










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkSerie correlación geológica
versión On-line ISSN 1666-9479
Ser. correl. geol. vol.32 no.1 San Miguel de Tucumán jun. 2016
ARTICULO
Bioestratigrafía, paleogeografía y paleoecología del Paleozoico de Sierra de Zenta, Cordillera Oriental Argentina
BIOSTRATIGRAPHY, PALEOGEOGRAPHY AND PALEOECOLOGY OF THE PALEOZOIC FROM ZENTA RANGES, EASTERN RANGE OF ARGENTINA.
Lucia ARÁOZ1, Sol NOETINGER2, María del Milagro VERGEL1 y Mercedes DI PASQUO3
1) Instituto Superior de Correlación Geológica (INSUGEO) CONICET - UNT - Facultad de Ciencias Naturales e IML, Miguel Lillo 205 – (4000) San Miguel de Tucumán.
2) Museo Argentino de Ciencias Naturales – CONICET, Angel Gallardo 470, Ciudad A. de Buenos Aires (C1405DJR), Argentina.
3) CICYTTP-CONICET, Laboratorio de Palinoestratigrafía y Paleobotánica, Dr. Matteri y España SN, Diamante (E3105BWA), Entre Ríos, Argentina.
Resumen: El conocimiento de los palinomorfos, incluyendo acritarcos, quitinozoos, algas prasinofíceas, criptosporas, esporas triletes y otros, del Paleozoico de la Sierra de Zenta (Cordillera Oriental Argentina) ha tenido un importante incremento en los últimos años. Este crecimiento en los estudios se refere tanto a su distribución temporal, composición de las asociaciones, relaciones con otros grupos fósiles, así como las características intrínsecas de las microforas. En este caso, el lapso de tiempo de distribución de los palinomorfos analizados abarca desde el Ordovícico temprano (Tremadociano temprano) hasta el Carbonífero tardío (Pennsylvaniano medio). El presente artículo reúne el conocimiento actual de las microforas identificadas en estratos ordovícicos, silúricos, devónicos y carboníferos de diferentes secciones de la Sierra de Zenta, con consideraciones bioestratigráficas, paleogeográficas y paleoecológicas.
Palabras clave: Palinología. Paleozoico. Paleoambientes. Noroeste de Argentina.
Abstract: The knowledge of palynomorphs, inclu-ding acritarchs, chitinozoan, prasinophytes, cryptospores, trilete spores and associated forms within the Paleozoic of the Zenta Range (Eastern Range of Argentina) display a significant increase in recent years. This increment in the studies comprises temporal distribution, composition of assemblages, relationships with other fossil groups as well as the intrinsic characteristics of the micro foras. The current study updates the knowledge on the palyno-morphs spanning the Early Ordovician (Early Tremadocian) to the Late Carboniferous (Middle Pennsylvanian) of different sections of the Zenta Range, regarding biostratigraphic, paleogeographic and paleoecological considerations.
Key words: Palynology. Paleozoic. Paleoenvironments. Northwestern Argentina.
Introducción
La Cuenca Paleozoica Andina Central cubre un amplio sector del centro - oeste sudamericano, con un desarrollo areal que abarca principalmente regiones de Perú, Bolivia y el noroeste argentino (NOA). En términos generales, está caracterizada por secuencias de edades cambro - silúricas, altamente fosilíferas y dominadas litológicamente por facies silicoclásticas. En el noroeste argentino incluye aforamientos en Puna, Cordillera Oriental y Sierras Suban-dinas, extendiéndose hacia el subsuelo de Llanura Chaqueña. Esta cuenca paleozoica, al igual que las cuencas mesozoicas superpuestas, se sitúan en áreas pericratónicas de antepais (Astini, 2003, 2008) y que fueron modificadas en su fisiografía por los efectos de la tectónica, principalmente del Ciclo Andino (Ramos, 1999).
La Sierra de Zenta es una unidad orográfica del noroeste argentino destacable, que se desarrolla en las provincias geológicas de Cordillera Oriental y Sierras Subandinas, en un área también llamada Sistema Interandino por Ramos (1999) en las provincias de Jujuy y Salta, cubriendo casi 15.000 km2. Cuenta con cumbres que llegan a los 5000 m.s.n.m., donde se destaca el desarrollo de potentes aforamientos asignados al Precámbrico, Paleozoico y Terciario (Starck, 1996, 1999; Aráoz et al., 2008; Aráoz, 2009 a). A pesar de su extensión e importancia, el conocimiento geológico de la Sierra de Zenta estuvo limitado durante mucho tiempo por la dificultad de los accesos y marcadas elevaciones, situación que paulatinamente se fue revirtiendo en la última década gracias al desarrollo de nuevos caminos.
Diferentes proyectos de investigación abordaron su estudio con el objeto de profundizar sobre la estratigrafía, paleontología y bioestratigrafía, con el fin de lograr un mayor entendimiento de los eventos tanto geológicos como biológicos ocurridos y el marco paleoambiental en el que se desarrollaron.
El estudio de los palinomorfos y microforas, particularmente, tuvo un desarrollo muy variable en el noroeste argentino, desde sus inicios en la década del 1980 (Volkheimer et al., 1980; Martin, 1982; Ottone et al., 1992, 1995; Manca et al., 1995; Rubinstein, 1997; Ottone, 1999; Rubinstein y Toro, 1999; Rubinstein et al., 1999), con una notable ausencia de trabajos para la Sierra de Zenta hasta el año 2001, cuando se realizaron los primeros relevamientos palinológicos en estratos paleozoicos con un análisis de sus implicancias bioestratigráficas y paleogeográficas (Aceñolaza et al., 2001, 2003; Araóz y Vergel, 2001; Aráoz, 2002).

Figura 1. Ubicación de la Sierra de Zenta en el noroeste argentino y localidades estudiadas. / Figure 1. Location of the Sierra de Zenta in Northwest Argentina and localities studied. 1. Abra de Zenta, 2. Abra Límite, 3. Abra Blanca, 4. Abra Rocío, 5. Santa Ana, 6. Caspalá, 7. Abra Azul, 8. Abra Buey Ojo, 9. Doblozno, 10. Laguna Verde, 11. Abra Llana.
El posterior conocimiento de las microforas y su evolución, el análisis sedimentológico de los perfiles relevados y la información aportada por otros fósiles allí presentes (e.g. invertebrados, trazas) (Aceñolaza et al., 2001, 2003; Aceñolaza y Milana, 2005; Aceñolaza y Aceñolaza, 2007; Aceñolaza y Heredia, 2008; Tortello y Aceñolaza, 2010; Noetinger et al., 2016), contribuyó en estas primeras etapas a una mejor comprensión de los aspectos paleoambientales, que fueron relevantes para un entendimiento general de la geología en esta región del NOA durante tiempos paleozoicos.
El presente trabajo es una revisión actualizada con una reevaluación de la información palinológica existente para el Paleozoico que afora en diversas secciones de la Sierra de Zenta, y que permitieron ajustar las edades que se extienden desde el Tremadociano hasta el Penn-sylvaniano.
Entre los materiales analizados se identifican tanto formas de origen marino (acritarcos, quitinozoos, algas prasinofíceas) como continental (criptosporas, granos de polen y esporas), cuya importancia bioestratigráfica y su relación con otros restos fósiles hallados (trilobites, graptolites, conodontes, braquiópodos, equinodermos, moluscos y trazas fósiles) contribuyen a clarificar el escenario paleoambiental para la región.
