).
GÉNERO ADORFIABURMANN 1970
Especie tipo Adorfia firma Burmann 1970
Observaciones: se sigue el criterio deMolyneux (1987); Molyneux y Rushton (1988); Ottone et al. (1992) y Tongiorgi et al. (2003) que consideran que Adorfia y Multiplicisphaeridium son géneros separados.
- Adorfia hoffmanensis (Cramer, Allam, Kanes y Díez) Ottone en Ottone et al. 1992.
1974 Multiplicisphaeridium hoffmanensis Cramer et al., p. 185, pl. 27, fig.10
1992 Adorfia hoffmanensis (Cramer et al.) - Ottone en Ottone et al., pp. 96-97, pl.1, figs. 2, 4, 6, 10.
Dimensiones (5 ejemplares): Diámetro vesícula: 25-30 µm; cant. de procesos: 10-15; long. de los procesos 10-15 µm; ancho base procesos: 8-10 µm. Fig. 15-I.
Observaciones. La descripción del ejemplar corresponde a la de Ottone et al. (1992) aunque tiene el ancho de las ramificaciones ligeramente mayor.
Registro con control independiente. Registro más bajo: Yushanian medio, Zona de A. suecicus, Sur de China (Li, 1987). En Argentina: Arenigiano: Sierra de los Colorados (Ottone et al.,1992); Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Quebrada Grande y Quebrada de la Curva (Rubinstein y Toro, 1999; 2001).Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo: M68 (R).
GÉNERO ARKONIA BURMANN 1970
Especie tipo Arkonia virgata Burmann 1970
- cf. Arkonia sp.
1970 Arkonia virgata Burmann, p. 298, pl. 7, fig. 6.
Dimensiones (1 ejemplar):Long. Lados: 20 µm;long. de los procesos: 12-22 µm; ancho base procesos: 3 µm. Fig. 12-H.
Descripción. Cuerpo central triangular, comprimido, con los lados rectos a ligeramente cóncavos y un proceso que emerge de cada esquina. Los procesos son cónicos y gruesos, con bases anchas, casi sin modificación de la forma y con punta redondeada. En la superficie de la vesícula se observan finas estrías paralelas a los lados que se extienden a los procesos.
Observaciones. La mala preservación del ejemplar no permite una buena observación de las estrías por lo que se asigna como cf. Arkonia sp.
Registros en Argentina: Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo. M81 (RR).
GÉNERO AUREOTESTA (VAVRDOVÁ) BROCKE, FATKA Y SERVAIS 1998
Especie tipo Aureotesta clathrata Vavrdová 1972
- Aureotesta sp.
1998 Aureotesta clathrata (Vavrdová 1972) - Brocke, Fatka y Servais, pp .5-8, pl. 1, figs. A-M; pl..
2, figs. A-K.
Dimensiones (30 ejemplares): Long. Vesícula: 25-32 µm; ancho vesícula: 18-28 µm; long. de los procesos: 8-25 µm; base procesos: 2-4 µm; ancho red: 3-6 µm. Fig. 15-J.
Descripción. Vesícula de cuerpo central aplanado, de pared delgada y contorno cuadrado, rectangular, trapezoidal o pentagonal, con los lados rectos a ligeramente convexos. La vesícula sostiene entre cuatro y siete procesos largos, cónicos, de bases anchas y con los extremos afilados. Los procesos son huecos, sin comunicación con el interior de la vesícula por la presencia de tapones en la base de los mismos. La distribución de los procesos es irregular, generalmente desde los ángulos y condiciona la forma de la vesícula. Los procesos sostienen una ornamentación formada por numerosas excrescencias como pelos, irregularmente ramificados y conectados entre sí, formando una malla que rodea los procesos y que puede extenderse en los sectores cercanos a las bases. El tamaño de la red filamentosa es variable, generalmente entre 3-6 µm a cada lado de los procesos y disminuye hacia la parte terminal de los mismos. La superficie de la vesícula es ligeramente granulada y en algunos casos con estrías paralelas a los lados.
Observaciones.Aureotesta? sp. Tiene una morfología similar a A. clathrata var. clathrata, principalmente por el retículo de filamentos alrededor de los procesos y sobre la vesicula. Sin embargo, marcadas diferencias en otros caracteres y su variabilidad, la separan tanto de la especie como del género, como la forma del cuerpo central, el número de procesos, la presencia de tapones en las bases de los mismos, estrías en el cuerpo.
Registros previos seleccionados. Aureotesta aparece en el Arenigiano (Brocke, 1992; Vavrdová, 1993) hasta el Llanvirniano temprano (Servais y Maletz, 1992; Servais, 1993). En Argentina A. clathrata var. clathrata no fue registrada, pero si A. clathrata var. simplex. (Rubinstein et al., 2007 b).
En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M68 (CC).
GÉNERO BALTISPHAERIDIUM EISENACK 1958 EX EISENACK 1959 EMEND. STAPLIN, JANSONIUS Y POPOCK 1965 EMEND. EISENACK 1969
Especie tipo Baltisphaeridium longispinosum (Eisenack) Eisenack 1959
- Balthisphaeridium coolibahense Playford y Wicander 1988
1988 Balthisphaeridium coolibahensePlayford y Wicander. p. 8, figs. 3 A-F.
Dimensiones (2 ejemplares): Diámetro vesícula: 45x47 µm. Fig. 11-E.
Registros con control independiente. Registro más bajo: Yushanian Medio, Zona A. suecicus, Sur de China (Yin, 1995). Registro más alto: Arenigiano medio a superior (Fortey y Shergold, 1984). En Argentina: Arenigiano: Famatina (Achab et al., 2006). Dapingiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M18 (RR).
- Baltisphaeridium fragile Tongiorgi, Yin, Di Milia 1995.
1995 Baltisphaeridiumfragile Tongiorgi, Yin, Di Milia, pp. 23-24, pl. 1, figs. 3, 8.
Dimensiones (1 ejemplar): Diámetro vesícula: 27-29 µm; long. de los procesos: 10-14 µm. Fig. 13-C.
Observaciones. La descripción corresponde a la diagnosis original aunque el diámetro de la vesícula y la longitud de los procesos son menores.
Registros previos seleccionados. Arenigiano: Sur de China (Tongiorgi et al., 1995; Tongiorgi et al., 2003). Ashgilliano tardío: Suecia (Le Hérissé, 1988). En Argentina: Floiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR).
- Baltisphaeridium omnivagum Playford y Wicander 1988.
1988 BaltisphaeridiumomnivagumPlayford y Wicander. pp.8-10, figs. 3 G -L.
Dimensiones (15 ejemplares): Long. Vesícula: 37-43 µm; ancho vesícula: 29-33µm; cant. de procesos: 40-45; long. de los procesos: 7-15µm; base procesos: 0,5-1 µm. Fig. 11-F.
Descripción. Vesícula originalmente esférica, de contorno circular a subcircular, con un número variable de procesos homomórficos, simples, elongados y delgados como espinas. Procesos rectos a ligeramente curvados, aparentemente sólidos y no comunicados con el interior de la vesícula. La pared vesicular es delgada y lisa y deformada por pliegues de compresión periféricos.
Registros previos. Arenigiano: Australia (Playford y Wicander, 1988). En Argentina: Arenigiano: Famatina, como B. cf. B. omnivagum (Achab et al., 2006). Dapingiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M18 (CC).
- Baltisphaeridium sp. cf. B. crinitumMartin 1978 en Dean y Martin 1978.
1978 Balthisphaeridium crinitum Martin, sp. nov., en Dean y Martin, p. 7, pl. 3, fig. 5.
Dimensiones (1 ejemplar): Diámetro vesícula: 35x37 µm; cant. de procesos: 90 aprox.; long. de los procesos: 1-3 µm; ancho base procesos: 1 µm. Fig. 11-D.
Descripción. Vesícula de contorno circular a subcircular y pared lisa, con los procesos distribuidos irregularmente en la superficie de manera irregular, con una separación de 4 a 8 µm. Los procesos son homomórficos, simples, acuminados y filosos en su extremo distal, con una fina granulación, huecos y no comunicados con el interior de la vesícula. No se observa la abertura de enquistamiento.
Observaciones. La fina granulación en los procesos podrían ser restos de los pelos descriptos para la especie, además el rango de tamaños es menor al de la diagnosis por lo que el espécimen se describe como B. sp. cf. B. crinitum.
Registros en Argentina: Dapingiano:Sierra de Zenta (Aráoz, 2009). Este trabajo. M18 (R).
- Baltisphaeridium sp. 1
Dimensiones (9 ejemplares): Diámetro vesícula: 35-48µm; long. de los procesos: 2-4 µm. Fig. 10-D.
Descripción. Vesícula de contorno circular a subcircular, de pared delgada y superficie lisa con varios pliegues concéntricos en la periferia. Alrededor de 60 a 80 procesos homomórficos se distribuyen de manera irregular sobre toda la superficie. Estos son simples, alargados, acuminados y filosos en el extremo distal, además son huecos y sin comunicación con el interior de la vesícula. Los ejemplares asignados a Balthisphaeridium sp. 1 presentan una característica estructura irregular de color oscuro sobre la superficie.
Observaciones.Balthisphaeridium sp. 1 es comparable con B. crinitum pero no se observa la ornamentación de pelos en los procesos. Balthisphaeridium sp. 1 tiene una morfología similar a B. omnivagum pero se diferencia de este por la presencia de la estructura irregular de la superficie que es constante en todos los ejemplares y cuya naturaleza es desconocida aunque podría ser preservacional o por causas paleoambientales.
Registros en Argentina: Tremadociano Inferior:Sierra de Zenta (Aráoz, 2009). Este trabajo. M2 (C).
GÉNERO BARAKELLA CRAMER Y DÍEZ 1977
Especie tipo Barakella fortunata Cramer y Díez 1977
- Barakella felix Cramer y Díez 1977.
1977 Barakellafelix Cramer y Díez, pp. 345, pl. 5, figs. 1, 2, 11; text-figure 3:22.
Dimensiones (1 ejemplar): Long. Vesícula: 36 µm; ancho vesícula: 25 µm; cant. de los procesos en polo antiapical: 14; cant. de procesos en polo apical: 10; long. de los procesos principales: 5-16 µm; long. de los procesos secundarios: 4-7 µm. Fig. 11-G.
Descripción. Vesícula subrectangular con procesos primarios delgados y largos dispuestos en diferente número en cada uno de los polos diferenciados o en los ángulos del cuerpo. La porción antiapical de la vesícula se diferencia por la presencia de un parche de elementos filamentosos, anastomosados e interconectados periféricamente. Algunos procesos accesorios similares a los primarios pero más pequeños se observan en las áreas polares. En la parte central de la vesícula se diferencian fuertes pliegues diacrodianos.
Registros previos seleccionados. Arenigiano tardío: Marruecos (Cramer y Díez, 1977). Barakella felix tiene su FAD en la base del Dapingiano (Li et al., 2002) y Servais et al. (2008) lo hace extensivo a todo el género. En Argentina: Floiano tardío de Cordillera Oriental, como Barakella Rubinstein et al. (2007 b). Dapingiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M18 (RR).
- Barakella rara (Lu) Tongiorgi, Yin y Di Milia 1995
1987 Actinotodissus rarus Lu, p. 89, pl. 1, figs. 1, 2, 10.
1995 Barakella rara (Lu) emend. Tongiorgi et al., pp. 24-26, pl. 1, figs. 9,10.
Dimensiones (1 ejemplar): Long. Vesícula: 30 µm; ancho vesícula: 24 µm. Fig. 11-H.
Observaciones. El ejemplar observado tiene un tamaño menor al de la diagnosis original.
Registros con control independiente. Registro más bajo: Arenigiano temprano: Zona Azygograptus suecicus y Zona Oepikodus evae, Sur de China (Li, 1987). Registro más alto: Llanvirniano temprano, Zona D. murchisoni, Túnez (Vecoli, 1999). En Argentina: Arenigiano medio: Área de Santa Victoria, como B. cf. B. rara, Zona de Didymograptus bifidus (Rubinstein et al., 1999; Rubinstein y Toro, 2001), que Rubinstein et al. (2007 b) consideran equivalente a la Biozona de D. bifidus del margen occidental de la Cordillera Oriental del Floiano tardío. Dapingiano: Sierra de Zenta (Aráoz, 2009). Darriwiliano: Sierra de Zapla, como B. cf. B. rara (Rubinstein y de la Puente, 2008).Este trabajo: M18 (RR).
- Barakella sp.
Dimensiones (1 ejemplar):Long. Vesícula: 30 µm; ancho vesícula: 25 µm. Fig. 12-L.
Descripción. Vesícula de forma subrectangular, con dos polos diferenciados y ligeramente redondeados que sostienen procesos heteromórficos en cada uno. Los procesos de primer orden miden 10-12 µm de longitud, son ahusados con terminaciones distales acuminadas, huecos y comunicados con la cavidad de la vesícula. En el polo apical hay 7 procesos y en el polo antiapical 10 procesos. Los procesos de segundo orden son pequeños, de 5-6 µm y están entre los procesos principales. En el sector central de la vesícula se observan pliegues longitudinales tipo diacrodianos.
Observaciones. Se diferencia de B. rara porque ésta tiene un menor número de procesos más gruesos y una forma más rectangular; y de B. felix porque ésta tiene una vesícula rectangular y los procesos son más cónicos y cortos.
Registro en Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo. M87 (RR).
GÉNERO CORYPHIDIUM (VAVRDOVÁ) SERVAIS, LI, MOLYNEUX Y VECOLI 2008
Especie tipo Coryphidium bohemicum Vavrdová 1972
- Coryphidium sp. cf. C. elegans Cramer, Allan, Kanes y Díez 1974
1974 Coryphidium elegans Cramer, Allan, Kanes y Díez, p. 184, pl. 27, figs. 12-19.
Dimensiones (5 ejemplares):Long. lados: 21-30 µm; cant. de procesos: 40-60; long. de los procesos: 2-3 ìm. Fig. 15-L.
Descripción. Vesícula de cuerpo central hueco y contorno cuadrangular, con ángulos redondeados y lados rectos a convexos. La superficie tiene ornamentación de finas crestas o estrías bajas, subparalelas a los lados de la vesícula y más abundantes en el sector central. La vesícula sostiene numerosos procesos con distribución irregular y mayor concentración en los ángulos. Los procesos son cortos y de aspecto sólido, delgados y flexibles como espinas o pelos, con las bases ligeramente más anchas y terminaciones simples acuminadas o bulbosas.
Observaciones. Diferencias en la forma de las estrías y la disposición de los procesos no permiten una asignación segura con la especie. C. elegans difiere de C. bohemicum en que éste tiene los procesos digitados, de aspecto romo y no filosos.
Registros con control independiente.Coryphidium elegans se registró en Zona D. bulla, Marruecos (Soufiane y Achab, 1993) y Zona de A. suecicus, Sur de China (Brocke et al., 2000). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M61 (R), M68 (C), M69 (RR).
GRUPO CORYPHIDIUM MILADAE - CORYPHIDIUM TADLA
1976 Coryphidium miladum Cramer y Díez, p. 205, pl. 23, figs. 9, 14, 22, 23; fig-text. 2, 6.
1976 Coryphidium tadlum Cramer y Díez, p. 206, pl. 23, figs. 3, 4, 15, 18, 19; fig-text. 2, 9.
1999 Grupo Coryphidium miladae-Coryphidium tadla Rubinstein en Rubinstein et al., pl.3, fig.3,7; pl.5, fig. 4.
Dimensiones (14 ejemplares):Long. lados: 24-30µm. Fig. 12-A.