La información está organizada por edades, desde la más antigua a la más moderna, y por la ubicación geográfica de las localidades portadoras (Fig. 1), ilustrándose los palinomorfos más destacados de cada período, tomados de los trabajos originales citados. Al final del texto se incluye un apéndice con el listado de géneros y especies mencionados.
Ordovícico
El Ordovícico afora en diferentes sectores de la Sierra de Zenta, pero las localidades estudiadas están ubicadas principalmente en el sector norte y centro de la misma. En la sección norte se identificó una secuencia con base discordante que inicia en la localidad de Abra de Zenta y se extiende hacia el sur hasta Abra Límite, donde los estratos ordovícicos están separados por una discordancia de los niveles suprayacentes asignados al Silúrico, Devónico y Carbonífero (Aceñolaza et al., 2001, 2003; Aráoz y Vergel 2001; Aráoz, 2002, 2009 a, b; Vergel et al., 2008 a, b).
En el sector central las secuencias ordovícicas aforantes muestran espesores cercanos a los 1000 metros y fueron descriptas en las secciones de Santa Ana - Caspalá (Tremadociano, filoiano), Abra Buey Ojo - Doblozno (Tremadociano, filoiano, Dapingiano) y Laguna Verde (Darriwiliano/Sanbdiano) (Aráoz y Aceñolaza, 2004; Aráoz et al., 2008, 2015; Aráoz, 2009 a, b; di Pasquo et al., 2011). De la Puente y Rubinstein (2013) identificaron quitinozoos en la Sierra de Zenta: Euconochitina paschaensis en niveles del Tremadociano tardío asignados a la Formación Santa Rosita, cercanos a la localidad de Santa Ana; Lagenochitina sp. del filoiano medio temprano de la parte superior de la Formación Acoite y Eremochitina baculata del filoiano medio tardío de la parte superior de la Formación Acoite, y Tanuchitina sp. cf. fistulosa en la Formación Caspalá en cercanías de la localidad de Caspalá.
Los datos palinológicos de niveles ordovícicos fueron integrados y permitieron definir cinco asociaciones en base a la distribución de los elementos identificados, tres de las cuales fueron definidas en el perfil de Abra de Zenta y dos en un perfil integrado a partir de secciones aforantes en el sector central de la sierra (Aráoz, 2009 a). Algunos palinomorfos destacados del Ordovícico son ilustrados en la figura 2.
Abra de Zenta
Los estudios palinológicos realizados en esta localidad proceden de un perfil que se extiende entre Abra de Zenta y Abra Límite (Aceñolaza et al., 2001, 2003; Aráoz, 2002, 2009 a, b) el cual aportó la primera información conocida de microfósiles asociados a invertebrados y trazas fósiles. Las trazas procedende los niveles inferiores y medio del perfil y fueron referidas a una típica icno asociación de Cruziana, de la transición cambro - ordovícica, mientras que los invertebrados incluyen trilobites, braquiópodos, gasterópodos y rostroconchos, siendo el conjunto, los primeros restos fósiles del Ordovícico de la Sierra de Zenta.
Los palinomorfos presentes en el perfil son relativamente abundantes siendo el principal componente los acritarcos, acompañados en menores y variables proporciones por prasi-nofitas, quitinozoos y criptosporas. A partir de sus distribuciones estratigráficas Aráoz (2009 a), identificó tres asociaciones palinológicas (Asociación Sierra de Zenta): ASZ1, ASZ2 y ASZ3. En el sector inferior a medio del perfil asignado a la Formación Santa Rosita (Grupo Santa Victoria), se definió la ASZ1 en base a la presencia exclusiva de los acritarcos: Acanthodiacrodium angustum (Fig. 2.1), Acanthodiacrodium spinum, Acanthodiacrodium ubui, Balthisphaeridium sp. 1, Crista-llinium cambriense, Cristallinium ovillense, Cymatiogalea cuvillieri, Cymatiogalea velifera, Saharidia downieii, Saharidia fragilis (Fig. 2.3), Vulcanisphaera africana (Fig. 2.2) y Vulcanisphaera britannica. Acompañan otros acritarcos (Acanthodiacrodium spp., Cymatiogalea membranispina, Cymatiogalea spp., Dasydiacro-dium spp., Eliasum llaniscum, Eupoikilofusa sp.1, Ferromia pelita, Leiofusa spp., Micrhystridium spp., Polygonium spp., Vulcanisphaera tuberata, Vulcanisphaera turbata), prasinofitas (Cymatiosphaera spp., Leiosphaeridia spp., Pterospermella spp.) y criptosporas indeterminadas. La edad determinada para la ASZ1 es Tremadociano temprano tardío, debido a que Acanthodiacrodium angustum está restringido estratigráficamente al Tremadociano y Saharidia fragilis es registrada a nivel mundial en el lapso Cámbrico tardío - Tremadociano (Moczydłowska et al., 2004). Además hay una abundancia de formas típicamente tremadocianas como Acanthodiacrodium, Dasydiacrodium, Polygonium y Vulcanisphaera. Confrman esta edad los trilobites de la Biozona de Kainella meridional inde-finiendo así, los niveles más antiguos identificados en la sierra (Aceñolaza et al., 2001, 2003).
La ASZ2 procede de los sectores medios del perfil, de niveles de la Formación Acoite (Grupo Santa Victoria), definida por los acritarcos: Cymatiogalea granulata, Dactylofusa velifera var. brevis, Eisenackidium orientalis, Eisenackidium sp. 1, Eupoikilofusa spp., Polygonium gracile y Vavrdovella areniga. La asociación incluye además otros acritarcos, prasinofitas (Pterospermella sp.), criptos-poras y los primeros quitinozoos identificados en la sierra (Conochitina sp.). A pesar de que el área donde se define la ASZ2 estaría incluida la transición tremadociana - foiana y de la presencia de Dactylofusa velifera var. brevis y Eisenackidium orientalis que por su amplia distribución en el filoiano de la Cuenca Andina Central son reconocidas como especies guía para el NOA, se destaca la ausencia de formas típicas de la asociación messaoudensis - trifdum, característica de esa edad, que si fue reconocida en otras localidades del Noroeste Argentino. Por ello, la ASZ2 es considerada de edad foiana, mientras que el quitinozoo indicaría una edad no más antigua que Tremadociano tardío (Aráoz, 2009 a).
En el sector superior del perfil, en niveles de la Formación Acoite, fue definida la asociación ASZ3 en base a los acritarcos: Balthisphaeridium coolibahense, Balthisphaeridium omnivagum, Balthisphaeridium sp. cf. B. crinitum, Barakella rara (Fig. 2.4), Barakella felix, Polygonium dentatum, Stelliferidium sp. y Tectitheca sp. cf. T.additionalis, prasinofitas: Pterospermella sp. y criptosporas. Barakella tiene su First Appearance Datum (FAD) en la base del Dapingiano, aunque Rubinstein et al. (2007) lo registran en el filoiano tardío de Cordillera Oriental, por lo que no se descarta esa edad para la ASZ3.
Una discontinuidad en el perfil, cercana a la localidad de Abra Límite, separa los niveles ordovícicos de los suprayacentes, donde la palinología permitió identificar asociaciones que asignan edades Silúrico, Devónico y Carbonífero (Vergel et al., 2008 b; di Pasquo et al., 2011).
Abra Buey Ojo, Doblozno, Abra Llana, Laguna Verde, Santa Ana, Caspalá
Estas localidades del sector central de la sierra, fueron relevadas y estudiadas en conjunto y la información palinológica integradas para un análisis general. Aráoz (2009 a) resumió la información geológica y palinológica unificando los perfiles parciales y definió las asociaciones ASZ4 y ASZ5, establecidas conjuntamente con las ASZ1, ASZ2 y ASZ3 de Abra de Zenta, extendiendo el conocimiento de los sucesos paleoforísticos del Ordovícico de la Sierra de Zenta. El perfil unificado tiene su base en el área de Abra Buey Ojo, donde el sector inferior de esta secuencia heterolítica grano creciente fue parcialmente comparado a la Formación Zanjón (Aráoz y Aceñolaza, 2004; Vergel et al., 2008 b; Aráoz, 2009 a, b). En esos aforamientos fue de-finida la ASZ4, con la presencia exclusiva de los acritarcos: Balthisphaeridium fragile, Coryphidium bohemicum, Cristallinium dentatum, Cymatiogalea deunffi (Fig. 2.5), Dasydiacrodium sp. 1, Eupoikilofusa striatogranulata, Multiplicisphaeridium spp., Picostella turgida, Pirea sp. cf. P. colliformis, Polygonium spp., Stellechinatum uncinatum (Fig. 2.7), Stellechinatum sp. 1, Stelliferidium stelligerum, Striatotheca princi-palis (Fig. 2.12), Tectitheca additionalis, Tectitheca sp. cf. T. additionalis, Veryhachium lairdii group y Veryhachium symmetricum. Acompañan otros acritarcos como Acanthodiacrodium spp., Cymatiogalea granulata, Cymatiogalea sp., Dactylofusa velifera var. brevis (Fig. 2.8), Eisenackidium orientalis (Fig. 2.9), Eisenackidium sp. 1, Leiofusa sp., Micrhystridium spp., Polygonium gracile, Stelliferidium sp., Vavrdovella areniga (Fig. 2.10) y Vulcanisphaera tuberata; las prasinofitas Cymatiosphaera spp., Leiosphaeridia spp. y Pterospermella sp.; los quitinozoos Euconochitina sp. y Eremochitina sp. (Fig. 2.14) y criptosporas indeterminadas (Fig. 2.6). Para la ASZ4 se asignó una edad filoiano, cercana al límite Trema-dociano / filoiano, parcialmente equivalente a la ASZ2 definida en Abra de Zenta, aunque la composición de las microforas es marcadamente diferente, con una abundancia y diversidad mayor en la ASZ4 que en la ASZ2 (Aráoz, 2009 a).
Aforamientos ordovícicos continúan de manera saltuaria hacia el sur, y a un kilómetro de distancia aproximadamente, se reconocieron niveles con microforas vinculadas a graptolites de los géneros Baltograptus, Didymograptus, Expansograptus y Tetragraptus y restos de trilobite Thysa-nopyge, registros que mencionan Aráoz (2009 a, b) y Aráoz et al. (2015). La secuencia continúa hasta Laguna Verde, donde se encuentran los niveles ordovícicos más jóvenes reconocidos hasta el momento, representados por la Formación Caspalá. Allí la secuencia es predominantemente arenosa, con alta tasa de bioturbación y presencia de trazas fósiles (Cruziana, Lockeia, Monomor-phichnus y Skolithos) asociadas a una fauna de conodontes de importancia bioestratigráfica (Ace-ñolaza y Heredia, 2008; Carlorosi y Heredia, 2013; Aceñolaza et al., 2015). La ASZ5 fue definida a partir de los niveles con graptolites y continúa hasta Laguna Verde mostrando una amplia extensión en cuanto a niveles involucrados y a diversidad taxonómica, con niveles que tienen una gran abundancia y diversidad palinológica, a pesar de lo cual no se pudo establecer una subdivisión. En general, los taxones presentes en la ASZ5 tienen rangos de distribución amplios a partir del filoiano o Dapingiano y se extienden hasta el Darriwiliano e incluso al Hirnantiano. En esa heterogeneidad de formas, se destaca una amplia representación de acritarcos considerados clave como Dactylofusa velifera var. brevis, Eisenackidium orientalis, Vavrdovella areniga, Stellechinatum uncinatum y Coryphidium que fueron decisivos en la definición de una sola asociación. La ASZ5 se caracteriza por los acritarcos Acanthodiacrodium crassus, Adorfa hoffmanensis, cf. Arkonia sp., Au-reotesta sp., Coryphidium sp. cf. C. elegans, Coryphidium miladae - tadla group (Fig. 2.13), Coryphidium spp., Cymatiogalea messaoudensis var. messaoudensis (Fig. 2.11), Ericanthea pollicipes, Leiofusa fusiformis, Leiofusa tumida, Petaloferidium bulliferum, cf. Picostella sp. 1, Pirea sp. cf. P. ornata, Poikilofusa spp.,

Figura 2. Palinomorfos del Ordovícico. 1-4: Abra de Zenta. 5, 6, 7, 12: Abra Buey Ojo. 11, 13: Laguna Verde. 1-13: escala 10 µm; 14: escala 100 µm / Figure 2. Ordovician palynomorphs. 1-4: Abra de Zenta. 5, 6, 7, 12: Abra Buey Ojo. 11, 13: Laguna Verde. 1-13: ííuá 10 iim, 14: scak 100 iim. 1. Acanthodiacrodium angustum, 2. Vulcanisphaera africana, 3. Saharidia fragilis, 4. Barakella rara, 5. Cymatiogaka deunffi, 6. criptosporas indeterminadas / indeterminated cryptospores, 7. Stelkchinatum uncinatum, 8. Dactyhfusa velifera var. brevis, 9. Eisenacki " ' talis, 10. Vavrdovella areniga, 11. Cymatiogaka messaoudensis var. messaoudensis, 12. Striatotheca prinápalis, 13. Coryphidium milada - tadla Group, 14. Uremochitina sp.
Priscogalea sp., Stelliferidium striatulum, Striatotheca rugosa, Striatotheca sp. cf. S. transformata, Striatothe-ca spp., Tectitheca valida, Veryhachium minutum, Veryhachium rhomboidium, Veryhachium trispinosum group. Esta asociación presenta una mayor diversidad de quitinozoos (Euconochitina sp., Eremo-chitina sp. y Lagenochitina sp.) y prasinofíceas como Pterospermella colbathii. La edad asignada para la ASZ5, prácticamente en toda su extensión, es Dapingiano / Darriwiliano, más probablemente Darriwiliano (Aráoz, 2009 a). Los niveles más altos cercanos a Laguna Verde incluyen una asociación de conodontes recientemente revisada que contiene Erraticodon patu, Baltoniodus trian-gularis, Baltoniodus cf. B. triangularis, Trapezognathus diprion, entre otros, identificándose la Zona B. triangularis que sugiere una edad Dapingiana temprana (Heredia et al., 2008; Carlorosi y Heredia, 2013; Aceñolaza et al., 2015), lo cual permite ajustar mejor la edad de la palinofora.
De la Puente y Rubinstein (2013) mencionaron para niveles tremadocianos asignados a la Formación Santa Rosita cercanos a la localidad de Santa Ana, quitinozoos equivalentes a la asociación de Euconochitina paschaensis; también quitinozoos asignados al filoiano de la Formación Acoite y otros para el Ordovícico más tardío de la Formación Caspalá, con Tanuchitina sp. cf. fistulosa, de pobre preservación que probablemente indique un retrabajo por la glaciación hirnantiana.