Registros en Argentina: Arenigiano medio: Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Quebrada Grande (Rubinstein y Toro, 1999). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M56 (R), M68 (CC).
GÉNERO CRISTALLINIUM VANGUESTAINE 1978
Especie tipo Cristallinium cambriense (Slavíková) Vanguestaine 1978
Cristallinium cambriense (Slavíkovà) Vanguestaine 1978
1968 Dictyotidium cambriense Slavíkovà, p. 201, pl. 2, figs. 1,3.
1972 Cymatiosphaera ovillensis Cramer y Diez, p. 44, pl. 2, figs. 4,7,10.
1972 Dictyotidium cambriense Martin, p. 42, pl. 2, fig. 13, pl. 6, fig. 4.
1977 Dictyotidium cambriense Martin, pl. 4, fig. 12, text-fig. 13.
1978 Cristallinium cambriense (Slavíková) Vanguestaine, pp. 270-271, pl. 2, figs. 16,17, pl. 3, figs. 16, 26.
Dimensiones (2 ejemplares): Diámetro vesícula: 30-35 ìm.Número de campos: 15-20. Fig. 10-E.
Observaciones. La mala preservación de la vesícula no permite una buena observación y recuento de los campos en la superficie de la vesícula.
Registros con control independiente. Registro más bajo: Cámbrico temprano, Zona Proampyx linnarssoni (como C. cf. cambriense) Västergötland (Moczydlowska y Vidal, 1986). Cámbrico medio: Zona Ptychagnostusatavus-Tomagnostus fissus, Terranova (Martin y Dean, 1981). Registro más alto: Tremadociano - Arenigiano: Gales (Molyneux y Dorning, 1989). En Argentina: Tremadociano Inferior: Nazareno (Rubinstein, 1997); Sierra de Zenta (Aceñolaza et al., 2003; Aráoz, 2009). Este trabajo. M2 (RR), M3 (RR).
- Cristallinium dentatum (Vavrdová) Martin 1982
1976 Staplinia dentatum Vavrdová, p. 58, pl., figs. 5,6, text-fig. 5.
1978 ?Dictyotidiumdentatum (Vavrdová) Martin, en Dean y Martin, p. 8, pl., 3, figs. 14, 28.
1982 Cristallinium dentatum (Vavrdová) Martin, p. 36.
1990 Cristallinium dentatum (Vavrdová 1976) Martin 1982 Fensome et al., p. 161.
Dimensiones (4 ejemplares):Diámetro vesícula: 30-55 ìm; alto procesos: 1-2 ìm. Fig. 13-K.
Registros previos seleccionados. Arenigiano: Checoslovaquia (Vavrdová, 1976); Sudoeste de China (Li, 1987); Pakistán (Tongiorgi et al., 1994); Terranova (Martin, 1982). En Argentina: Arenigiano: como C. cf. C. dentatum, Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Qda Grande (Rubinstein y Toro, 1999). Floiano/Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (R), M53 (R).
- Cristallinium ovillense (Cramer y Diez) Martin 1981
1972 Zonosphaeridium ovillensis Cramer y Diez, p. 44, pl. 2, figs. 5, 8, 11.
1981 Cristallinium ovillense (Cramer y Diez de Cramer) Martin en Martin y Dean, pp. 17-18; pl. 3. fig. 16.
Dimensiones (1 ejemplar):Diámetro vesícula: 26x31 ìm. Fig. 10-F.
Registros con control independiente. Registro más bajo: Cámbrico Medio, zona Paradoxites paradoxissimus, Noruega (Welsch, 1986). Registro más alto: Cámbrico Superior, Zonas de Olenus y Parabolina spinulosa, Terranova (Martin y Dean, 1981). En Argentina: Cámbrico Superior, Quebrada de Moya (Aráoz y Vergel, 2006). Tremadociano Inferior: Sierra de Zenta (Aráoz, 2009). Este trabajo: M2 (RR).
GÉNERO CYMATIOGALEA (DEUNFF) DEUNFF 1964
Especie tipo Cymatiogalea margaritata Deunff 1961
- Cymatiogalea cuvillieri (Deunff) Deunff 1964
1961 Priscogaleacuvillieri Deunff, p. 41, pl. 1, fig. 2.
1964 Cymatiogaleacuvillieri (Deunff) Deunff, p. 124, pl. 1, figs. 2, 3.
Dimensiones (3 ejemplares):Diámetro vesícula: 20-30 ìm. Fig. 10-G.
Registros con control independiente. Registro más bajo: Cámbrico Superior: Zona Parabolina spinulosa, Rusia (Volkova, 1990). Registro más alto: Tremadociano tardío, Zona Paltodus deltifer, Suecia (Tongiorgi en Bagnoli et al., 1988). En Argentina: Tremadociano: Sierra de Zenta (Aráoz, 2009); Noroeste Argentino (Martin, 1982; Bultynck y Martin, 1982); Nazareno, como C. sp. cf. C. cuvillieri (Rubinstein, 1997). Arenigiano: La Huerta y Río Acoite (Rubinstein y Toro, 1999; 2001). Este trabajo: M11 (R).
- Cymatiogalea deunffii Jardiné, Combaz, Magloire, Peniguel y Vachey 1974
1974 Cymatiogalea deunffi n. sp. Jardiné et al., pp. 123, pl. 1, figs. 7,8.
Dimensiones (2 ejemplares):Diámetro vesícula: 23-26 µm; ancho membrana: 6-10 ìm. Fig. 11-J.
Descripción. Vesícula originalmente esférica, de pared delgada y superficie lisa a microgranulada, con un gran piloma o apertura poligonal a subcircular bordeada por una cresta. La vesícula está rodeada por una membrana delgada y transparente, que tiene los bordes irregulares, ligeramente arqueados que delimitan campos poligonales de la superficie. No se observó el opérculo.
Registros previos seleccionados. Tremadociano:Bohemia (Fatka, 1992, 1993); España (Mette, 1989); Inglaterra y Gales (Molyneux y Dorning, 1989; Cooper y Molyneux, 1990; Millward y Molyneux, 1992). Arenigiano: Pakistán (Tongiorgi et al., 1994). En Argentina: Tremadociano Tardío: Región Pascha - Incamayo (Rubinstein y Toro, 2002; de la Puente y Rubinstein, 2009); Cerro San Bernardo (Rubinstein et al., 2007b). Floiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR).
- Cymatiogalea granulata Vavrdová 1966
1966 Cymatiogalea granulata Vavrdová, p. 412, pl. 3, fig. 2.
Dimensiones (15 ejemplares): Diámetro vesícula: 18-28 x 25-35 ìm; long. de procesos: 8-12 ìm. Fig. 13-L.
Observaciones. La membrana delgada muchas veces se preserva de manera fragmentaria y las ramificaciones distales de los procesos no confirman la asignación a la especie.
Registros en Argentina: Arenigiano: Los Colorados y Río Cajas (Ottone et al., 1992); Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999; Rubinstein y Toro, 2001); Qda. Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001). Floiano: Sierra de Zapla (Rubinstein, 2005; Rubinstein et al., 2007 b; Rubinstein y de la Puente, 2008). Floiano/Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M14 (RR), M31 (RR), M51 (R), M53 (R), M68 (C), M81 (RR), M85 (RR), M86 (RR).
- Cymatiogalea messaoudensis Jardiné, Combaz, Magloire, Peniguel y Vachey 1974 var.
messaoudensis
1974 Cymatiogalea messaoudii Jardiné Combaz, Magloire, Peniguel y Vachey al., pp. 123, pl. 2, figs. 5,6.
1997 Cymatiogalea messoudensis Jardiné et al. var. messaoudensis Servais y Molyneux, pp. 126-127, pl. 2, figs. 14-15, pl. 3, figs. 9-16, pl, 4, figs. 1-7.
Dimensiones (2 ejemplares): Diámetro vesícula: 18x45 µm; long. de los procesos: 5-20 ìm. Fig. 14-G.
Descripción. Vesícula de forma originalmente esférica que tiene una gran apertura circular y la superficie dividida en campos poligonales definidos por el alineamiento de los procesos. Estos son numerosos, probablemente alrededor de treinta, con bases muy anchas que se fusionan y extienden formando una membrana. El extremo distal de los procesos es complejo, con más de tres órdenes de ramificaciones. La superficie de la vesícula puede ser lisa o microgranulada y la membrana generalmente transparente y lisa.
Observaciones. La delineación de los campos poligonales no siempre aparece clara mientras que las bases de los procesos se unen en las bases con una membrana poco desarrollada, similar a las descriptas por Servais y Molyneux (1997).
Registros previos seleccionados. Tremadociano: Sahara (Jardiné et al., 1974). Vavrdová (1982, 1988; 1989; 1990b) la cita en asociaciones del Arenigiano tardío y Ordovícico tardío de Bohemia probablemente debido a retrabajo. En Argentina: Tremadociano Tardío: Región de Pascha (de la Puente y Rubinstein, 2009); Cerro San Bernardo (Rubinstein et al., 2007b). Arenigiano: Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999; 2001); Qda Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001). Floiano: Sierra de Zapla (Rubinstein, 2005; Rubinstein et al., 2007b; Rubinstein y de la Puente, 2008). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M53 (RR), M62 (RR).
GÉNERO DACTYLOFUSA (BRITO Y SANTOS) EMEND. COMBAZ 1967 EMEND. CRAMER 1970
Especie tipo Dactylofusa maranhensis Brito y Santos 1965
Observaciones. Se sigue a Dorning (1994) que diferencia los géneros Dactylofusa (Brito y Santos 1965) Cramer 1970, Eupoikilofusa Cramer 1970; Leiofusa (Eisenack 1938) Cramer 1970 y Poikilofusa Staplin, Jansonius y Popock 1965.
- Dactylofusa velifera (Cocchio) forma brevis Albani 1989
1982 Dactylofusa velifera Cocchio, pp. 32-33, pl. 1, figs. 20-21, text-fig. 3.
1989 Eupoikilofusa velifera (Cocchio) - Albani, pp. 18-19, pl. 2, figs. 12-19.
Dimensiones (380 ejemplares): Long. Vesícula: 25-47 ìm; ancho vesícula: 13-22 ìm; ancho membrana: 3-9 ìm. Fig. 12-B.
Observaciones. Se remarca la amplia variabilidad morfológica de este taxón en los niveles de Sierra de Zenta, que se manifiesta en el rango de tamaño, la forma general de la vesícula y su proporcion de longitud y ancho, los velos y la presencia de las espinas en los polos que son muy raras. No se observó diferencias en la distribución de estas formas a lo largo de los perfiles.
Registros con control independiente. Registro más bajo: Arenigiano más temprano, Zona Tetragraptus akzharensis, Santa Victoria (Rubinstein et al., 1999). Registro más alto: Zhejiangian temprano (Darriwiliano temprano), Zona U. autrodentatus, Sur de China (Brocke et al., 2000); Tremadociano/Floiano/Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M13 (R), M15 (R), M31 (R), M37 (R), M51 (R), M53 (CC), M56 (CC), M62 (CC), M66 (CC), M67 (CC), M68 (AA), M69 (CC), M81 (R), M86 (CC).
GÉNERO DASYDIACRODIUM (TIMOFEEV 1959) EX DEFLANDRE Y DEFLANDRE-RIGAUD 1962, EMEND. MOCZYDLOWSKA Y STOCKFORS 2004
Especie tipo Dasydiacroidium eichwaldii Timofeev 1959
- Dasydiacrodium spp.
Dimensiones (43 ejemplares): Long. vesícula: 18-26 µm; ancho vesícula: 13-22 µm; long. de los procesos: 6-23 ìm. Fig. 12-C.
Descripción. Vesícula de contorno poligonal, bipolar, con los polos separados por una zona ecuatorial angosta y sin procesos. La vesícula tiene la pared delgada y la superficie lisa a finamente granulada, los procesos son lisos. Cada polo sostiene de 4 a 8 procesos homomórficos, simples, delgados y acuminados, huecos y comunicados con la cavidad interna de la vesícula, con las bases ensanchadas y cónicas que modifican el contorno de los polos. Puede haber procesos de menor tamaño intercalados entre los otros.
Registros en Argentina: Tremadociano/ Floiano/ Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo. M2 (C), M31 (R), M53 (R), M56 (C), M62 (RR), M67 (RR), M68 (R), M69 (R), M85 (RR).
GÉNERO EISENACKIDIUM (CRAMER Y DÍEZ 1968) EISENACK, CRAMER Y DÍEZ 1973
Especie tipo Eisenackidium triplodermun (Cramer) Cramer y Diez 1968
- Eisenackidium orientalis Rubinstein en Rubinstein et al. 1999
1992 Eisenackidium sp. A Ottone en Ottone, Toro y Waisfeld, p. 104; pl.3, figs. 6,11.
1999 Eisenackidium orientalis (Rubinstein 1999) p. 280-282, pl. 4, figs. 1,8-10, pl. 5, figs. 7,10, pl. 6, figs. 1-3
Dimensiones (132 ejemplares): Long. Vesícula: 25-40 µm; ancho vesícula: 18-30 µm; long. de los procesos: 5-28 µm; µm. Fig. 12-D.
Observaciones. Rubinstein en Rubinstein et al. (1999) remarca una considerable variabilidad de formas en el Arenigiano. E. orientalis es una forma endémica para el Noroeste Argentino, típicamente arenigiana aunque no se descarta para la misma una edad tremadociana superior (de la Puente et al., 2009).
Registros con control independiente. Arenigiano temprano: Río la Huerta, Zona Baltograptus deflexus, con dudas y Zona Tetragraptus akzharensis (Rubinstein et al., 1999). Arenigiano Medio: Río la Huerta, Zona Didymograptellusbifidus (Rubinstein et al., 1999; Rubinstein y Toro, 2001). En Argentina: Arenigiano: Los Colorados y Río Cajas, como Eisenackidium sp. 1 (Ottone et al., 1992); Secciones Río Acoite y Parcha (Rubinstein et al., 1999; Rubinstein y Toro, 2001); Qda. Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001); Famatina (Achab et al., 2006). Floiano: Sierra de Zapla (Rubinstein, 2005; Rubinstein y de la Puente, 2008); Sierra de Zenta (Aráoz, 2009). Darriwiliano: Sierra de Zenta (Vergel et al., 2008 b; Aráoz, 2009). Este trabajo: M13 (RR), M15 (R), M17 (R), M31 (R), M51 (CC), M53 (CC), M56 (CC), M61 (R), M62 (C), M66 (C), M67 (CC), M68 (CC), M69 (CC), M81 (CC), M85 (C), M86 (C).
- Eisenackidium sp. 1
Dimensiones (78 ejemplares): Long. Vesícula: 20-34 µm; ancho vesícula: 20-28 µm; long. de los procesos: 6-18 µm; ancho µm. Fig. 14-B.
Descripción. Vesícula de contorno circular a subcircular, a veces ligeramente alargadas, con la pared de doble capa. La pared interna más gruesa y aparentemente lisa forma el cuerpo interno que está rodeado por la pared externa más delgada, transparente y plegada que se extiende formando los procesos. Se diferencian entre 8 y 15 procesos simples en cada polo, con una distribución bipolar, homomórficos, relativamente delgados o con engrosamientos irregulares y terminaciones distales afiladas o truncas.