Silúrico
Abra Límite
Los únicos niveles silúricos estudiados palinológicamente en la Sierra de Zenta se encuentran en la localidad de Abra Límite, en un intervalo entre discordancias que separan estratos del Silúrico de los infrayacentes del Ordovícico y suprayacentes del Devónico temprano. Allí, di Pasquo et al. (2010) identificaron dos niveles con palinomorfos de regular preservación, fuerte piritización y alto grado de madurez térmica (TAI alrededor de 3), con esporas triletes, acritarcos, quitinozoos y criptosporas. di Pasquo et al. (2010) reconocen dos asociaciones pali-nológicas, la asociación inferior (A1) que incluye: Ambitisporites avitus (Fig. 3.1), Amicosporites sp. (Fig. 3.10), Quadrisporites variabilis (Fig. 3.2), Crassiangulina variacornuta (Fig. 3.3), Duvernaysphaera sp. cf. D. blanca (Fig. 3.12), Eisenackitina sp. cf. E. bohemica, Margachitina sp. cf. M. saretensis, An-cyrochitina fragilis (Fig. 3.6), Angochitina sinica (Fig. 3.4) y Angochitina sp. cf. A. filosa. Según estos autores, el conjunto sugiere una edad no más antigua que Aeroniano y no más joven que Prido-liano - Lochkoviano. La asociación superior (A2) incluye entre sus taxones: Crassiangulina varia-cornuta, Quadrisporites variabilis, Quadrisporites granulatus, Duvernaysphaera sp. cf. D. blanca, Ononda-gaella asymmetrica (Fig. 3.5), Veryhachium valiente, Veryhachium downieii, Leiofusa banderillae (Fig. 3.7), Goniosphaeridium sp. cf. G. uncinatum, Diexalophasis denticulata (Fig. 3.8), Cheilotetras sp. (Fig. 3.11), Imperfectotriletes sp., Dyadospora murusattenuata, Synsphaeridium sp., Eisenackitina sp. cf. E. bohemica, Desmochitina sphaerica, Angochitina chlupaci (Fig. 3.9), Angochitina sinica y Conochitina pachycephala. La asociación de acritarcos y quitinozoos mencionados, sugieren una edad Ludlowiano - Pridolia-no, por lo que ambas asociaciones incluyen formas que atraviesan el límite Silúrico - Devónico, siendo evidencia de la presencia de este lapso de tiempo en la Sierra de Zenta.

Figura 3. Palmomorfos del Silúrico. Escala 10 j_im / Figure 3. Silurian palynomorphs. Scale 10 ¡im. 1. Ambitisporites avitus, 2. Quadrisporites variabilis, 3. Crassiangx/Iina variacornuta, 4. Angochitina sínica, 5. Onondagaella asymmetrica, 6. Ancyrochitina fragi/is, 7. Leiofasa banderillae, 8. Diexalophasis denticulata, 9. Angochitina chlupaá, 10. Amicosporites sp., 11. Chálotetras sp., 12. Duvernaysphaera sp. cf. D. blanca.
Devónico
Los niveles de esta edad que fueron reconocidos en la Sierra de Zenta se encuentran en dos aforamientos, uno de ellos en la localidad de Abra Límite y el otro en Abra Azul. Los palinomorfos identificados son ilustrados en la figura 4.
Abra Límite
En los niveles devónicos de esta localidad se recuperaron palinomorfos entre los que se identificaron criptosporas, esporas triletes, acritarcos, prasinofitas, clorofitas y quitinozoos (Noetinger y di Pasquo, 2008; Vergel et al., 2008 b; di Pasquo et al., 2011). A partir de su análisis, Noetinger y di Pasquo (2010) diferenciaron dos asociaciones en estos niveles, la AL1 proveniente de los sectores basales y medios del aforamiento, que incluyen principalmente elementos marinos como los quitinozoos Angochitina filosa (Fig. 4.3), Cingulochitina serrata, Angochitina chlupaci, Hoegisphaera sp. cf. H. glabra (Fig. 4.5), Ancyrochitina sp. cf. A. tomentosa y Ramochitina sp. y los acritarcos: Multiplicisphaeridium robertinum, Diexallophasis remota, Multiplisphaeridium ramuscu-losum (Fig. 4.6), Dictyotidium variatum (Fig. 4.4) y Veryhachium sp. La asociación AL2, proviene del sector superior del perfil y se caracteriza por una disminución de la biodiversidad y un predominio de elementos continentales como esporas (Retusotriletes sp., Fig. 4.8) y microfitoplancton de agua dulce y salobre (Quadrisporites sp., Fig. 4.7) incluyendo restos de fitoclastos, mientras que los acritarcos (Ammonidium garrasinoi, Fig. 4.1) y quitinozoos son raros o están ausentes.
Los componentes de la AL1 permiten definir para la misma una edad Lochkoviano s.l., considerando que no hay especies restringidas al Silúrico tardío, mientras que en la AL2, la aparición de Dibolisporites farraginis (Fig. 4.2) sugiere una edad Eifeliano tardío a Givetiano (Devónico Medio). La presencia de la diamictita suprayacente a AL2 datada como Pennsylvaniano (di Pasquo y Vergel, 2008) refuerza la edad asignada. Vergel et al. (2008 a) indican una edad Devónico tardío a Mississippiano tardío al hiato de esta sección.
Abra Azul
Noetinger (2011) y Noetinger et al. (2015) analizaron los datos procedentes de este sector del Devónico que incluye palinomorfos, braquiópodos (Australospirifer antarticus, Probos-cidina arcei, Australocoelia tourtelotti, Notiochonetes falklandica) y trazas fósiles que fueron asignados a niveles de la Formación Pescado.
La palinofora incluye esporas trilete (Acinosporites lindlarensis var. minor, Ambitisporites avitus/dilutus morphon, Apiculiretusispora brandtii (Fig. 4.10), Apiculiretusispora plicata, Archaeozono-triletes chulus var. chulus, Cymbosporites senex, Dibolisporites bullatus, Dibolisporites nodosus (Fig. 4.11), Dibolisporites quebencensis (Fig. 4.12), Dictyiotriletes emsiensis morphon (Fig. 4.13), Leiotriletes sp., Retusotriletes albarinii, Retusotriletes maculatus, cf. Streelispora granulata, Synorisporites papillensis); crip-tosporas (Cymabohilates sp. cf. C. comptulus, Dyadospora murusattenuata, Gneudnaspora divellomedia var. minor, Tetrahedraletes medinensis); acritarcos (Veryhachium lairdii group, Veryhachium trispinosum group); clorofitas (Quadrisporites granulatus, Quadrisporites variabilis) y quitinozoos (Angochitina sp. A, en Noetinger y di Pasquo, 2013, Fig. 4.9). Según los trabajos de Noetinger (2011) y Noetin-ger et al. (2016) la distribución estratigráfica de estos palinomorfos sugiere una edad Lochko-viano tardío a Pragiano - Emsiano temprano, aunque la co-ocurrencia con los braquiópodos identificados, limita la edad de los niveles a un Pragiano tardío y Emsiano temprano.