Observaciones.Eisenackidium sp. 1 se diferencia de Eisenackidium orientalis en que la primera tiene rangos de longitudes de vesícula un poco menores, un mayor número de procesos similares entre si, y un rango de longitudes y anchos menor. E. orientalis incluye formas con procesos delgados y procesos adicionales pero Eisenackidium sp. 1 igualmente muestra un mayor número de procesos. Se destaca la amplia variabilidad de formas incluidas en Eisenackidium sp. 1 y no se descarta que sea una variacion de E. orientalis aunque se la mantiene separada hasta estudios más amplios.
Registros en Argentina: Floiano/ Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo. M15 (R), M31 (C), M51 (C), M53 (C), M55 (R), M56 (R), M62 (C), M66 (C), M67 (C), M68 (CC), M69 (C), M81 (R), M86 (R).
GÉNERO ERICANTHEA CRAMER Y DIEZ 1977
Especie tipo Ericanthea pollicipes Cramer y Diez 1977
- Ericanthea pollicipes Cramer y Diez 1977
1977 Ericanthea pollicipes Cramer y Díez, p. 346; pl. 4, figs. 1-3
Dimensiones (8 ejemplares): Diámetro de una célula: 13-16 µm; diámetro total del cenobio: 40-43 µm. Fig. 12-F.
Descripción. Acritarcos cenobiales que consisten en siete células, cinco ubicadas en un mismo plano y las otras dos en el centro, a cada uno de los lados del grupo de cinco. Estas células tienen forma de urna con los bordes lisos y las aberturas orientadas radialmente. Se observan estrías longitudinales irregulares en la pared de las células casi imperceptibles.
Registros en Argentina: Llanvirniano: Sierra de Zapla (Rubinstein, 2005; Rubinstein y de la Puente, 2008). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M53 (RR), M62 (RR), M85 (R), M86 (RR).
GÉNERO EUPOIKILOFUSA CRAMER 1970
Especie tipo Eupoikilofusa stratifera typica Cramer y Díez 1972
- Eupoikilofusa striatogranulata Vavrdová 1989
1974 Dactylofusa striatogranulata Jardiné et al., p. 128; pl. 2, fig. 1.
1974 Dactylofusa striatogranulata Jardiné et al., p. 137.
1984 Poikilofusa striatogranulata (Jardiné et al.) Playford y Martin, pp. 208-209, figs. 8K-L.
1989 Eupoikilofusa striatogranulata Vavrdová, p. 405, text-fig. 1a.5.
Dimensiones (15 ejemplares): Long. Vesícula: 65-115 ìm; ancho máximo: 16-25 ìm; long. de los procesos: 12-20 ìm. Fig. 14-C.
Registros en Argentina: Arenigiano: como Dactylofusa striatogranulata, Famatina (Achab et al., 2006). Llanvirniano tardío - Caradociano basal: como Dactylofusa cf. D. striatogranulata, Sierra de Zapla (Rubinstein, 2005). Floiano/ Darriwliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M53 (C).
- Eupoikilofusa spp.
Dimensiones (7 ejemplares): Long. Vesícula: 60-90 ìm; ancho vesícula: 10-23 ìm; long. de los procesos: 8-15 ìm. Fig. 11-L.
Descripción. Vesícula fusiforme más o menos elongada, con los bordes rectos a ligeramente curvados que se adelgaza hacia los polos de manera abrupta con los procesos que surgen de los mismos con un adelgazamiento marcado. Los procesos están bien diferenciados de la vesícula, tienen aspecto de espinas, son delgados y terminados en punta, huecos y comunicados con la cavidad de la vesícula. La pared de la vesícula es simple y tiene la superficie cubierta con gránulos, dispuestos en hileras que siguen un patrón longitudinal y que se hacen más finas hacia los polos.
Observaciones. La vesícula se asemeja a E. striatogranulata por la ornamentación de estrías alineadas longitudinalmente pero no se observa su unión en los polos. Algunos ejemplares se asemejan a Dactylofusa striata (Vecoli, 1999).
Registro en Argentina: Tremadociano/ Floiano/ Darriwiliano:Sierra de Zenta (Aráoz, 2009).
Este trabajo: M8 (R), M14 (RR), M18 (R), M68 (R).
GÉNERO LEIOFUSA (EISENACK 1938) EISENACK 1938 EMEND. CRAMER 1970
Especie tipo Leiofusa fusiformis (Eisenack) Eisenack 1938
- Leiofusa fusiformis (Eisenack) Eisenack 1938
1934 Ovum bispidum fusiformis Eisenack, p. 62, pl. 4, fig. 19.
1938 Leiofusa fusiformis Eisenack, p. 28; pl. 4, fig. 10.
Dimensiones (12 ejemplares): Long. Vesícula: 70-150 µm; ancho vesícula: 15-30 µm; long. procesos: 15-25 µm. Fig. 14-E.
Observaciones: Los procesos tienen una longitud muy variable con los extremos acuminados. Se midió la longitud total de la vesícula ya que los procesos no siempre se encuentran bien diferenciados.
Registros en Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Wenlockiano: San Juan (Rubinstein y Brussa, 1999). Este trabajo: M62 (C), M67 (RR), M68 (C), M69 (RR).
- Leiofusa tumida Downie 1959
1959 Leiofusatumida Downie, p.65, pl. 11, fig. 5.
Dimensiones (7 ejemplares):Long. Vesícula: 15-22 µm; ancho vesícula: 10-18 µm; long. procesos: 23-40 µm. Fig. 14-F.
Descripción. Vesícula fusiforme, hueca con un cuerpo central más ancho, ligeramente ovalado, que se adelgazada rápidamente formando los procesos que están bien diferenciados y tienen una longitud mayor que la del cuerpo central, además son huecos y están libremente comunicados con la cavidad de la vesícula.
Registros previos seleccionados. Ashgiliano: Bohemia (Konzalová-Mazancová, 1969); Libia, como L. aff. tumida (Hill y Molyneux, 1988). Devónico Inferior: España (Cramer, 1964). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Ordovícico-Silúrico: Salar del Rincón (Rubinstein y Vaccari, 2004). Este trabajo: M53 (RR), M56 (R), M62 (R).
GÉNERO MULTIPLICISPHAERIDIUM (STAPLIN 1961) STAPLIN, JANSONIUS Y POPOCK 1965 EMEND. EISENACK 1969 EMEND. LISTER 1970 EMEND. TURNER 1984
Especie tipo Multiplicisphaeridium ramispinosum Staplin 1961
- Multiplicisphaeridium? spp.
Dimensiones (22 ejemplares):Diámetro vesícula: 19x50 ìm; long. procesos: 5-12 ìm; ancho base procesos: 3-5 ìm. Fig. 14-J.
Descripción. Vesículas subesféricas, de contorno subcircular a ovalado, con la pared delgada y superficie lisa a finamente granulada. Entre 15 y 24 procesos homomórficos, cilíndricos, huecos y comunicados con la cavidad interior se distribuyen regularmente en todo el cuerpo. La forma de los procesos es variable, generalmente cilíndrica, con las bases ensanchadas y el contacto con la pared de la vesícula es curvado. El extremo distal de los procesos se ramifica en cuatro brazos, con longitud variable, también huecos y con los extremos acuminados.
Observaciones. La preservación es regular, las vesículas generalmente están incompletas, con la superficie corroída y los procesos rotos. Los especímenes tienen una gran variabilidad morfológica. La ramificación terminal de los procesos es cerrada como en Multiplicisphaeridium, sin embargo no puede descartarse similitudes con formas como Cymatiogaleamessaoudensis, de la que se diferencia porque Multiplicisphaeridium? tiene menor número de procesos y no se observa el ensanchamiento en sus bases ni ramificaciones expandidas en los extremos, ni la apertura polar.
Registro en Argentina: Floiano/ Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M34 (RR), M51 (R), M53 (C), M55 (C), M56 (R), M62 (CC).
GÉNERO PETALOFERIDIUM JACOBSON 1978
Especie tipo Petaloferidium stigii Jacobson 1978
- Petaloferidium bulliferum Yin, Di Milia y Tongiorgi 1998
1998 Petaloferidium bulliferumYin, Di Milia y Tongiorgi. pp. 227-229, pl. 1,figs.1, 2a,b; pl. 2, fig. 1.
Dimensiones (1 ejemplar): Diámetro vesícula: 18x23 ìm; long. procesos: 2-3 ìm.
Observaciones. La mala preservación del ejemplar no permitió una buena determinación del número de procesos, contándose alrededor de 12.
Registros con control independiente. Yushanian Medio, zona A. suecicus, Sur de China (Yin et al., 1998). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M56 (RR).
GÉNERO PICOSTELLA (CRAMER, ALLAM, KANES Y DÍEZ 1974) YIN, DI MILIA Y TONGIORGI 1998
Especie tipo Picostella perforata Cramer, Allam, Kanes y Díez 1974
- Picostella turgida Yin, Di Milia y Tongiorgi 1998
1998 Picostella turgida Yin, Di Milia y Tongiorgi, p. 233-236, pl. 2: 4; pl. 3: 2-5; pl. 4: 1-3, 4a-b,5, 6.
Dimensiones (10 ejemplares): Diámetro vesícula: 20-25 ìm; cant. procesos: 8-12; long. procesos:15-20 ìm. Fig. 13-A.
Registros con control independiente. Registro más bajo:Arenigiano temprano, Zona de Didymograptus bifidus, Río de la Huerta, Argentina (Rubinstein y Toro, 2001). Registro más alto: Arenigiano temprano, Zona de A. suecicus, Sur de China (Yin et al., 1998). En Argentina: Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M51 (RR), M53 (RR), M67 (R), M68 (R).
GÉNERO PIREA VAVRDOVÁ 1972
Especie tipo Pirea dubia Vavrdová 1972
Observaciones. Servais y Molyneux (1997) definen tres categorías según sean formas lisas (Pirea sp. cf. P.colliformis), formas ornamentadas (Pirea sp. cf. P. ornata) y formas con estrías por alineamiento de granos en la base del proceso (Pirea sp. cf. P. sinensis).
- Pirea sp. cf. P. colliformis (Burmann 1970) Eisenack, Cramer y Díez 1976
1972 Pirea dubia Vavrdová, p. 83, pl. 1, fig. 4.
1976 Pirea colliformis (Burmann) Eisenack, Cramer y Díez, p. 318, pl. 13, fig. 4.
1997 Pirea sp. cf. P. colliformis (Burmann) Eisenack, Cramer y Díez 1976 - Servais y Molyneux, p. 130, pl. 6, fig. 7.
Dimensiones (1 ejemplar):Long. Vesícula: 38 ìm; ancho máximo: 25 ìm; long. proceso: 5 ìm. Fig. 13-B.
Descripción. Vesícula de contorno oval y forma de pera, con un estrechamiento en la parte apical diferenciando un proceso grueso y corto con la base ligeramente expandida. La vesícula es hueca igual que el proceso que es distalmente cerrado, con libre comunicación entre ellos. La superficie de la vesícula y el proceso son lisas, con algunos pliegues producto de la compresión.
Registros previos seleccionados. Formas lisas de Pirea fueron son registradas en el Ordovícico de peri-Gondwana y Sur de China por Tongiorgi et al. (1995) entre otros. En Argentina: Pirea sp. cf. P. dubia, Salta (Volkheimer et al., 1980). Arenigiano Medio: Sección La Huerta (Rubinstein y Toro, 2001). Floiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR).
- Pirea sp. cf. P. ornata (Burmann 1970) Eisenack, Cramer y Díez 1976
1976 Pirea ornata (Burmann) Eisenack, Cramer y Díez, pp. 320, pl. 15, figs. 9-12.
1997 Pirea sp. cf. P. ornata (Burmann) Eisenack, Cramer y Díez 1976 - Servais y Molyneux, p. 130, pl. 6, fig. 5.
Dimensiones (1 ejemplares): Long. Vesícula: 40 µm;ancho máximo: 35 µm; long. proceso: 6 ìm. Fig. 14-L.
Descripción. Vesícula de contorno oval que se extiende en el polo apical formando un proceso corto. Este proceso poco diferenciado tiene su extremo distal redondeado dando a la vesícula una apariencia de petaca o pera. La superficie está cubierta regularmente por una ornamentación de pequeñas agujas delgadas de menos de 1 µm de longitud, que tienen las bases bulbosas o como granos. Los elementos esculturales de la superficie tienden a hacerse más largos y cercanos entre sí en la porción antiapical de la vesícula, mientras que el
Registros previos seleccionados. Arenigiano: Pirea ornata, Cerdeña (Albani, 1989). Arenigiano-Llanvirniano: República Checa (Vavrdová, 1977; 1979; 1982; 1990a). Llanvirniano: Alemania (Burmann, 1970; Servais y Maletz, 1993); Plataforma Rusa (Burmann, 1970); Norte de Africa (Vecoli, 1999). Ordovícico: como Pirea sp. cf. P. ornata, peri-Gondwana y Sur de China (Tongiorgi et al., 1995). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M62 (RR).
GÉNERO SAHARIDIA COMBAZ 1967
Especie tipo Saharidia downiei Combaz 1967
- Saharidia fragilis (Downie) Combaz 1967
1958 Leiosphaeridia fragile Downie, pp. 344-345, pl. 17, fig. 6.
1967 Saharidia fragilis (Downie) Combaz, p. 13; pl. 2, figs. 35-36.
Dimensiones (65 ejemplares): Diámetro: 30-85 µm; ancho anillo: 5-8 ìm. Fig. 10-L.
Observaciones. La pared es muy frágil de modo que la vesícula aparece muchas veces colapsada, quebrada o plegada, de modo que la observación del piloma y otros detalles morfológicos es dificultosa. Se observa una gran variabilidad morfológica, especialmente en el rango de tamaños, la ornamentación y la presencia del anillo periférico. En algunos ejemplares se observa la presencia de un piloma en el sector central, con un diámetro menor a un cuarto del total del diámetro de la vesícula.
Registros con control independiente. Registro más bajo: Cámbrico tardío, Zona Peltura, Terranova (Martin y Dean, 1988). Registro más alto: Tremadociano, Inglaterra (Downie, 1958); Argelia (Combaz, 1967); Francia (Martin, 1972). En Argentina: Cámbrico-Tremadociano: Quebrada de Moya (Rubinstein et al., 2003; Aráoz y Vergel, 2006; Vergel et al., 2007). Tremadociano: Sierra de Zenta (Aceñolaza et al., 2003; Vergel et al., 2008 b; Aráoz, 2009). Este trabajo: M2 (AA); M5 (RR).
GÉNERO STELLECHINATUM TURNER 1984
Especie tipo Stellechinatum celestum (Martin) Turner 1984
- Stellechinatum uncinatum (Downie) Molyneux 1987
1958 Hystrichosphaeridium longispinosum var. uncinatum Downie, p. 337, text-fig. 2a.
1987 Stellechinatum uncinatum (Downie) Molyneux, pp. 342-344, text- figs. 93, 94.
Dimensiones (29 ejemplares): Diámetro: 15x35 µm; long. procesos: 9-18 ìm. Fig. 15-C.