Figura 4. Palinomorfos del Devónico. 1-8: Abra Límite. 9-13: Abra Azul. Escala: 20 µm./ Figure 4. Devonian palyno-morphs. 1-8: from Abra Límite. 9-13: from Abra Azul. Scale: 20 µm. 1. Ammonidium garrasinoi, 2. Dibolisporites farraginis, 3. Angochitina filosa, 4. Dictyotidium variatum, 5. Hoegisphaera sp. cf. H. glabra, 6. Multiplicisphaeridium ramusculosum, 7. Quadrispo-rites granulatus, 8. Retusotriletes sp., 9. Angochitina sp., 10. Apiculiretusispora brandtii, 11. Dibolisporites? nodosus, 12. Dibolisporites quebecensis, 13. Dictyotriletes emsiensis morphon.
Carbonífero
El Carbonífero fue identificado en aforamientos ubicados en el norte de la Sierra de Zenta, en la sección ubicada entre Abra Límite y Abra Rocío y además en la localidad de Abra Blanca que cuenta con muy escasos datos palinológicos (Vergel et al., 2008 a, b). Los palinomor-fos carboníferos son ilustrados en la figura 5.
Abra Límite - Abra Rocío
Los niveles del Pennsylvaniano que fueron identificados y estudiados por di Pasquo y Vergel (2008) y di Pasquo et al. (2011), se encuentran ubicados en una sección extendida entre Abra Límite y Abra Rocío, en aforamientos que corresponden a la Formación Tarija, en el sector más austral de la Cuenca Tarija. Allí se identificaron esporas: Cristatisporites chacoparanensis (Fig. 5.1), Cristatisporites scabiosus, Cristatisporites stellatus (Fig. 5.8), Cristatisporites menendezii, Stenozonotriletes menendezii, Vallatis-porites arcuatus (Fig. 5.9), Granulatisporites varigranifer (Fig. 5.6), Endosporites zonalis, Leiotriletes tenuis (Fig. 5.3), Retusotriletes anfractus (Fig. 5.2), Raistrickia accinta, Velamisporites australiensis (Fig. 5.11), Apiculispori-tes variornatus (Fig. 5.10) junto a varias especies de los géneros Punctatisporites, Calamospora y Leiotriletes. Además se identificaron granos de polen monosacado: Potonieisporites novicus, Potonieisporites lelei (Fig. 5.7), Potonieisporites magnus (Fig. 5.4), Divarisaccus stringoplicatus (Fig. 5.5) y Plicatipollenites malabarensis. Las características de la asociación polínica y distribución estratigráfica de los taxones indicaría que se corresponden con la Biozona de Dictyotriletes bireticulatus - Cristatisporites chacoparanensis (BC) de la Cuenca Tarija (di Pasquo, 2003), asignada al Carbonífero Tardío medio (Pennsylvaniano, Moscovia-no). Esta asignación estaría sustentada por el registro de formas exclusivas de la biozona (Raistrickia accinta, Endosporites zonalis, Velamisporites australiensis y Cristatisporites chacoparanaensis), además de la presencia de especies comunes que se reconocen en las diferentes secciones como Cristatisporites stellatus, Velamisporites australiensis y Granulatisporites varigranifer.
Abra Blanca
En niveles aforantes en cercanía de la localidad de Abra Blanca, Vergel et al. (2008 a,b) estudiaron una microfora cuya principal composición es de granos de polen monosacados y esporas triletes que permitieron asignar una edad pennsylvaniana. Estas microforas contienen especies que fueron comparadas con las de otras localidades de la Cuenca Tarija, tanto del norte de Argentina y del sur de Bolivia, como de otras cuencas neopaleozoicas de Argentina y Brasil. Estas autoras confrman la existencia de la discordancia Devónico - Carbonífero en la región, a la vez que establecen que el hiato abarca por lo menos el lapso de tiempo desde el Devónico tardío hasta el Mississippiano tardío (di Pasquo et al., 2011).
Evolución paleoambiental de la Sierra de Zenta
Los datos de palinomorfos revisados fueron utilizados para realizar un análisis de la evolución paleoambiental para la Sierra de Zenta desde el Ordovícico hasta el Carbonífero (Fig. 6).
Durante el Ordovícico la región de la Cuenca Andina Central se ubicaba en el margen gondwánico occidental y en paleolatitudes intermedias, a partir de lo cual se reconocen extensos depósitos de plataforma con importantes variaciones litológicas laterales como resultado de los diversos paleoambientes y restricciones a nivel local (Astini, 2003). Para la región de Zenta la sedimentación habría sido sobre el fanco oriental de la prolongación norte del Cratógeno Central Argentino, donde se ha inferido una restricción ambiental que caracterizaba la vida en la plataforma con aguas someras (Aceñolaza et al., 2001).
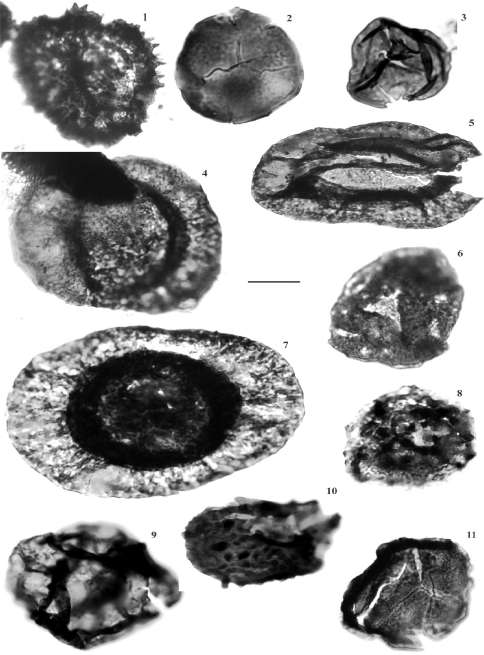
Figura 5. Palinomorfos del Carbonífero. Escala: 30 |im / Figure 5. Carboniferous palynomorphs. Scale: 30 fim. 1. Crista-tisporites chacoparanensis, 2. Retusotri/etes anfractus, 3. Leiotriktes tennis, 4. Potoniesporites magnas, 5. Divarisaccus stringoplicatus, 6. Granulatisporites varigranifer, 7. Potoniesporites lelei, 8. Cristatisporites stellatus, 9. Vallatisporites arcuatus, 10. Apiculatisporites variornatus, 11. Velamisporites australiensis.

Figura 6. Cuadro con la litoestratigrafía y localidades con las asociaciones mencionadas en el texto. / Figure 6. Table with the lithostratigraphy and localities with the associations mentioned in the text. A. Abra de Zenta, B. Abra Buey - Laguna Verde, C. Abra Límite, D. Abra Azul, E. Abra Límite - Abra Rocío, F. Abra Blanca.
En general, la mayoría de los modelos de distribución para el Paleozoico temprano asùmen que al igual que el fitoplancton moderno, los acritarcos muestran una correlación directa entre su diversidad y morfología con los paleoambientes en que vivían, aunque en algunos casos consideran que a lo largo de una transecta onshore - offishore, la más alta diversidad de acritarcos se encuentra en ambientes de plataforma externa, mientras que las asociaciones de microfitoplancton de plataforma interna y ambientes de cuenca profunda se caracterizan por una diversidad reducida y dominio de formas simples o esferomorfos. No obstante, hay especies que muestran limitaciones en paleoambientes específicos que no pueden dejar de ser consideradas (Vecoli, 2000, 2004; Tongiorgi et al., 2003).