Registros con control independiente. Registro más bajo: Cámbrico tardío: Terranova, Zona Parabolina spinulosa (Martin y Dean, 1988). Registro más alto: Caradociano medio: Mar de Botnia (Tynni, 1975). En Argentina: Arenigiano: Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Qda. Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001). Floiano: Sierra de Zenta (Aráoz, 2009). Darriwiliano: Sierra de Zenta (Vergel et al., 2008 b; Aráoz, 2009). Este trabajo: M31 (C), M32 (R), M51 (R), M53 (C), M61 (R), M62 (C), M66 (RR), M67 (R), M85 (R), M87 (R).
- Stellechinatum sp. 1
Dimensiones (5 ejemplares): Diámetro vesícula: 23x28 ìm; cant. procesos: 9-18; long. procesos: 10-19 ìm. Fig. 15-B.
Descripción. Vesícula de contorno poligonal con procesos regularmente distribuidos en la superficie. Estos son homomórficos simples y cónicos con los extremos terminados en finas puntas, huecos y libremente comunicados con la cavidad interna de la vesícula. Las bases de los procesos están ligeramente expandidas con un contacto curvado con la pared. La superficie de la vesícula presenta una fina granulación, mientras que los procesos tienen una fina ornamentación de pelos delgados desde la base, con longitud menor a 1 µm ás abundantes en los extremos.
Observaciones. Se diferencia de S. uncinatum en el menor número de procesos y en la ornamentación menos desarrollada de espinas de menor tamaño. Se diferencia de S. brachyscolum en que éste último tiene procesos más gruesos, con las bases más marcadas y las espinas de los procesos más robustas en la base.
Registro en Argentina: Floiano/ Darriwiliano:Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M53 (RR).
GÉNERO STELLIFERIDIUM DEUNFF, GóRKA Y RAUSCHER 1974
Especie tipo Stelliferidium striatulum (Vavrdova 1966) Deunff Górka y Rauscher 1974
- Stelliferidium stelligerum (Górka 1967) Deunff, Górka y Rauscher 1974
1967 Cymatiogalea stelligera Górka, pp. 2-3, pl. 1, figs. 1-4.
1972 Cymatiogalea stelligera (Górka) Martin pp. 40-41, pl. 1, figs. 1-14, pl. 3, fig. 12, pl. 4, fig. 15.
1974 Stelliferidium stelligerum (Górka) Deunff, Górka y Rauscher, p. 15.
Dimensiones (3 ejemplares):Diámetro vesícula: 20x40 ìm; diametro apertura: 35 ìm; cant. procesos: mas de 30; long. procesos: 8-11 ìm. Fig. 15-D.
Observaciones. Es característica a presencia de estrías radiales en la base de los procesos que se unen con la de los procesos adyacentes. Muchos de los procesos aparecen rotos dificultando su recuento, además de algunos que aparecen incompletos, sin los extremos. La variabilidad de la morfología y tamaño de los procesos de S. stelligerum permite incluir este ejemplar en la especie aún cuando no se observan detalles de los procesos.
Registros con control independiente. Tremadociano Superior, Zona Paltodus deltifer, Suecia (Bagnoli et al., 1988). Registros en Argentina: Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M56 (RR).
- Stelliferidium striatulum (Vavrdová 1966) Deunff, Górka y Rauscher 1974
1966 Balthisphaeridium striatulum Vavrdová, pp. 411-412, pl. 1, figs. 2a, b, pl. 2, fig. 3, text-fig. 3a.
1974 Stelliferidium striatulum (Vavrdová) Deunff et al., p. 16, pl. 6. figs. 2, 19, 20.
Dimensiones (1 ejemplar): Diámetro vesícula: 28x34 ìm; long. procesos: 4-6 ìm; ancho procesos: 1-1,5 ìm. Fig. 15-E.
Registros en Argentina: Arenigiano: Famatina (Achab et al., 2006). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M53 (RR).
- Stelliferidium spp.
Dimensiones (9 ejemplares):Diámetro vesícula: 25-35 ìm; cant. procesos: 15-35; long. procesos: 7-9 ìm. Fig. 13-E.
Descripción. Vesícula hemiesférica que sostiene procesos de forma y tamaño variable. La superficie está cubierta por estrías que tienen una disposición radial. Los procesos son cilíndricos, huecos sin comunicación con el interior de la vesícula, con los extremos ramificados en 3 o 4 brazos con terminaciones cerradas y redondeadas.
Observaciones. El número de procesos dado en la descripción es aproximado, ya que muchos aparecen fracturados o ausentes.
Registro en Argentina: Dapingiano/ Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M18 (RR), M37 (RR), M62 (RR), M66 (RR), M68 (RR).
GÉNERO STRIATOTHECA BURMANN 1970
Especie tipo Striatotheca principalis Burmann 1970
- Striatotheca principalis Burmann 1970
1970 Striatotheca principalis Burmann, p. 300, pl. 11, fig. 1.
1970 Striatotheca principalis var. parva Burmann, p. 300, pl. 8, fig. 6.
1970 Striatotheca principalis var. principalis Burmann, p. 300, pl. 11, fig. 1.
Dimensiones (1 ejemplar): Long. Vesícula: 35 ìm; ancho vesícula: 30 ìm; long. procesos: 12-18 ìm. Fig. 13-F.
Observaciones. Burmann (1970) separó dos variedades intraespecíficas, Striatotheca principalis var. principalis y S. principalis var. parva, según el tamaño y la densidad de las estrías. Ottone (en Ottone et al., 1992) considera que ambas variedades son prácticamente indistinguibles, con variaciones graduales de tamaño y Tongiorgi et al. (2003) remarca que podrían aceptarse como variedades subespecíficas. Para la identificación de los especímenes de S. principalis de la Sierra de Zenta, se siguió a Ottone en Ottone et al. (1992) y no se considera las variantes intraespecíficas.
Registros con control independiente. Registro más bajo: Arenigiano temprano, Terranova (Martin, 1982) y Zona D. deflexus, Sur de China (Li, 1987). Registro más alto: Llanvirniano temprano, Zona D. murchisoni, Túnez (Vecoli, 1999). En Argentina: Arenigiano: Secciones La Huerta, Río Acoite, Parcha (Rubinstein et al., 1999); Qda. Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001); Los Colorados (Ottone et al., 1992; Rubinstein et al., 2007 b); Famatina, como S. principalis parva (Achab et al., 2006). Floiano: Sierra de Zapla, como S. principalis parva (Rubinstein, 2005; Rubinstein et al., 2007 b; Rubinstein y de la Puente, 2008). Floiano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M37 (RR).
- Striatotheca rugosa Tongiorgi, Yin, Di Milia 1995
1995 Striatotheca rugosa Tongiorgi et al., p. 42; pl. 4; figs. 8-11.
Dimensiones (3 ejemplares):Diámetro vesícula: 20x33 µm; long. procesos: 25-28 ìm. Fig. 12-A.
Observaciones. Se observa una ornamentación de granos alineados a lo largo de las estrías.
Registros previos seleccionados. Tremadociano Tardío - Arenigiano: Pakistán (Quintavalle et al., 2000). Arenigiano Temprano: Sur de China (Tongiorgi et al., 1995; 2003). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M68 (RR), M87 (RR).
- Striatotheca sp. cf. S. transformata Burmann 1970
1970 Striatotheca transformata Burmann, pp. 302, pl. 10, fig. 1.
Dimensiones (1 ejemplar):Long. lados: 25-28 ìm; long. procesos: 5-7 ìm; numero de estrías: 8-10; distancia entre estrías: 1-1,5 µm. Fig. 11-B.
Descripción. Vesícula de contorno irregular, originalmente trapezoidal de aspecto globoso, con procesos que se levantan gradualmente en distintos planos, a partir de adelgazamientos en los ángulos. Los procesos son cortos, cónicos y terminados en una punta y la pared de la vesícula es delgada con la superficie surcada por estrías bien definidas cercanas unas a otras y que tienen una leve curvatura hacia la parte media de los lados.a diferencia de la pared de los procesos que es lisa.
Observaciones. Aún cuando el espécimen muestra una buena correspondencia con la especie, la vesícula fracturada y abierta no permite confirmar la asignación a la especie.
Registros en Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M66 (RR).
GÉNERO TECTITHECA BURMANN 1968
Especie tipo Tectitheca valida Burmann 1968
- Tectitheca additionalis Burmann 1968
1968 Tectitheca additionalis Burmann, p. 649, pl. 6, fig. 5.
Dimensiones (17 ejemplares): Long. Vesícula: 32-40 ìm; ancho vesícula: 20-25 ìm; long. procesos: 10-23 ìm. Fig. 13-G.
Registros En Argentina: Arenigiano: Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Quebrada Grande y Quebrada de la Curva (Rubinstein y Toro, 1999; 2001). Floiano Darriwiliano: Sierra de Zapla, como T. addtionalis-valida (Rubinstein y de la Puente, 2008). Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M67 (R), M68 (R), M85 (R).
- Tectitheca valida Burmann 1968
1968 Tectitheca valida Burmann, p. 648, pl. 6, fig. 2.
Dimensiones (2 ejemplares):Long. Vesícula: 45 ìm; ancho vesícula: 30 ìm; long. procesos: 35-40 ìm. Fig. 11-I.
Registros en Argentina. Arenigiano: Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Quebrada Grande y Quebrada de la Curva (Rubinstein y Toro, 1999; 2001) Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M85 (RR), M86 (RR).
- Tectitheca sp. cf. T. additionalis Burmann 1968
Dimensiones (8 ejemplares):Long. Vesícula: 25-30ìm; ancho vesícula: 18-22 ìm; long. procesos: 10-22 ìm.
Descripción. Cuerpo central de contorno pentagonal, donde se diferencia la parte superior cónica y la parte inferior elongada, ligeramente redondeada. De la parte cónica surge gradualmente un proceso apical de longitud variable y terminado en punta. Por debajo del plano ecuatorial se observan 8 o más procesos y otros tantos idénticos en la base del cuerpo y otros procesos adicionales en la mitad antiapical del cuerpo.
Registros en Argentina: Dapingiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo. M18 (RR), M31 (RR),M68 (RR),M81 (R),M85 (RR).
GÉNERO VERYHACHIUM (DEUNFF 1954) EMEND. DOWNIE Y SARJEANT 1963 EMEND. TURNER 1984
Especie tipo Veryhachium trisulcum Deunff 1951 ex Deunff 1959
- Veryhachium lairdii (Deflandre 1946) Deunff 1959 ex Downie 1959
1946 Hystrichosphaeridium lairdi Deflandre card. 1112 (nom.nud).
1959 Veryhachiumlairdi (Deflandre) Deunff, p. 28, pl. 8, figs. 75-79.
Dimensiones (27 ejemplares): Long. Vesícula: 18-40 ìm; ancho vesícula: 15-30 ìm; long. procesos: 18-45 ìm. Fig. 12-E.
Registros con control independiente. Yushanian medio, Zona A. suecicus, Zona O. evae, Sur de China (Tongiorgi et al., 1995). En Argentina: Arenigiano: Los Colorados y Río Cajas (Ottone et al., 1992); Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Qda. Grande y Qda. de la Curva (Rubinstein y Toro, 1999; 2001); Famatina (Achab et al., 2006). Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M34 (RR), M62 (C), M67 (C), M68 (C), M81 (R), M85 (RR).
- Veryhachium minutum Downie 1958
1958 Veryhachium minutum Downie, p. 344, pl. 17, fig. 4, text-fig. 3C.
Dimensiones (7 ejemplares):Long. Vesícula: 15-20 ìm; ancho vesícula: 10-13 ìm; long. procesos: 5-10 ìm.
Registros previos seleccionados. Tremadociano: Gales (Downie, 1958); Francia (Martin, 1972). Tremadociano - Llanvirniano: Bélgica (Martin, 1969). Arenigiano: Bohemia (Vavrdová, 1972).Llandeilo: Gales (Turner, 1985). Caradociano: Bélgica (Martin et al., 1970). Devónico Superior: (Stockmans y Willière, 1962). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M68 (RR), M90 (C).
- Veryhachium rhomboidium (Downie 1959) Turner 1984
1959 Veryhachium rhomboidium - Downie, p. 62, pl., 12, fig. 10.
1984 Veryhachium rhomboidium - Turner, p. 145.
Dimensiones (1 ejemplar):Long. Vesícula: 20 ìm; ancho vesícula: 15 ìm; long. procesos: 20-35 ìm. Fig. 15-G.
Registros previos seleccionados. Caradociano: Inglaterra (Turner, 1984). Llandoveriano-Wenlock: Bélgica (Martin, 1969). Wenlock: Inglaterra (Downie, 1959). Devónico Superior: Bélgica (Stockmans y Willière, 1962). Pérmico: Gran Bretaña (Wall y Downie, 1963). En Argentina: Arenigiano:Área Santa Victoria, como Veryhachium sp. cf. rhomboidium (Rubinstein et al., 1999). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M62 (RR).
- Veryhachium symmetricum (Lu 1987) Tongiorgi, Yin y Di Milia 1995
1987 Veryhachium symmetricum Lu, p. 99, pl. 3 figs. 9,18.
1995 Veryhachium symmetricum (Lu Li-chang) Tongiorgi et al., pp. 43-45, pl. 4, figs. 7, 12-13.
Dimensiones (9 ejemplares):Long. Vesícula: 18-20 ìm; cant. procesos: 8-14; long. procesos: 11-20 ìm. Fig. 13-I.
Observaciones. Los ejemplares descriptos tienen tamaños menores a los de la diagnosis, aunque la morfología se corresponde a la especie.
Registros con control independiente. Yushanian medio, Zona A. suecicus, Zona O. evae, Sur de China (Tongiorgi et al., 1995). En Argentina: Floiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M31 (RR), M68 (R), M85 (R), M87 (RR).
Grupo Veryhachium trispinosum (Eisenack 1938) Deunff 1954 ex Downie 1959 1938 Hystrichosphaeridium trispinosum Eisenack, p. 14, fig. 2,3.
1959 Veryhachium trispinosum (Eisenack) Downie, pp. 68-69, pl. 12, fig. 7.
Dimensiones (9 ejemplares):Long. lados: 25-40 ìm; long. procesos: 15-20 ìm. Fig. 15-H.
Registros en Argentina: Arenigiano: Los Colorados y Río Cajas (Ottone et al., 1992); Secciones La Huerta, Río Acoite y Parcha (Rubinstein et al., 1999); Quebrada Grande y Quebrada de la Curva (Rubinstein y Toro, 1999; 2001); Famatina (Achab et al., 2006). Darriwiliano: Sierra de Zapla (Rubinstein y de la Puente, 2008). Aeroniano - Telychiano (Silúrico): San Juan (Rubinstein y Brussa, 1999). Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M62 (R), M68 (R).
GÉNERO PTEROSPERMELLA EISENACK 1972
Especie tipo Pterospermella aureolata (Cookson y Eisenack) Eisenack 1972
División Chlorophyta Pascher 1914
Clase Prasinophyceae Christensen 1962
Orden Pterospermatales Schiller 1925
Familia Pterospermellaceae Eisenack 1972
- Pterospermella colbathii Vavrdová 1990
1990 Pterospermella colbathii Vavrdová, pp. 240-241, pl. 2, figs. 3, 4, 7, 8.
Dimensiones (1 ejemplares):Diámetro vesícula: 30x35 µm; ancho membrana:5-7 ìm.Fig. 11-K.
Descripción. Vesícula con el cuerpo central esférico de pared gruesa y rodeada por una delgada y translúcida membrana que tiene un ancho variable y es soportada por engrosamientos que salen del cuerpo central.