Las características paleogeográficas y paleoambientales repercutieron en las biotas presentes que muestran diferencias según la ubicación geográfica de las localidades estudiadas y que, en el caso de las microforas son muy marcadas, por lo cual las correlaciones de las mismas en la Sierra de Zenta, no siempre fueron posibles.
El Abra de Zenta es una sección que fue interpretada como un ambiente restringido de plataforma mostrando una somerización hacia el sector medio, donde se encuentran niveles de coquinas producto de eventos tempestíticos. Incluyen invertebrados e "icnofacies de Cruziana", cercanas a niveles arenosos con óndulas indicando zonas con energía baja a media, y donde se observa una posterior profundización (Aceñolaza et al., 2003). Las microforas en esta sección son en general de baja diversidad, con un dominio de formas simples (Saharidia), pequeñas (Micrhystridium), o sensibles a las condiciones paleoambientales (Acanthodiacrodium an-gustum), mientras que las criptosporas sugieren en conjunto, paleoambientes proximales, relativamente someros, cercanos al continente o zonas costeras. Estos datos serían concordantes con los obtenidos de la sedimentología y los macrofósiles presentes.
La sección que afora en Laguna Verde ha sido interpretado como depositado en un ambiente de plataforma externa con una somerización hacia la parte superior del mismo, evidenciada por el aumento de los niveles de granulometrías más gruesas. Además, la presencia de trazas de tipo penetrativas (Skolithos) en las areniscas más gruesas y coquinas con restos de braquiópodos, especialmente en la parte superior del perfil, sugiere también una somerización y paleoambientes de mayor energía. Los palinomorfos muestran una disminución en cantidad y diversidad y las proporciones de criptosporas y quitinozoos son menores con respecto a los acritarcos.
En estos niveles, la presencia del conodonte Erraticodon presenta afinidades con la Provincia Australiana y apoya la hipótesis de que durante el Ordovícico el noroeste de Argentina estaba localizado en latitudes medias a altas e incluía faunas de aguas frías (Carlorosi y Heredia, 2013). Asimismo otras formas registradas en Laguna Verde: Baltoniodus triangularis, Baltoniodus cf. B. triangularis y Trapezognathus diprion indicarían la presencia de corrientes con faunas de conodon-tes provenientes de Báltica, también de aguas frías (Bergström, 1990; Bagnoli y Stouge, 1991).
Para los niveles silúricos de Abra Límite, las asociaciones palinológicas y los datos se-dimentológicos de las sección, determinan un paleoambiente de depositación marino de plataforma bajo condiciones euxínicas, que se inferen por la presencia de pirita framboidal en la exina de los palinomorfos. Un aporte continental estaría indicado por las criptosporas y esporas triletes halladas. La existencia de palinomorfos ordovícicos retrabajados en los niveles silúricos y del conglomerado con nódulos ferruginosos entre los paquetes pelíticos - arenosos, sería el refejo de variaciones en el nivel del mar relacionadas a los eventos de transgresión y regresión ocurridos durante el Llandoveriano y Ludlowiano (di Pasquo et al., 2010, 2011).
La evolución paleoambiental del Devónico en la porción sur de la Cuenca de Tarija, en los aforamientos de Sierra de Zenta muestra algunas variaciones analizadas por Noetinger (2011). Para el Abra Límite, Noetinger y di Pasquo (2010) describen los niveles lochkovianos representados por una asociación compuesta principalmente por elementos marinos con una alta proporción de material orgánico amorfo, lo cual sugiere un depocentro marino de plataforma para la Formación Baritú en este sector. Mientras que para el Abra Azul, la información pali-nológica, palinofacial conjuntamente con los datos de braquiópodos y trazas fósiles presentes, sugieren que durante el Pragiano se desarrolló un ambiente marino con una somerización hacia el tope de la sección, en donde la conexión marina parece estar restringida basado en los acritar-cos que sustentan ambientes marinos euxínicos, acorde a las trazas de pirita que tienen algunos ejemplares. La tendencia regresiva es indicada por la desaparición de palinomorfos marinos y aporte de fitoclastos junto a altas proporciones de esporas y criptosporas que indicarían aporte de zonas continentales. La fase de contracción de cuenca propuesta por Alvarez et al. (2003) durante el Pragiano y Emsiano, es evidenciada por la presencia de cuerpos de arena y el aumento en el suministro de componentes terrígenos (alta concentración de fitoclastos, criptosporas y esporas) hacia el tope de la sección.
Di Pasquo y Vergel (2008) reconocen en la Sierra de Zenta localidades (Abra Límite - Abra Rocío, Abra Blanca) con unidades del Pennsylvaniano que son consideradas dentro del sector más austral de la Cuenca Tarija. El paquete de diamictitas de la Formacion Tarija brindó una asociación palinológica comparable a las microforas de otras localidades de la Cuenca Tari-ja en el norte de Argentina y sur de Bolivia y otras cuencas neopaleozoicas de Argentina y Brasil (Cristatisporites stellatus, Granulatisporifer varigranifer) que se presenta con una pobre preservación y variables grados de carbonización (di Pasquo et al., 2011).
Estos trabajos realizados sobre las microforas del intervalo Ordovícico - Pennsylvania-no de la Sierra de Zenta documentan variaciones en su composición con respecto a asociaciones correlativas identificadas en otras localidades del noroeste de Argentina (Volkheimer et al., 1980; Ottone et al., 1992, 1995; Rubinstein et al., 1999, 2007; di Pasquo, 2003, 2007 a, b, 2009; del Papa y di Pasquo, 2007; di Pasquo y Noetinger, 2008 a, b; Noetinger, 2011; Noetinger y di Pasquo, 2011; De la Puente y Rubinstein, 2013; di Pasquo et al., 2015). Estas diferencias com-posicionales y palinofaciales, permiten corroborar la idea de que hubo importantes cambios paleoambientales a nivel local durante la depositación de las sucesiones en todos los niveles estudiados. La integración de los datos palinológicos, de invertebrados y trazas fósiles obtenidos en conjunto con la evidencia sedimentológica apoyan en general, una depositación en ambientes marinos de plataforma restringidos, con continuas variaciones del nivel del mar e indicios de una mayor oxigenación, aportes de origen terrestre y alternancia con depocentros de mayor profundidad e infuencia de corrientes entre el Ordovícico temprano y el Devónico. En las secciones analizadas pudo evidenciarse un hiato en la sedimentación que abarca el Devónico tardío y el Mississippiano. A partir del Pennsylvaniano se evidencia la sedimentación continental infuenciada por eventos de glaciación - deglaciación entre las formaciones Tarija y Escarpment caracterizadas por sucesiones de fangolitas y diamictitas de matriz fina arcillo-limosa (con clastos estriados y angulosos), heterolitas y areniscas finas a gruesas (Schulz et al., 1999; Azcuy y di Pasquo, 2000; Starck y del Papa, 2006).
Conclusiones
Una marcada diversidad de microfósiles de paredes orgánicas (acritarcos, quitinozoos, algas prasinofíceas, criptosporas, esporas y granos de polen) ocurre en las secuencias paleozoicas de la Sierra de Zenta. El análisis de los diferentes grupos palinológicos, su distribución temporal y la comparación con microforas de otras localidades del resto del mundo, permitió establecer correlaciones parciales con biozonas definidas, sustentadas por especies diagnósticas (Aráoz, 2009 a; Noetinger, 2011). En general, las asociaciones identificadas en la Sierra de Zenta, contienen una baja presencia de formas diagnósticas gondwánicas, por lo que las correlaciones son dificultosas y parciales, pero sugieren un endemismo tanto para los tiempos ordovícicos como devónicos temprano a medio (Aráoz, 2009 a; Noetinger, 2011). Las asociaciones palinológicas del Pennsylvaniano (Biozona BC en di Pasquo, 2003) en cambio, contienen especies comunes a otras foras de la Cuenca Tarija caracterizadas por helechos, lico-fitas, esfenofitas (esporas), coniferales y cordaitales (granos de polen monosaccado) y permiten extender su registro al extremo sudoccidental de la cuenca.