Registros con control independiente. Llanvirniano temprano: República Checa, niveles con Expansograptus ferrugineus (Vavrdová, 1990 a). Registros previos seleccionados: Arenigiano: República Checa (Vavrdová, 1990 a). Llanvirniano: Argelia (Vecoli, 1999). En Argentina: Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo: M85 (RR).
- Pterospermella sp. 1
Dimensiones (11 ejemplares): Dimensiones: longitud vesícula: 80-115 µm; ancho vesícula: 20-32 µm. Fig. 12-G.
Descripción. Vesícula fusiforme, con dos polos adelgazados y terminando en punta pero sin procesos. La pared de la vesícula es doble, con una capa interna más gruesa, que tiene la superficie psilada y es cubierta por una segunda pared más delgada, translúcida y lisa.
Observaciones. En general estos ejemplares tienen pobre preservación. Se asignó esta forma a Pterospermella sp. 1 por la pared de doble capa.
Registros en Argentina: Tremadociano/ Floiano/ Dapingiano/ Darriwiliano: Sierra de Zenta (Aráoz, 2009). Este trabajo. M2 (RR), M5 (RR), M13 (RR), M21 (RR), M51 (RR), M61 (RR), M66 (RR), M68 (R).
Conclusiones
Se confirma la presencia de depósitos ordovícicos en la Sierra de Zenta, revalidando esta edad para algunos niveles registrados anteriormente como de edad ordovícica indiferenciada, Ordovícico del Grupo Santa Victoria o siluro-devónica.
Se amplía el conocimiento de la estratigrafía de la Sierra de Zenta reconocida como una zona de características transicionales entre las provincias geológicas Cordillera Oriental y Sierras Subandinas. En el sector norte de la sierra hay una mayor semejanza con Cordillera Oriental donde se identificaron las formaciones Santa Rosita y Acoite (Grupo Santa Victoria); y en el sector centro sur con Sierras Subandinas donde las unidades fueron definidas informalmente como Unidades Formacionales I, II y III.
En base a las asociaciones palinológicas identificadas y coincidente con la edad dada por los trilobites de la Biozona de Kainella meridionalis, se confirma que en el Abra de Zenta - Abra Límite las edades son Tremadociano, Floiano y Dapingiano.
Se rectifica que las edades siluro - devónicas señaladas en la sección de Abra de Zenta por la literatura geológica (op. cit.) corresponden a la sección que en este trabajo se menciona como Abra Límite, ubicada al sur de la citada localidad.
Se recuperaron diferentes grupos de palinomorfos como acritarcos, quitinozoos, algas prasinofíceas y criptosporas, con un notable predominio de acritarcos, presentes con una abundancia y diversidad taxonómica muy variable.
En base a la distribución temporal (aparición/desaparición) de acritarcos seleccionados, se definieron informalmente cinco asociaciones palinológicas. Entre Abra de Zenta y Abra Límite (Perfil N° 1) se identificaron las asociaciones ASZ1, ASZ2 y ASZ3 y entre Abra Buey Ojo y Laguna Verde (Perfiles N° 2, 3, 4 y 5) las asociaciones ASZ4 y ASZ5.
Las edades asignadas a las asociaciones palinológicas son: ASZ1: Tremadociano Inferior; ASZ2: Floiano temprano sin descartar la posibilidad de un Tremadociano tardío y ASZ3: Dapingiano a Floiano tardío.
La diferencia composicional entre las ASZ2 y ASZ4 permite aceptar para la ASZ4 una edad ligeramente más jóven que la de la ASZ2.
En los niveles más altos de la Unidad Formacional III en el Perfil N° 5, donde se identifica la ASZ5, se encuentra el conodonte Erismodus aff. quadridactylus que ratifica una edad darriwiliana alta/ sandbiana.
Se observaron importantes diferencias en la composición de las asociaciones de Sierra de Zenta con respecto a otras coetáneas, con una ausencia de formas típicas de la edad a nivel mundial. Estas diferencias podrían deberse a factores paleoambientales locales, ya que la región de la Sierra de Zenta correspondería a un sector más interno de la Cuenca Andina en tiempos ordovícicos.
Se destaca la alta representatividad de los acritarcos Eisenackidium orientalis y Dactylofusa velifera forma brevis en la mayoría de las muestras. Para Eisenackidium orientalis, caracteristico del Floiano, se extiende su registro temporal al menos hasta el Darriwiliano.
La superabundancia de Saharidia, la baja diversidad taxonómica y niveles que incluyen casi exclusivamente Micrhystridium en la parte inferior y media del Perfil N°1 confirman paleoambientes relativamente someros. En el Perfil N°4 la proporción relativamente alta de formas complejas como Polygonium, Cymatiogalea, Stellechinatum, Veryhachium y Tectitheca permiten interpretar ambientes abiertos y con cierta distancia de la costa. En la parte superior del Perfil N°5 las formas presentes y las aracterísticas sedimentológicas permiten inferir una somerización.
La presencia de Cymatiogalea deunffii, Grupo Vavrdovella areniga, Pirea, Striatotheca, Coryphidium y Veryhachium en las asociaciones permite establecer afinidades con la Provincia Mediterránea o peri-Gondwana, que representan una asociación de aguas frías para el Ordovícico temprano.
Bibliografía
1. Aceñolaza, F. y Aceñolaza, G. 2005. La formación Puncoviscana y unidades estratigráficas vinculadas en el Neoproterozoico - Cámbrico Temprano del Noroeste Argentino. Latin American Journal of Sedimentology and Basin Análisis , 12 (2): 65-87. [ Links ]
2. Aceñolaza, F.G. y Aceñolaza, G.F.; 2008. El Ediacarano Cámbrico temprano de la Provincia de Jujuy: caracteres paleontológicos, paleoecológicos y paleogeográficos. XVII Congreso Geológico Argentino, Relatorio 2: 29-36. [ Links ]
3. Aceñolaza, G.F. y Aceñolaza, F.G. 2002. Ordovician trace fossils of Argentina. En: Aceñolaza, F.G. (Ed.): Aspects ofthe ordovician system in Argentina . Serie Correlación Geológica, 16: 177-194. [ Links ]
4. Aceñolaza, G.F. y Aceñolaza, F.G. 2007. Bivalvian trace fossils of the Labrado Formation (Lower Ordovician) from Zenta Range (Jujuy province): preservational controls of some push-and-pull deposit-feeder traces. VReunión Argentina de Icnología y Tercera Reunión de Icnología del Mercosur. Laboratorio de Geología Andina, CADIC / CONICET, Ushuaia, 17. [ Links ]
5. Aceñolaza, G.F., Aráoz, L., Vergel, M.d.M., Tortello, M.F. y Nieva, S.M. 2003. Paleontology and biostratigraphy of the Lower Ordovician strata cropping out at the Abra de Zenta (Cordillera Oriental, Jujuy and Salta provinces), NW Argentina. En: Albanesi, G.L.; Beresi, M.S. and Peralta, S.H. (Eds.): Ordovician from the Andes. Serie de Correlacion Geológica, 17: 23-28. [ Links ]
6. Aceñolaza, G.F., Aris, J. y Milana, J.P. 2006. Taphonomy of shell beds and the open surface furrow origin in Cruziana (trace fossil): a test case in the Ordovician of NW Argentina. 9° Congreso Argentino de Paleontología y Bioestratigrafía . Publicación Especial de la Academia Nacional de Ciencias, 237. [ Links ]
7. Aceñolaza, G.F., Fogliata, A., Nieva, S.M. y Mas, G. 2008. Los nódulos fosfáticos en el Ordovícico de la Sierra de Zenta (Sistema Interandino de las Provincias de Jujuy y Salta): caracteres geoquímicos y ambientes de depositación. XVII Congreso Geológico Argentino, Jujuy, Actas I: 335-336. [ Links ]
8. Aceñolaza G.F. y Gutiérrez-Marco, J.C. 2002. Ordovician Echinoderms of Argentina. En: Aceñolaza, F.G. (Ed.): Aspects of the Ordovician System in Argentina . Serie de Correlación Geológica, 16: 121-130. [ Links ]
9. Aceñolaza, G.F. y Heredia, S. 2008. The status of the Cruziana (trilobite trace fossil) stratigraphy in Western Gondwana: The mixing of Lower and Upper Ordovician Elements in the Central Andean Basin of South America. En: I. Rábano, R. Gozalo and D. García Bellido (Eds.) Advances in Trilobite Research. Cuadernos del Museo Geominero, 9: 13-17. Madrid. [ Links ]
10. Aceñolaza, G.F. y Milana, J.P. 2005. Remarkable Cruziana beds in the Lower Ordovician of the Cordillera Oriental, NW Argentina. Ameghiniana, 42 (3): 633-637. [ Links ]
11. Aceñolaza, G.F., Tortello, M.F., Vergel, M. d. M. y Aráoz, L. 2001. Icnofósiles, trilobites y acritarcos de la Formación Santa Rosita aflorante en la región del Abra de Zenta (provincias Salta y Jujuy). IV Reunión Argentina de Icnología y 2da Reunión de Icnología del Mercosur , Resúmenes, 22. [ Links ]
12. Achab, A., Rubinstein, C.V. y Astini, R.A. 2006. Chitinozoans and acritarchs from the Ordovician peri-Gondwana volcanic arc of the Famatina System, nosthwestern Argentina. Review of Paleobotany and Palynology, 139: 129-149. [ Links ]
13. Albanesi, G.L., Ortega, G. y Zeballo, F.J. 2008. Faunas de conodontes y graptolitos del Paleozoico Inferior en la Cordillera Oriental Argentina. XVII Congreso Geológico Argentino. Relatorio, 2: 98-118. [ Links ]
14. Albanesi, G.L., Zeballo, F.J., Monaldi, C.R. y Ortega, G. 2007.La Zona de conodontes de Paroistodus proteus - Acodusdeltatus y graptolitos asociados en el Tremadociano superior del noroeste argentino. Ameghiniana Suplemento 44 (4), Resumenes 87 R. [ Links ]
15. Albani, R. 1989. Ordovician (Arenigian) acritarchs from the Solanas Sandstone Formation, Central Sardinia, Italy. Bolletino della Società Paleontologica Italiana , 28 (1): 3-37. [ Links ]
16. Amengual, R. y Zanettini, J.C. 1973. Geología de la Comarca de Cianzo y Caspala (Provincia de Jujuy). Revistade laAsociación Geológica Argentina ,28 (4): 341-352. [ Links ]
17. Aráoz, L.; 2002: Palinología de los niveles inferiores de la Formación Santa Rosita, en el Abra de Zenta, Provincias de Salta y Jujuy,República Argentina. Seminario Inédito, Facultad de Ciencias Naturales e I.M.L., Universidad Nacional de Tucumán, 187 pp. [ Links ]
18. Aráoz, L. 2008. Aspectos palinológicos del Paleozoico Inferior de la Sierra de Zenta, Cordillera Oriental Argentina. Acta Geológica Lilloana, 20 (2): 7. Suplemento. [ Links ]
19. Aráoz, L. 2009. Estudios microflorísticos del Ordovícico de la Sierra de Zenta, Cordillera Oriental Argentina. Tesis Doctoral. Inédita, 364 p. [ Links ]
20. Aráoz, L. y Aceñolaza; G.F. 2004. Primeros registros palinologicos para el Tremadociano Superior de la Sierra de Zenta, Cordillera Oriental Argentina. XI Reuniao de Paleobotanicos e Palinologos, Gramado, Brasil, Boletim de Resumos: 9. [ Links ]
21. Aráoz, L., Aceñolaza, G.F., Vergel, M.d.M., Heredia, S., Tortello, F. y Milana, J.P. 2008. El Ordovícico del sector central de la Sierra de Zenta (Sistema Interandino de las provincias de Jujuy y Salta): cronoestratigrafía y correlación. XVII Congreso Geológico Argentino, Jujuy, Actas I: 339-340. [ Links ]
22. Aráoz, L. y Vergel, M.d.M. 2001. Acritarcos del Ordovicico Inferior (Formación Santa Rosita) en el Abra de Zenta, Cordillera Oriental, Argentina. V Jornadas de Comunicaciones, Facultad de Ciencias Naturales e I.M.L. Serie Monográfica y Didáctica , 41: 55. [ Links ]
23. Aráoz, L. y Vergel, M.d.M. 2006. Palinología de la transición cambro - ordovícica en Quebrada de Moya, Cordillera Oriental, Argentina. Revista Brasileira de Paleontología, 9 (1): 1-8. [ Links ]
24. Astini. R.A. 2008. Sedimentación, facies, discordancias y evolución paleoambiental durante el cambro-ordovícico. XVII Congreso Geológico Argentino , Relatorio, 2: 50-72. [ Links ]
25. Astini, R.A. y Marengo, L. 2006. Paleoambientes y estratigrafía secuencial de depósitos marinos marginales del Ordovícico de la sierra de Zapla (Sierras Subandinas, noroeste argentino) y su relación con la Cuenca Andina Central. Revista Geológica de Chile, 33 (2): 247-276. [ Links ]
26. Astini, R.A., Waisfeld, B., Toro, B.A. y Benedetto, J.L. 2004. El Paleozoico inferior y medio de la región de Los Colorados, borde occidental de la Cordillera Oriental (provincia de Jujuy). Revista de la Asociación GeológicaArgentina 59 (2): 243-260. [ Links ]
27. Bagnoli, G., Stouge, S. y Tongiorgi, M. 1988. Acritarchs and conodonts from the cambro - ordovician Furuhäll (Köpingsklint) section (Öland, Sweden). Riv. It. Paleont. Strat., 94 (2): 163-248. [ Links ]
28. Benedetto, J.L., Aceñolaza, G.F., Albanesi, G.L., Alfaro, M.B., Brussa, E.D., Buatois, L.A., Carrera, M.G., Cech, N., Esteban, S.B., Heredia, S., Mangano, M.G., Ortega, G., Ottone, E.G., Rubinstein, C.V., Salas, M.J., Sanchez, T.M., Toro, B.A., Tortello, M.F., Vaccari, N.E. y Waisfeld, B.G. 2007. Los fósiles del Proterozoico Superior y Paleozoico Inferior de Argentina. Ameghiniana, Publicación Especial, 11: 9-32. [ Links ]
29. Boso, M.A. y Monaldi, C.R. 2008. Cuenca Silúrico Devónica y los depósitos ferríferos asociados en la provincia de Jujuy. XVII Congreso Geológico Argentino, Relatorio 2: 155-165. [ Links ]
30. Brackebush, L. 1892. Die Korddillerenpaensse zwischen der Argentinischen Republic und Chile, von 22° - 35° s. Br. Zeittschrift Deutsche Gessellschaft für ErdkundeXXVII: 250-348, Berlín. [ Links ]
31. Brito, I.M. y Santos, A.S. 1965. Contribuicao ao conhecimento dos micrófosseis silurianos e devonianos da Bacia do Maranhão. I Leiofusidae. (Notas prelim. Estud.).Div. Geol. Miner., 129: 3-25. [ Links ]
32. Brocke, R. 1992. First results of palynological investigations of the lower Arenig from the Yangtze platform, China. 8thInternaitonal Palynological Congress Aix-en-Provence, Abstracts, 18. [ Links ]
33. Brocke, R., Fatka, O. y Servais, T. 1998. A review of the ordovician acritarchs Aureotesta and Marrocanium. Annales dela Société Géologique de Belgique , 120 (1): 1-21. [ Links ]
34. Brocke, R., Li, J. y Wang, Y. 2000. Upper Arenigian to lower Llanvirnian acritarch assemblages from South China: a preliminary evaluation. Review of Palaeobotany and Palynology,113: 27-40. [ Links ]
35. Bultynck, P. y Martin, F. 1982. Conodontes et Acritarches de l' Ordovicien Inférieur et acritarches du Silurien inférieur de la partie septentrionale de la Cordillère Argentine. Bulletin de l' Institut royal des Sciences naturelles deBelgique , Sciences de la Terre, 53 (4): 1-21. Bruxelles. [ Links ]
36. Burmann, G. 1968. Diacrodien aus dem unteren Ordovozium. Palöntol. Abhand. Abteil. B 2 (4): 635-652. [ Links ]
37. Burmann, G. 1970. Weitere organische Mikrofossilien aus dem unteren Ordovizium. Paläont. Abh., Abt. B, Paläobot., 3 (3-4): 289-332. Berlin. [ Links ]
38. Castillo, A.L. 1999. Depósitos de baritina de Cordillera Oriental, Salta. En: Recursos Minerales de la República Argentina (Ed. E.O. Zappettini), Instituto de Geología y Recurso Minerales SEGEMAR, Anales 35:499-503. Buenos Aires. [ Links ]
39.Castillo, A.L., Bataglia, R.R. y Moya, M.C. 1996. Depósitos Minerales en los Distritos Santa Victoria, Zenta, e Iruya (Precámbrico - Paleozoico Inferior), Salta, Argentina. XII Congreso Geológico de Bolivia. Tarija, Memorias. [ Links ]
40. Christensen, T. 1962. Alger. In: Böcher, T. W., Lange, M. & Sørensen, T. Botanik, Bd. 2 (Systematisk Botanik), 2, 172pp. [ Links ]
41. Cocchio, A.M. 1982. Donées nouvelles sur les Acritarches du Trémadoc et de l'Arenig Dans le massif de Mouthoumet (Corbières, France). Rev. Micropaléont. 25 (1): 26-39. Paris. [ Links ]
42. Combaz, A. 1967. Un microbios du Trémadocien dans une sondage d'Hassi Messaoud. Act. Soc. Linn. Bordeaux, B 29: 1- 26. [ Links ]
43. Cookson, I.C. y Eisenack, A. 1958. Microplankton from Australian and New Guinea Upper Mesozoic sediments. Proc. R. Soc. Victoria, 70:19-79. [ Links ]
44. Cooper, A.H., Fortey, N.J., Hughes, R.A., Molyneux, S.G., Moore, R.M., Rushton, A.W.A. y Stone, P. 2004. The Skyddaw Group of the English Lake District. Memoir of the British Geological Survey (England & Wales), parts of sheets 22, 23, 24, 28, 29, 30, 31 y 48. The stratigraphy, correlation, provenance and palaeogeography of the Skiddaw Group (Ordovician) in the English Lake District. Geological Magazine 132 (2): 185-211. [ Links ]
45. Cooper, C. y Molyneux, S.G. 1990. The age and correlation of Skiddaw Group (early Ordovician) sedimentes in the Cross Fell inlier (northern England). Geological Magazine 127: 147-157. [ Links ]
46. Cramer, F.H. 1964. Microplankton from three Palaeozoic formations in the province of Leon, NW-Spain. LeidseGeologische Mededelingen ., 30, 253-361. [ Links ]
47. Cramer, F.A. 1970. Acritarchs from the Lower Silurian Neahga Formation, Niagara Peninsula, North America. Can.J. Earth Sci ., 7 (4): 1077-1085. Ottawa. [ Links ]
48. Cramer, F.H., Allan, B., Kanes, W.H. y Díez, M.d.C. 1974. Upper Arenigian to Lower Llanvirnian Acritarchs from the subsurface of the Tadla Basin in Morocco. Palaeontolographica (B), 145 (5/6): 182-190. [ Links ]
49. Cramer, F.H. y Díez, M. 1968. Consideraciones taxonómicas sobre las acritarcas del Silúrico Medio y Superior del Norte de España. Instituto Geológico y Minero de España, Boletín 79 (6): 541-574. [ Links ]
50. Cramer, F.H. y Díez, M.d.C. 1972.North American Silurian palynofacies and their spatial arrangement: acritarchs. Palaeontographica Abteilung B Palaophytologie 138 (5-6): 107 180. [ Links ]
51. Cramer, F.H. y Díez, M. 1976. Seven new Late Arenigian species of the acritarch genus Coryphidium Vavrdová, 1972. Paläont. Z. , 50 (3-4): 201-208. [ Links ]
52.Cramer, F.H. y Díez, M. 1977. Late Arenigian (Ordovician) acritarchs from Cis-Saharan Morocco. Micropaleontology, 23 (3): 339-360. [ Links ]
53. Dean, W.T. y Martin, F. 1978. Lower Ordovician acritarchs and trilobites form Bell Island, eastern Newfounland. Bull. Geol. Surv. Canada , 284: 1-35. [ Links ]
54. Deflandre, G. 1946. Le problème des Hystrichosphères. Bulletin de l' Institut océanographique, 918: 1-23. Mónaco. [ Links ]
55. Deflandre, G. y Deflandre-Rigaud, M. 1962. Nomenclature et systématique des histrichosphères (sens. lat.). Observations et rectifications. Review Micropaléontologie, 4 (4): 190-196. [ Links ]
56. De la Puente, G.S. y Rubinstein, C.V. 2009. Late Tremadocian chitinozoans and acritarchs from Northwestern Argentina (Western Gondwana) Review of Palaeobotany and Palynology, 154: 65-78. [ Links ]
57. De la Puente, G.S., Rubinstein, C.V. y Toro, B.B. 2009. Nuevos registros de quitinozoos, acritarcos y graptolitos del área de Pascha-Incamayo, Cordillera Oriental, Argentina. XIV Simposio Argentino de Paleobotánica y Palinología, Mar del Plata. Resúmenes, p. 1. [ Links ]
58. Deunff, J. 1951. Sur la présence de micro-organismes (Hystrichosphères) dans les schistes ordoviciens du Finistère. Compte Rendu Hebdomadaire des Séances de l'Académie des Sciences 233: 321- 323. [ Links ]
59. Deunff, J. 1954. Veryhachium, genre Nouveau d' Hystrichosphères du Primaire. C. r. Soc. géol. France, Paris, 13: 305- 307. [ Links ]
60. Deunff, J. 1959. Microorganismes planctoniques du Primaire armoricain. I. Ordovicien du Veryhach (presquîle de Crozon). Bulletin de la Société géologique et minéralogique de Bretagne, 2: 1 41. [ Links ]
61. Deunff, J. 1961.Un microplancton a hystrichosphères dans le Tremadoc du Sahara. Revue de Micropaléontologie, 4 (1): 37-52. [ Links ]
62. Deunff, J. 1964. Systématique de microplancton fossile à Acritarches ; révision de deux genres de l'Ordovicien inférieur. Revue de micropaléontologie, 7 (2): 119-124. [ Links ]
63. Deunff, J., Górka, H. y Rauscher, R. 1974. Observations nouvelles et precisions sur les Acritarches à large ouverture polaire du Paléozoïque inférieur. Géobios, 7(1): 5-18. [ Links ]
64. Di Pasquo, M. y Vergel, M.d.M. 2008. Primer registro palinológico del Pennsylvaniano del Norte de la Sierra de Zenta, Provincia de Jujuy, Argentina. XII Simpósio de Paleobotânicos e Palinólogos. Florianópolis, SC, Brasil. [ Links ]
65. Dorning, K.J. 1994. Dactylofusa, Eupoikilofusa, Leiofusa and Poikilofusa: four fusiform acritarch genera. Contributions to the CIMP Symposium on Palynology, Palaeoenvironments and Stratigraphy, University of Sheffield, 15. [ Links ]
66. Downie, C. 1958. An assemblagge of microplankton from the Shineton Shales (Tremadocian). Proceedings of theYorkshire Geological Society , 31 (4): 331-349. [ Links ]
67. Downie, C. 1959. Hystrichospheres from the Silurian Wenlock Shale of England. Paleontology, 2 (1): 56-71. [ Links ]
68. Downie, C. y Sarjeant, W.A.S. 1963. On the interpretation and status of some hystrichosphere genera. Palaeontology, 6: 83-96. [ Links ]
69. Eisenack, A. 1934. Neue Mikrofossilien des baltischen Silurs III und neue Mikrofossilien des böhmischen Silurs I. Paläont. Zeit. 16 (1-2) : 52-72. [ Links ]
70. Eisenack, A. 1938. Hystricosphaerideen und verwandte Formen im baltischen Silur.Z. Geschiebeforsch., 14 (1): 1-30. [ Links ]
71. Eisenack, A. 1958. Mikroplankton aus dem norddeutschen Apt nebst einigen Bemerkungen über fossile Dinoflagellaten. N. Jb. Geol. Paläont. Abh., 106 (3): 383-422. [ Links ]
72. Eisenack, A. 1959. Neotypen baltischer Silur-Hystrichosphären ind neue Arten. Palaeontographica A 112 (5-6): 193- 211. [ Links ]
73. Eisenack, A. 1969. Zur Systematik einiger paläozoischer Hystrichosphären (Acritarchs) des baltischen Gebietes. N. Jb. Geol. Paläont . 133 (3): 245-266. [ Links ]
74. Eisenack, A. 1972. Kritische beinerkung zur Gattung Pterospermopsis (Chlorophyta, Prasinophyceae). NeusJahrbuch fur Geologic und Palaöntlogie , Monatsshefte, 10, 596-601. [ Links ]
75. Eisenack, A., Cramer, F.H. y Diez Rodríguez, M.D.C. 1973. Katalog der fossilen Dinoflagellaten, Hystrichosphären undverwandten Mikrofossilien. Band III, Acritarcha, Teil 1. Stuttgart. 1104 pgs. [ Links ]
76. Eisenack, A., Cramer, F.H. y Díez, M.d.C. 1976. Katalog der fossilen Dinoflagellaten, Hystrichosphären und verwandtenMikrofossilien. Band IV Acritarcha 2. Teil, E. Schweizerbart´sche Verlagsbuchhandlung, 863 pp. Stuttgart. [ Links ]
77. Evitt, W.R. 1963. A discussion and proposals concerning fossil dinoflagellates, hystrichospheres, and acritarchs, II. Proceedings of the National Academy of Sciences 49: 298-302. [ Links ]
78. Fatka, O. 1992. Early Arenig acritarchs from lower Klabava Formation (Prague Basin, Czechoslovakia). Vest. Čsk.Geol. Ust., 67: 277-287. [ Links ]
79. Fatka, O. 1993. Chitinozoans and acritarchs in latest Tremadoc-early Arenig sediments of the Prague Basin, Czech Republic. En: Molyneux, S.G.; Dorning, K.J. (Eds.): Contributions to Acritarch and Chitinozoan Research.Spec. Pap.Palaeontology ,48: 29-36. [ Links ]
80. Fensome, R.A., Williams, G.L., Bars, M.S., Freeman, J.M. y Hill, J.M. 1990. Acritarchs and fossil prasinophytes: an index to genera, species and intraspecific taxa. American Association of Stratigraphic Palynologists Foundation. Contributions Series, 25, 771 pp. [ Links ]
81. Fombella, M.A. 1977. Acritarcos de la Formación Oville, edad Cámbrico Medio - Tremadociano, Provincia de León, España. Palinología, Número Extraordinario 1: 245-261. [ Links ]
82. Fortey, R.A., Basset, M.G., Harper, D.A.T., Hughes, R.A., Ingham, J.K., Molyneux, S.G., Owen, R.M., Rushton, A.W.A., Sheldon, P.R. 1991. Progress and problems in the selection of stratotypes for the bases of series in the Ordovician System of the historical type area in the U.K. In Advances in Ordovician Geology (Eds. C.R. Barnes and S.H. Wiliams), pp. 5-25. Geological Survey of Canada, Paper No. 90-0. [ Links ]
83. Fortey, R.A. y Shergold, J.H. 1984. Early Ordovician trilobites, Nora Formation, central Australia. Palaeontology, 27: 315-366. [ Links ]
84. Ghavidel-Syooki, M. y Vecoli, M. 2008. Palynostratigraphy of Middle Cambrian to lowermost Ordovician stratal sequences in the High Zagros Mountains, southern Iran: Regional stratigraphic implications, and palaeobiogeographic significance. Review of Palaeobotany and Palynology,150:97-114. [ Links ]
85. González M.A., Tchilinguirian, P., Pereyra, F., Ramallo, E. y González, O.E. 2004. Hoja Geológica 2366-IV Ciudad de Libertador General San Martin, Provincias de Jujuy y Salta. (1:250.000). Boletín N° 274 SEGEMAR, IGRM. 132 pp. Edición 2004. [ Links ]
86. Górka, H. 1967. Quelques nouveaux Acritarches des silexites du Trémadocien supérieur de la region de Kielce (Montagne de Ste Croix, Pologne). Cahiers de Micropaléontologie Série I (6, 441), 8 pp. [ Links ]
87. Groeber, P. 1938. Mineralogía y Geología. Espasa-Calpe Argentina, 1-492, Buenos Aires. [ Links ]
88. Heredia, S. y Aceñolaza, G.F. 2005. The Trapezognathus diprion conodont zone and correlation o fan outstanding ocurrente of the Cruzianarugosa group (trace fossils) in the Lower Ordovician of western Gondwana. En: Pankurst, R.J. y Veiga, R. (Eds.) Gondwana 12: Geological and biological heritage of Gondwana. Academia Nacional de Ciencias: 197. [ Links ]
89. Heredia, S., Carlorosi, J., Aceñolaza, G. y Aceñolaza, F. 2008. El Género Trapezognathus (Conodonta) en el Ordovícico de la Cordillera Oriental: Implicancias Estratigráficas y Bioestratigráficas. XVII Congreso Geológico Argentino, Jujuy. Actas I: 357. [ Links ]
90. Hill, P. J. y Molyneux, S.G. 1988. Biostratigraphy, palynofacies and provincialism of Late Ordivician Early Silurian acritarchs from northeast Libya. 27-43. En: El-Arnauti, A.; Owens, B.; Thusu, B. (Eds.): Subsurfacepalynostratigraphy of northeast Libya . Garyounis University Publications, Benghazy, 276 pp. [ Links ]
91. Jachowicz, M. 1995. Ordovician acritarch assemblages from Central and Northwestern Saudi Arabia. Review ofPalaeobotany and Palynology , 89: 19-25. [ Links ]
92. Jacobson, S.R. 1978. Acritarchs from the Upper Ordovician Clays Ferry Formation, Kentucky, U.S.A. Palinología, Número Extraordinario 1: 293-301. [ Links ]
93. Jacobson, S.R. y Achab, A. 1985. Acritarch biostratigraphy of the Dicellograptus complanatus graptolite zone from the Vaureal formation (Ashgillian), Anticosti Island, Quebec, Canada. Palynology, 9: 165-198. [ Links ]