El análisis de toda la información palinológica disponible para los niveles paleozoicos de la Sierra de Zenta, en conjunto con el resto de los grupos fósiles y los estudios sedimento-lógicos, permitió lograr una visión más amplia e integradora de los eventos bioestratigráficos ocurridos, como del marco paleoambiental local y paleobiogeográfico en el cual se desarrollaron.
Agradecimientos
Se agradece a las instituciones INSUGEO, MACN, CICYTTP donde fueron desarrolladas las tareas de laboratorio y gabinete, y al INSUGEO por permitir el uso de vehículos para la realización de las diferentes campañas. El presente trabajo fue realizado en el marco del proyecto de CONICET, PIP N° 1522012010018.
Apéndice de especies mencionadas en orden alfabético según diferentes grupos fosilíferos
Acritarcos
Acanthodiacrodium angustum (Downie 1958) Combaz 1967
Acanthodiacrodium crassus (Loeblich y Tappan 1978) Vecoli 1999
Acanthodiacrodium spinum Rasul 1979
Acanthodiacrodium ubui Martin 1969
Acanthodiacrodium spp.
Adorfa hoffmanensis (Cramer, Allan, Kanes y Díez 1974) Ottone en Ottone, Toro y Waisfeld 1992
Ammonidium garrasinoi Ottone 1996
cf. Arkonia sp.
Aureotesta sp.
Balthisphaeridium coolibahense Playford y Wicander 1988
Balthisphaeridium fragile Tongiorgi, Yin, Di Milia 1995
Baltisphaeridium omnivagum Playford y Wicander 1988
Baltisphaeridium sp. cf. B. crinitum Martin 1978
Balthisphaeridium sp. 1
Barakella felix Cramer y Díez 1977
Barakella rara (Lu 1987) Tongiorgi, Yin y Di Milia 1995
Coryphidium bohemicum Vavrdová 1972
Coryphidium miladae - tadla Group Rubinstein en Rubinstein, Toro y Waisfeld 1999
Coryphidium sp. cf. C. elegans Cramer, Allan, Kanes y Díez 1974
Coryphidium spp.
Crassiangulina variacornuta Wauthoz, Dorning y Le Hérissé 2003
Cristallinium cambriense (Slavikova 1968) Vanguestaine 1978
Cristallinium dentatum (Vavrdová 1976) Martin 1982
Cristalllinium ovillense (Cramer y Diez 1972 b) Martin 1982
Cymatiogalea cuvillieri (Deunff 1961) Deunff 1964
Cymatiogalea deunffi Jardiné, Combaz, Magloire, Peniguel y Vachey 1974
Cymatiogalea granulata Vavrdová 1966
Cymatiogalea membranispina Deunff 1961
Cymatiogalea messaoudensis Jardiné, Combaz, Magloire, Peniguel y Vachey 1974 var. messaoudensis autonym.
Cymatiogalea velifera (Downie 1958) Martin 1969
Cymatiogalea spp.
Dactylofusa velifera (Cocchio 1982) Albani 1989 var. brevis sensu Albani 1989
Dasydiacrodium sp. 1
Dasydiacrodium spp.
Dictyotidium variatum Playford 1977
Diexalophasis denticulata Loeblich 1970
Diexallophasis remota (Deunff 1955) Playford 1977
Eisenackidium orientalis Rubinstein en Rubinstein, Toro y Waisfeld 1999
Eisenackidium sp.1
Eliasum llaniscum Fombella 1977
Ericanthea pollicipes Cramer y Díez 1977
Eupoikilofusa striatogranulata Vavrdová 1989
Eupoikilofusa sp. 1
Eupoikilofusa spp.
Ferromia pellita (Martin 1975) Martin 1996
Goniosphaeridium sp. cf. G. uncinatum (Downie 1958) Kjellstrom 1971
Leiofusa banderillae Cramer 1964
Leiofusa fusiformis (Eisenack 1934) Eisenack 1938
Leiofusa tumida Downie 1959
Leiofusa spp.
Micrhystridium spp.
Multiplisphaeridium ramusculosum Deunff 1976
?Multiplicisphaeridium robertinum (Cramer 1964) Lister 1970
Multiplicisphaeridium spp.
Onondagaella asymmetrica (Deunff 1954) Cramer 1966
Petaloferidium bulliferum Yin, Di Milia y Tongiorgi 1998
Picostella turgida Yin, Di Milia y Tongiorgi 1998
cf. Picostella sp. 1
Pirea sp. cf. P. colliformis (Burmann 1970) Eisenack, Cramer y Díez 1976
Pirea sp. cf. P. ornata (Burmann 1970) Eisenack, Cramer y Díez 1976
Polygonium dentatum (Timofeev 1959 ex Konzalová-Mazancová 1969) Albani 1989
Polygonium gracile Vavrdová 1966
Polygonium spp.
Poikilofusa spp.
Priscogalea sp.
Saharidia downieii Combaz 1967
Saharidia fragilis (Downie 1958) Combaz 1967
Stellechinatum uncinatum (Downie 1958) Molyneux 1987
Stellechinatum sp. 1
Stelliferidium stelligerum (Górka 1967) Deunff, Górka y Rauscher 1974
Stelliferidium striatulum (Vavrdová 1966) Deunff, Górka y Rauscher 1974
Stelliferidium sp.
Striatotheca principalis Burmann 1970
Striatotheca rugosa Tongiorgi, Yin, Di Milia 1995
Striatotheca sp. cf. S. transformata Burmann 1970
Striatotheca spp.
Synsphaeridium sp.
Tectitheca additionalis Burmann 1968
Tectitheca sp. cf. T. additionalis Burmann 1968
Tectitheca valida Burmann 1968
Vavrdovella areniga (Vavrdová 1973) Loeblich y Tappan 1976
Veryhachium downiei Stockmans y Willière 1962
Veryhachium lairdii group Servais, Vecoli, Li, Molyneux, Raevskaya y Rubinstein 2007
Veryhachium minutum Downie 1959
Veryhachium rhomboidium (Downie 1959) Turner 1984
Veryhachium symmetricum (Lu 1987) Tongiorgi, Yin y Di Milia 1995
Veryhachium trispinosum group Servais, Vecoli, Li, Molyneux, Raevskaya y Rubinstein 2007
Veryhachium valiente Cramer 1964
Veryhachium sp.
Vulcanisphaera africana (Deunff 1961) Rasul 1976
Vulcanisphaera britannica Rasul 1976
Vulcanisphaera tuberata (Downie1958) Eisenack, Cramer y Diez Rodríguez 1973
Vulcanisphaera turbata Martin 1982
Clorofitas
Quadrisporites granulatus (Cramer 1966 b) Cramer y Díez 1972 a
Quadrisporites variabilis (Cramer 1966 b) Ottone y Rosello 1996
Prasinofitas
Cymatiosphaera spp.