94. Jardiné, S., Combaz, A., Magloire, L., Peniguel, G. y Vachey, G. 1974. Distribution stratigraphique des acritarches dans le Paleozoique du Sahara Algerien. Review of Palaeobotany and Palynology, 18: 99-129. [ Links ]
95. Keidel, J. 1925. Sobre el desarrollo paleogeográfico de las grandes unidades geológicas de la República Argentina.Anales de la Sociedad Argentina de estudios Geográficos GAEA 4. [ Links ]
96. Konzalová-Mazancová, M. 1969. Acritarcha Evitt 1963 aus dem Unter-Asgill Böhmens. Palaeontographica, AbteilungB , 165: 81-92. [ Links ]
97. Le Hérissé, A. 1988. Acritarches et kystes d'algues Prasinophycées du Silurien de Gotland, Suède. Palaeontographicaitalica , 76: 57-302. Pisa. [ Links ]
98. Li, J. 1987. Ordovician acritarchs from the Meitian Formation of Guizhou province, south-west China. Palaeontology, 30 (3): 613-634. [ Links ]
99. Li, J., Servais, T. y Brocke, R. 2002. Chinese Paleozoic acritarch research: review and perspectives. Review ofPalaeobotany and Palynology , 118: 181-193. [ Links ]
100. Lister, T.R. 1970. The acritarchs and chitinozoa from the Wenlock and Ludlow Series of the Ludlow and Millichope areas, Shropshire. Palaeontographical Soc., Monogr. 528, 124 (1): 100 pp. London. [ Links ]
101. Loeblich, A.R. y Tappan, H. 1976. Some new and revised organic-walled phytoplancton microfossil genera. Journal of Paleontology 50: 301-308. [ Links ]
102. Loeblich, A.R. y Tappan, H. 1978. Some middle and late ordovician microphytoplankton from Central North America. Journal of Paleontology, 52 (6): 1233-1287. [ Links ]
103. Lu, L. 1987. Acritarchs from the Dawan Formation (Arenigian) of Huanghuachang in Yichang, western Hubei. Acta Micropaleontologica Sinica 4 (1): 87-102. [ Links ]
104. Manca, N., Heredia, S., Hunicken, M. y Rubinstein, C., 1995. Macrofauna, conodontes y acritarcos de la Formación Santa Rosita (Tremadociano), Nazareno, provincia de Salta, Argentina. Boletín de la Academia Nacional de Ciencias, Córdoba, Argentina 60 (3-4): 267-275. [ Links ]
105. Martin, F. 1969. Les acritarches de L'Ordovicien et du Silurien Belges, Détermination et Valeur stratigraphique. Memoires de l' Institut Royal des Sciences Naturelles de Belgique , 160 : 1-175. Bruxelles. [ Links ]
106. Martin, F. 1972. Les acritarchs de l'Ordovicien inferieur de la Montagne Noire (Herault, France). Bulletin Instituteroyal des Sciences naturelles de Belgique , 48 (10): 1 61. [ Links ]
107. Martin, F. 1977. Acritarches de Cambro-Ordovicien du Massif de Brabant, Belgique. Institute royal des Sciencesnaturelles de Belgique, Sciences de la terre, Bulletin 51: 1 33. [ Links ]
108. Martin, F. 1982. Some aspects of late Cambrian and early Ordovician acritarchs. En: Basset, M.G.; Dean, W.T. (Eds.): The Cambrian-Ordovician boundary: sections, fossil distributions, and correlations 29-40. National Museum of Wales, Geological Series 3. Cardiff. [ Links ]
109. Martin, F. 1993. Acritarchs: a review. Biological Reviews, 68(4): 475-538. [ Links ]
110. Martin, F. 1996. Systematic revision of the acritarch Ferromia pellita and its bearing on Lower Ordovician stratigraphy. Review of Palaeobotany and Palynology 93: 23-34. [ Links ]
111. Martin, F. y Dean, W.T. 1981. Middle and Upper Cambrian and Lower Ordovician acritarchs from Random Island, eastern Newfoundland. Geological Survey of Canada, Bulletin, 343: 1 43. [ Links ]
112. Martin. F. y Dean, W.T. 1988. Middle and Upper Cambrian acritarch and trilobite zonation at Manuels River and Random Island, eastern Newfoundland. Geological Survey Canada Bulletin,381: 1-99. [ Links ]
113. Martin, F., Michot, J. y Vanguestaine, M. 1970. Le flysch caradocien d'Ombert. Annales de la Société Géologique deBelgique , 93 : 337-362. [ Links ]
114. Mette, W. 1989. Acritarchs from lower Paleozoic rocks of the western Sierra Morena, SW-Spain and biostratigraphic results. Geologica et Palaeontologica, 23: 1-19. [ Links ]
115. Moczydlowska, M. y Stockfors, M. 2004. Acritarchs from the Cambrian-Ordovician boundary interval on Kolguev Island, Arctic Russia. Palynology, 28: 15-73. [ Links ]
116. Moczydlowska, M. y Vidal, G. 1986. Lower Cambrian acritarch zonation in southern Scandinavia and southeastern Poland. Geologiska Föreningens i Stockholm Förhandlingar, 108 (3): 201-223. [ Links ]
117. Molyneux, S.G. 1987. Appendix. Acritarchs and chitinozoa from the Arenig Series of south - west Wales. Bulletinof the British Museum , 41 (3): 309-364. [ Links ]
118. Molyneux, S.G. y Dorning, K.J. 1989. Acritarch dating of latest Tremadoc- earliest Arenig (early Ordovician) sediments in the Carmarthen district, south Wales. Geological Magazine, 126 (6): 707-714. [ Links ]
119. Molyneux, S.G. y Rushton, A.W.A. 1988. The age of the Watch Hill Grits (Ordovician), English Lake District: structural and palaeogeographical implications. Trans. Roy. Soc. Edinburgh Earth Sci., 79: 43-69. [ Links ]
120. Monaldi, C.R., Boso, M.A. y Fernández, J.C. 1986. Estratigrafía del ordovícico de la Sierra de Zapla, provincia de Jujuy. Revista de la Asociación Geológica Argentina, 41 (1-2): 62-69. [ Links ]
121. Nieniewski, A. y Wleklinski, E. 1950. Contribución al conocimiento del anticlinal de Zapla (provincia de Jujuy). Revista de la Asociación Geológica Argentina , 5 (4): 169-203. Bs. As. [ Links ]
122. Nieva, S.M. y Aceñolaza, G.F. 2006. Estructuras cono en cono en los estratos del Ordovícico Inferior de la Sierra de Zenta, Cordillera Oriental de Jujuy (Argentina). XVII Congreso Geológico Boliviano, Memorias: 59-62. Sucre. [ Links ]
123. Noetinger, S. y Di Pasquo, M. 2008. First record of a Devonian palynological assemblage from the Zenta Range, Eastern Cordillera, Northwestern Argentina. XII Simpósio de Paleobotânicos e Palinólogos. Florianopólis, SC, Brasil. [ Links ]
124. Ortega, G., Monaldi, C.R., Albanesi, G.L. y Zeballo, F.J. 2007. Las zonas de Araneograptus murrayi y Hunnegraptus copiosus (Tremadociano tardío) en la Sierra de Zenta, Cordillera Oriental, noroeste argentino. Ameghiniana Suplemento 44 (4), Resumenes, 97 R. [ Links ]
125. Ottone, J.C. 1999. Bioestratigrafía - Palinología. En: González Bonorino, G.; Omarini, R.; Viramonte, J. (Eds.): Geología del Noroeste Argentino, XIV Congreso Geológico Argentino, Salta, Relatorio I, 223-224. [ Links ]
126. Ottone, J., Toro, B. y Waisfeld, B. 1992. Lower ordovician palynomorphs from the Acoite formation, Northwestern Argentina. Palynology, 16: 93-116. [ Links ]
127. Ottone, E.G., Waisfeld, B.G. y Astini, R.A. 1995. Acritarcas del Ordovícico Temprano de la Quebrada de Chalala, Noroeste de argentina. Ameghiniana, 32 (3): 237-242. [ Links ]
128. Pascher, A. 1914. Über Flagellaten und Algen. Berichte der deutsche botanischen Gesellschaft 32: 136 160. [ Links ]
129. Playford, G. y Martin, F. 1984. Ordovician acritarchs from the Canning Basin, Western Australia. Alcheringa 8: 187 223. [ Links ]
130. Playford. G. y Wicander, R. 1988. Acritarch palynoflora of the Coolibah Formation (Lower Ordovician), Georgina Basin, Queensland.En: Jell, P.A.; Playford, G. (Eds.): Palynological and palaeobotanical studies in honour of Basil E. Balme. Mem. Ass. Australas. Palaeontolsts. 5: 5-40. [ Links ]
131. Quintavalle, M., Tongiorgi, M. y Gaetani, M. 2000. Lower to middle ordovician acritarchs and chitinozoans from northern Kakakorum Mountains, Pakistan. Rivista Italiana di Paleontologia e Stratigrafia, 106 (1): 3-18. [ Links ]
132. Ramos, V. A. 1999. Las provincias geológicas del territorio argentino. En: Caminos, R. (Ed.) Geología Argentina. Anales 29 (3): 41-96. Instituto de Geología y Recursos Minerales, Buenos Aires. [ Links ]
133. Ramos, V.A. y Coira, B. 2008. Las Provincias Geológicas de Jujuy. XVII Congreso Geológico Argentino, Jujuy. Relatorio 2: Geología y Metalogénesis: 11-16. [ Links ]
134. Rasul, S.; 1976: New species of the genus Vulcanisphaera (Acritarcha) from the Tremadocian of England. Micropaleontology 22 (4): 479-484. [ Links ]
135. Rasul, S. 1979. Acritarch zonation of the tremadocian series of the Shineton Shales, Wrekin, Shropshire, England. Palynology , 3: 53-72. [ Links ]
136. Rasul, S.M. y Downie, C. 1974. The stratigraphic distribution of Tremadoc acritarchs in the Shineton Shales succession, Shropshire, England. Review of Palaeobotany and Palynology, 18:1 9. [ Links ]
137. Rubinstein, C.V. 1997. Tremadocian acritarchs from northwestern Argentina. Review of Palaeobotany and Palynology, 98: 41-45. [ Links ]
138. Rubinstein, C.V. 2001. Correlaciones entre las cuencas ordovícicas y silúricas de la Argentina basadas en el estudio de acritarcos, criptoesporas y mioesporas. Correlacao de secuencias paleozoicas sul-americanas (J.H.G. Melo y G.J.S. Terra, Eds.) Ciencia Técnica-Petróleo 20. [ Links ]
139. Rubinstein, C.V. 2003. Ordovician acritarchs from the northwestern Argentina: new insights into the biostratigraphy andpaleoenvironmental aspects of the Central Andean Basin and Famatina. En: [ Links ]
140. Rubinstein; C.V. 2005. Ordovician to lower Silurian palynomorphs from the Sierras Subandinas (Subandean ranges), northwestern Argentina: a preliminary report. Carnets de Geologie, Memoir 2005/2, Abstract 09. [ Links ]
141. Rubinstein, C.V. y Brussa, E. 1999. A palynomorph and graptolite biostratigraphy of the Central Precordillera Silurian Basin, Argentina. Bollettino della Società Paleontologica Italiana,38 (2-3): 257-266. [ Links ]
142. Rubinstein, C.V. y de la Puente, S. 2008. Bioestratigrafía del Paleozoico Inferior en las Sierras Subandinas, Provincia de Jujuy. XVII Congreso Geológico Argentino, Jujuy. Relatorio 2: 128-133. [ Links ]
143. Rubinstein, C.V., de la Puente, G.S., Delabroye, A. y Astini, R. 2008. New palynological record of the Ordovician / Silurian Boundary in the Argentine Cordillera Oriental, Central Andean Basin. Terra Nostra 2008/2 IPC-XII IOPC-VIII Bonn, Alemania. Abstracts: 592. [ Links ]
144. Rubinstein, C.V., de la Puente, G.S., Servais, T., Vecoli, M. y Astini, R.A. 2007a. Advances in the Ordovician palynology of Argentina: biostratigraphy and paleobiogeography.En: Díaz-Martinez, E.; Rábano, I. (Eds.) 4th European Meeting on the Palaeontology and Stratigraphy of Latin America Cuadernos del Museo Geominero, N°8. Instituto Geologico y Minero de España. Madrid. 349-354. [ Links ]
145. Rubinstein, C.V., de la Puente, G.S., Toro, B.A. y Servais, T. 2007b. The presence of the messaoudensis trifidum acritarch assemblage (Upper Tremadocian Floian) in the Central Andean Basin, north-western Argentina:calibration with chitinozoans and graptolite zonation. Acta Palaeontologica Sinica, 46 (suppl.): 422-428. [ Links ]
146. Rubinstein, C.V. y Toro, B.A. 1999. Acritarch and graptolite biostratigraphy in the Lower Arenig of the peri- Gondwana related Eastern Cordillera, Argentina. Acta Universitatis Carolinae Geológica, 43 (1/2): 255-258. [ Links ]
147. Rubinstein, C.V. y Toro, B.A. 2001. Review of acritarch biostratigraphy in the Arenig of the Eastern Cordillera, Northwestern Argentina: new data and calibration with the graptolite zonation. Contributions to geology andpalaeontology of Gondwana in Honour of Helmut Wopfner . 421-439. Colonia. [ Links ]
148. Rubinstein, C.V. y Toro, B. 2002. The messaoudensis trifidum acritarch assemblage (late tremadoc early arenig) from Parcha, Eastern Cordillera, northwestern Argentina. CIMP International Meeting and Workshops, Lille, France. Paleozoic Palynology in the Third Millenium: new directions in acritarch, chitinozoan and miospore research. Abstract: 48. [ Links ]
149. Rubinstein, C.V., Toro, B.A. y Waisfeld, B.G. 1999. Acritarch biostratigraphy of the upper Tremadoc - Arenig of the Eastern Cordillera, northwestern Argentina: relationships with graptolite and trilobite faunas. Bollettino dellaSocieta Paleontologica Italiana , 38 (2-3): 267-286. [ Links ]
150. Rubinstein, C.V. y Vaccari, N.E. 2001. Palynomorphs of the Ordovician Silurian boundary in the Salar del Rincón Formation, Argentine Puna. Firts Meeting of the C.I.M.P. Spores and Pollen Subcommission, National University of Ireland, Cork; Ireland. Abstracts, pp. 40-41. [ Links ]
151. Rubinstein, C.V. y Vaccari, N.E. 2004. Cryptospore assemblages from the Ordovician-Silurian boundary in the Puna, North - West Argentina. Palaeontology,47: 1037-1061. [ Links ]
152. Salfity, J.A. y Monaldi, C.R. 1998. Mapa Geológico de la Provincia de Salta, Escala 1:500.000. Segemar - Servicio GeológicoMinero de la Argentina . Buenos Aires. [ Links ]
153. Sánchez, M.C. y Salfity, J.A. 1999.La cuenca cámbrica del Grupo Mesón en el Noroeste Argentino: desarrollo estratigráfico y paleogeográfico. Acta Geológica Hispánica, 34 (2-3): 123-139. [ Links ]
154. Schiller, J. 1925. Die planktonischen Vegetationen des Adriatischen Meeres. B. Chrysomonadina, Heterokontae, Cryptomonadina, Eugleninae, Volvocales. 1. Systematischer Teil. (Nach den Ergebnissen der österreichischen Adriaforschung in den Jahren 1911-1914). Archiv für Protistenkunde 53: 59-123. [ Links ]
155. Servais, T. 1993. A critical review of some Ordovician acritarch taxa and their stratigraphical implications in Belgium and Germany. Unpublished PhD Thersis, University of Liège: 1-327. [ Links ]
156. Servais, T., Li, J., Molyneux, S. y Vecoli, M. 2008. The Ordovician acritarch genus Coryphidium. Revue de micropaleontology , 51: 97-120. [ Links ]
157. Servais, T., Li, J., Stricanne, L., Vecoli, M. y Wicander, R. 2004. Acritarchs. En: Webby, B.; Droser, M.; Paris, F.; Percival, I. (Eds): The Great Ordovician Biodiversification Event. Columbia University Press, New York, pp. 348-360. [ Links ]
158. Servais, T., Li, J. y Yan, K. 2008. Acritarch biostratigraphy of the Lower - Middle Ordovician boundary: advances and problems. Terra Nostra, IPC-XII/IOPC-VII. Alemania [ Links ]
159. Servais, T. y Maletz, J. 1993. Lower Llanvirn (Ordovician) graptolites and acritarchs from the "Assise de Huy", Bande de Samre-et-Meuse, Belgium. Annales de la Société géologique de Belgique, 115: 265-285. [ Links ]
160. Servais, T. y Molyneux, S. 1997. The messaoudensis - trifidum acritarch assemblage (Ordovician: late Tremadoc - early Arenig) from the subsurface of Rugen (Baltic Sea, NE Germany). Palaeontographia Italica, 84: 113-161. [ Links ]
161. Slavíková, K. 1968. New finds of acritarchs in the Middle Cambrian of the Barradian (Czechoslovakia). Vest. Ú.ú. G. 43: 199-205. [ Links ]
162. Soufiane, A. y Achab, A. 1993. Quelques assemblages de Chitinozoaires de l'Ordovicien du Maroc. Bassin de Tadla. Geobios, 26, 535 553. [ Links ]
163. Staplin, F.L. 1961. Reef-controlled distribution of Devonian microplankton in Alberta. Palaeontology, 4: 392 424. [ Links ]
164. Staplin, F.L., Jansonius, J. y Pocock, S.A.J. 1965. Evaluation of some Acritarchous Hystrichosphere Genera. NeuesJarbuch für Geologie und Paläontologie . Abhandlungen, 123 (2): 167-201. [ Links ]
165. Starck, D. 1995. Silurian - Jurassic Stratigraphy and Basin Evolution of Northwestern Argentina. En: A. J., Tankard; R., Suarez S.; and H.J., Welsink; Petroleum basins of South America: AAPG Memoir, 62: 251-267. [ Links ]
166. Starck, D. 1996. Facies continentales en el siluro - devónico de la cuenca del Noroeste. Provincia de Salta, Argentina. XII Congreso Geológico de Bolivia, Tarija, Memorias 231-238. [ Links ]
167. Starck, D. 2008. La cuenca carbonífera - ¿eomesozoica? En la provincia de Jujuy, su estratigrafía y evolución tectosedimentaria. XVII Congreso Geológico Argentino, Relatorio, 2: 199-206. [ Links ]
168. Starck, D., Gallardo, E. y Schulz, E. 1992a. La discordancia precarbónica en la porción argentina de la Cuenca de Tarija. Boletín de Informaciones petroleras, Buenos Aires, Argentina, Tercera Epoca, Año IX, 29: 2-11. [ Links ]
169. Starck, D.; Gallardo; E.; Schulz, E.; 1992b: La Cuenca de Tarija: Estratigrafía de la porción argentina. Boletín de Informaciones petroleras, Bs. As., Argentina, 30: 2-14. [ Links ]
170. Starck, D., Gallardo, E. y Schulz, E. 1993a. The pre-Carboniferous unconformity in the Argentine portion of the Tarija Basin. XII International Congress on Carboniferous and Permian. Buenos Aires 2: 373-384. [ Links ]
171. Starck, D., Gallardo, E. y Schulz, E. 1993b. Neopaleozoic stratigraphy of the Sierras Subandinas Occidentales and Cordillera Oriental Argentina. XII International Congress on Carboniferous and Permian. Buenos Aires 2: 353-372. [ Links ]
172. Stockmans, F.; Willière, Y.; 1962: Hystrichospheres du Dévonien belge (sondage de l'Asile d' alienes a Tournai). Bulletin de la Société Belge Géologique, Paléontologie et Hydrologie 71: 41 - 77. [ Links ]
173. Sureda, R.J. 1999. Los yacimientos sedex de plomo y zinc en la Sierra de Aguilar, Jujuy. En: Recursos Minerales de laRepública Argentina (Ed. E.O. Zappettini), Instituto de Geología y Recurso Minerales SEGEMAR, Anales 35: 499-503. Buenos Aires. [ Links ]
174. Timofeev, B.V. 1958. Über das Alter sächsischer Grauwacken. Mikropaläontologische Untersuchungen von Proben aus der Weesensteiner und Lausitzer Grauwacke. Geologie 7 (3-6): 826-845. [ Links ]
175. Timofeev, B.V. 1959. The ancient flora of the Prebaltic and its stratigraphic importance. Vsesoyuznyi NeftyanoiNauchno-Issledovatelskii Geologroazvedochnyi Institut, Leningrad Vnigri , Trudy 129: 1-136. [ Links ]
176. Tongiorgi, M., Di Milia, A., Le Fort, P. y Gaetani, M. 1994. Palynological dating (Arenig) of the sedimentary sequence overlying the Ishkarwaz Granite (upper Yarkhun valley, Chitral, Pakistan). Terra Nova, 6: 595-607. [ Links ]
177. Tongiorgi, M., Yin, L.M. y Di Milia, A. 1995.Arenigian Acritarchs from the Daping Section (Yangtze Gorges area, Hubei Province, Southern China) and their palaeogeographic significance. Review of Paleobotany and Palynology, 86 (1-2): 13-48. [ Links ]
178. Tongiorgi, M., Yin, L. y Di Milia, A. 2003. Lower Yushanian to lower Zhejiangian palynology of the Yangtze Gorges area (Daping and Huanghuachang sections), Hubei Province, South China. Palaeontographica, Abt. B, 266: 1-160. [ Links ]
179. Toro, B.A., Rubinstein, C.V., Waisfeld, B.G., Astini, R.A. 2003. Calibración de las asociaciones de palinomorfos y trilobites de las zonas de graptolitos del Ordovícico temprano del área de Parcha, Cordillera Oriental Argentina. Ameghiniana. Resúmenes 40 (4). Suplemento: 94-95R. [ Links ]
180. Tortello, M.F., Aceñolaza, G.F. 2008. Los trilobites Tremadocianos del Abra de Zenta (Sistema Interandino, provincias de Jujuy y Salta). XVII Congreso Geológico Argentino, Jujuy. Actas, I: 375. [ Links ]
181. Tortello, M.F., Esteban, S.B., Aceñolaza, G.F., Aráoz, L., Powell, J.E., Carrizo, H. y Azcuy, C. 2008. Paleontología y bioestratigrafía de la Provincia de Jujuy. XVII Congreso Geológico Argentino, Jujuy. Relatorio: 589-405. [ Links ]
182. Turner, J.C.M. 1959. Estratigrafía del cordón de Escaya y de la sierra de Rinconada (Jujuy). Revista de la AsociaciónGeológica Argentina , 13 (1-2): 15-39. Buenos Aires. [ Links ]
183. Turner, J.C. 1960. Estratigrafía de la Sierra de Santa Victoria y adyacencias. Boletín de la Academia Nacional de Ciencias, 41 (2): 163-196. Córdoba. [ Links ]
184. Turner, J.C. 1964. Descripción geológica de la hoja 2c, Santa Victoria (provincias de Salta y Jujuy). Instituto Nacional de Geología y Minería, Boletín 104, 1-83. [ Links ]
185. Turner, J.C. y Mon, R. 1979. Cordillera Oriental. En: Turner, J.C. (Ed.): Geología Regional Argentina. Academia Nacional de Ciencias de Córdoba 1: 57-94. [ Links ]
186. Turner, R.E. 1984. Acritarchs from the type área of the Ordovician Caradoc Series, Shropshire, England. Palaeontographica, Abt. B. , 190 (4-6): 87-157. Sttuttgart. [ Links ]
187. Turner, R.E. 1985. Acritarchs from the type area of the Ordovician Llandeilo Series, South Wales. Palynology, 9: 211 234. [ Links ]
188. Tynni, R. 1975. Ordovician hystrichospheres and chitinozoans in limestone from the Bothnian Sea. GeologicalSurvey of Finland Bulletin 279: 1 59. [ Links ]
189. Vanguestaine, M. 1978. Données nouvelles dans l'Ordovicien inférieur du bassin de la Senne, Massif du Brabant, Belgique. Ann. Soc. Géol., Belgique 100 : 193-198. [ Links ]
190. Vavrdová, M. 1966. Paleozoic microplankton from central Bohemia. Cãs. Mineral. Geol., 11 (4): 409-414. [ Links ]
191. Vavrdová, M. 1972. Acritarchs from Klabava Shales (Arenig). Vest. Ustred. Ustavu Geol., 47: 79-86. [ Links ]
192. Vavrdová, M. 1974. Geographical differentiation of Ordovician acritarch assemblages in Europe. Review of Palaeobotanyand Palynology , 18: 171-175. [ Links ]
193. Vavrdová, M. 1976. Excystment mechanism of early Palaeozoic acritarchs. Cãsopis pro mineralogii a geologii.21: 55 64. [ Links ]
194. Vavrdová, M. 1977. Acritarchs from the Sárka Formation (Llanvirnian). Vestník Ustred. ústavu geol., 52: 109-118. [ Links ]
195. Vavrdová, M. 1979. Netromorphitae and some other acritarchs from the Bohemian Lower Ordovician. En: Pokorny, V. (Ed.). paleontologická Konference Katedry Paleontologie na Prirodovedecké Fakulte Univerzity Karlovy, Praha: 61-74. [ Links ]
196. Vavrdová, M. 1982. Recicled acritarchs in the uppermost Ordovician of Bohemia. Cãs. Mineral. Geol., 27 (4): 337- 345. [ Links ]
197. Vavrdová, M. 1988. Further acritarchs and terrestrial plants remains from the late Ordovician at the Hlásnpa Treban (Czechoslovakia). Cãs. Mineral. Geol., 33 (1): 1-10. [ Links ]
198. Vavrdová, M. 1989. New acritarchs and miospores from the Late Ordovician of Hlásnpa Treban, Czechoslovakia(Nová akritarcha a miospory ze svrchního ordoviku Hlásné Trebanòe (Ceskosloslovensko). Cãs. Mineral. Geol., 34 (4): 403-419. [ Links ]
199. Vavrdová, M. 1990a. Coenobiotal acritarchs and other palynomorphs from the Arenig/Llanvirn boundary, Prague basin. Vestník Ustred. ústavu Geol., 65 (4): 237-242. [ Links ]
200. Vavrdová, M. 1990b. Early Ordovician acritarchs from the locality Mýto near Rokycany (late Arenig, Czechoslo- vakia). Cãsospis pro Mineralogii a Geologii, 35 (3): 239-250. [ Links ]
201. Vavrdová, M. 1993. Acritarch assemblages in the Arenigian Series of the Prague Basin, Czech Republic. SpecialPapers in Palaeontology , 48: 125-139. [ Links ]
202. Vecoli, M. 1999. Cambro-Ordovician palynostratigraphy (acritarchs and prasinophytes) of the Hassi-R'mel area and northern Rhadames Basin, North Africa. Palaeontographia Italica, 86: 1-112. [ Links ]
203. Vecoli, M. 2000. Palaenvironmental interpretation of microphytoplankton diversity trends in the Cambrian - Ordovician of the northen Sahara Platform. Palaeogeography, Paleoclimatology, Palaeocology, 160:329-346. [ Links ]
204. Vecoli, M., Rubinstein, C., de la Puente, S. y Servais, T. 2006. Hirnantian palynomorphs (acritarcos, chitinozoans, cryptospores)from glacial-related sedimetns of North Africa and Argentina.RTS (Reunión de Sciences de la Terre), Université de Bournogne, Dijon, France. Abstracts. [ Links ]
205. Vergel, M.d.M., Aráoz, L. y Rubinstein, C.V. 2002. Ordovician palynomorphs of Argentina: An integrated Ap- proach. En: Aceñolaza F.G: (Ed.): Aspects of the Ordovician System in Argentina. Serie de Correlación Geológica, 16: 209- 224. [ Links ]
206. Vergel, M.d.M., di Pasquo, M., Aráoz, L. y Noetinger, S. 2008a. Asociaciones palinológicas del Paleozoico en la Sierra de Zenta, Provincia de Jujuy, Argentina. XII Simposio de Paleobotânicos e Palinólogos. Florianópolis, SC. Brasil [ Links ]
207. Vergel. M.d.M., di Pasquo, M., Noetinger, S., Aráoz, L. y Aceñolaza, G.F. 2008b. Estudios palinoestratigráficos del Paleozoico en la Sierra de Zenta, Provincia de Jujuy, Argentina. II Jornadas Geológicas, Fundación Miguel Lillo. Resúmenes, 152-153. [ Links ]
208. Volkheimer, W., Melendi, D.L. y Aceñolaza, F.G. 1980.Una microflora ordovícica de la Formación Mojotoro, provincia de Salta. Revista de la Asociación Geológica Argentina, 35 (3): 401-416. [ Links ]
209. Volkova, N.A. 1990. Middle and Upper Cambrian acritarchs in the East-European Platform. Ordena TrudovogoKrasnogo Znameni Geologicheskii Institut, Trudy 454, Nauka, Moscow, pp. 1-116. [ Links ]
210. Waisfeld, B.G., Vaccari, N.E. Toro, B.A., Rubinstein, C.V. y Astini, R.A. 2006. Revisión de la Biozona de Ogygiocarisaraiorhachis (Trilobita, Tremadociano tardío) en la región de Pascha - Incamayo, Cordillera Oriental, Argentina: Parte 1: Bioestratigrafía. Ameghiniana, 43 (4): 717-728. [ Links ]
211. Wall, D. y Downie, C. 1963. Permian hystrichospheres from Britain. Palaeontology, 5: 770 784. [ Links ]
212. Webby, B.D., Paris F., Droser, M.L. y Percival, I.G. 2004. The Great Ordovician Biodiversification Event. Columbia University Press, New York, 484 p. [ Links ]
213. Welsch, M. 1986. Die Acritarchen der hoheren Digermul-Gruppe, Mittelkambriumbis bis Tremadoc, Ost-Finnmark, Nord-Norwegen. Palaeontographica Abt. B, 201 (1-4): 1 109. [ Links ]
214. Yin, L.M. 1995. Early Ordovician acritarchs from Huanjiang region, Jilin, and Yichang region, Hubei, China. Palaeontographica sinica, ser. A , 185 (12): 170. [ Links ]
215. Yin Lei-ming, Di Milia, A. y Tongiorgi, M. 1998. New and emended acritarch taxa from the lower Dawan Formation (lower Arenig, Huanghuanchang Section, South China. Review of Palaeobotany and Palynology, 102: 223- 248. [ Links ]
216. Zanettini, J.C.M. 1973. Observaciones sobre la facies proximal del subgrupo Santa Bárbara, en el sector Noroeste de la Cordillera Oriental (Provincias de Salta y Jujuy). Revista de la Asociación Geológica Argentina, 28 (3): 219-226. [ Links ]
Recibido: 10 de Noviembre de 2009
Aceptado: 23 de Diciembre de 2009