Leiosphaeridia spp.
Pterospermella colbathii Vavrdová 1990
Pterospermella spp.
Esporas
Acinosporites lindlarensis Riegel 1968 var. minor McGregor y Camfeld 1976
Ambitisporites avitus/dilutus morphon Steemans, Le Hérissé y Bozdogan 1996
Amicosporites sp.
Apiculiretusispora brandtii Streel 1964
Apiculiretusispora plicata (Allen 1965) Streel 1967
Archaeozonotriletes chulus (Cramer 1966) Richardson y Lister var. chulus Richardson y Lister 1969
Apiculisporus variornatus di Pasquo, Azcuy y Souza 2003
Calamospora sp.
Cheilotetras s p.
Cristatisporites chacoparanensis Ottone 1989
Cristatisporites menendezii (Menéndez y Azcuy 1972) Playford 1978
Cristatisporites scabiosus Menéndez 1971
Cristatisporites stellatus (Azcuy 1975) Gutiérrez y Limarino 2001
Cymbosporites senex McGregor y Camfeld 1976
Dibolisporites bullatus (Allen 1965) Riegel 1973
Dibolisporites farraginis McGregor y Camfeld 1976
Dibolisporites nodosus Turnau 1986
Dibolisporites quebencensis McGregor 1973
Dictyotriletes emsiensis morphon Rubinstein, Melo y Steemans 2005
Endosporites zonalis (Loose 1934) Knox 1950
Granulatisporites varigranifer Menéndez y Azcuy 1971
Imperfectotriletes sp.
Leiotriletes tenuis Azcuy 1975
Leiotriletes sp.
Punctatisporites sp.
Raistrickia accinta Playford y Helby 1968
Retusotriletes albarinii Di Pasquo y Noetinger 2008 a
Retusotriletes anfractus Menéndez y Azcuy 1969
Retusotriletes maculatus McGregor y Camfeld 1976
Retusotriletes sp.
cf. Streelispora granulata en McGregor 1984
Stenozonotriletes menendezii Azcuy 1975
Synorisporites papillensis McGregor 1973
Vallatisporites arcuatus (Marques-Toigo 1974) Archangelsky y Gamerro 1979
Velamisporites australiensis (Playford y Helby 1968) di Pasquo, Azcuy y Souza 2003
Criptosporas
Cymabohilates sp. cf. C. comptulus Breuer, Al-Ghazi, Al Ruwaili, Higgs, Steemans y Wellman 2007
Dyadospora murusattenuata morphon Strother y Traverse 1979 sensu Steemans Le Hérissé y Bozdogan 1996
Gneudnaspora divellomedia (Tchibrikova) Balme 1988 var. minor Breuer, Al-Ghazi, Al-Ruwaili, Higgs, Steemans y Wellman 2007
Tetrahedraletes medinensis (Ströther y Traverse 1979) Wellman y Richardson 1993
Granos de polen monosacado
Divarisaccus stringoplicatus Ottone 1991
Potonieisporites lelei Maheshwari 1967
Potonieisporites magnus Lele y Karim 1971
Potonieisporites novicus Bhardwaj 1954
Plicatipollenites malabarensis (Potonié y Sah 1961) Foster 1975
Quitinozoos
Angochitina chlupaci Paris y Laufeld 1981 en Paris, Laufeld y Chlupáè 1981
Angochitina sinica Cramer 1970
Angochitina filosa Eisenack 1955
Angochitina sp. cf. A. filosa Eisenack 1955
Angochitina sp. A en Noetinger y di Pasquo 2013
Ancyrochitina fragilis Eisenack 1955
Ancyrochitina sp. cf. A. tomentosa Taugourdeau y de Jekhowsky 1960
Cingulochitina serrata Taugourdeau y de Jekhowsky 1960
Conochitina pachycephala Eisenack 1964
Conochitina sp.
Desmochitinas phaerica Taugourdeau y Jekhowsky 1960
Duvernaysphaera sp. cf. D. blanca Pöthe de Baldis 1974
Euconochitina paschaensis de la Puente y Rubinstein 2009
Euconochitina sp.
Eremochitina baculata Taugourdeau y de Jekhowsky 1960
Eremochitina sp.
Eisenackitina sp. cf. E. bohemica Eisenack 1934
Hoegisphaera sp. cf. H. glabra Staplin 1961
Lagenochitina sp.
Margachitina sp. cf. M. saretensis Boumendjel 2002
Ramochitina sp.
Tanuchitina sp. cf. fistulosa Taugourdeau y de Jekhowsky 1960
Conodontes
Erraticodon patu Cooper 1981
Baltoniodus triangularis (Lindström 1955)
Baltoniodus cf. B. triangularis (Lindström 1955)
Trapezognathus diprion (Lindström 1955)
Braquiópodos
Australospirifer antarticus Morris y Sharpe 1846
Proboscidina arcei Ulrich 1892
Australocoelia tourtelotti Boucot y Gill 1956
Notiochonetes falklandica Morris y Sharpe 1846
Graptolites
Baltograptus Maletz 1994
Didymograptus McCoy en Sedgwick y McCoy1851
Expansograptus Bouček y Přibyl 1952
Tetragraptus Salter 1863
Trilobite
Thysanopyge Kayser 1898
Trazas
Cruziana d'Orbigny 1842 Lockeia James 1879 Monomorphichnus Crimes 1970 Skolithos Haldemann 1840
Referencias
Aceñolaza, G.F. y Aceñolaza, F.G. 2007. Bivalvian trace fossils of the Labrado Formation (Lower Ordovician) from Zenta Range (Jujuy province): preservational controls of some push-and-pull deposit-feeder traces. V Reunión Argentina de Icnología y Tercera Reunión de Icnología del Mercosur. Laboratorio de Geología Andina, CADIC / CONICET, Ushuaia, 17. [ Links ]
Aceñolaza, G.F. y Heredia, S. 2008. The status of the Cruziana (Trilobite trace fossil) stratigraphy in Western Gondwa-na: The mixing of Lower and Upper Ordovician Elements in the Central Andean Basin of South America. Cuadernos del Museo Geominero, 9: 13 - 17. Madrid, España. [ Links ]
Aceñolaza, G.F. y Milana, J.P. 2005. Remarkable Cruziana beds in the Lower Ordovician of the Cordillera Oriental, NW Argentina. Ameghiniana, 42: 633-637. [ Links ]
Aceñolaza, G.F., Heredia, S. y Carlorosi, J. 2015. Chronostratigraphy and significance of the Rugosa Group (Cruziana, trace fossil) in the Ordovician strata of the South American Central Andean Basin. Comptes Rendus Palevol., 14: 85-93. [ Links ]
Aceñolaza, G.F., Tortello, M.F., Vergel, M.d.M. y Aráoz, L. 2001. Icnofósiles, trilobites y acritarcos de la Formación Santa Rosita aforante en la región del Abra de Zenta (provincias Salta y Jujuy). Cuarta Reunión Argentina de Icnología y Segunda Reunión de Icnología del Mercosur. (Tucumán), Resúmenes: 22. [ Links ]
Aceñolaza, G.F., Aráoz, L., Vergel, M.d.M., Tortello, M.F. y Nieva, S.M. 2003. Paleontology and biostratigraphy of the Lower Ordovician strata cropping out at the Abra de Zenta (Cordillera Oriental, Jujuy and Salta provinces), NW Argentina. En: Albanesi G.L., Beresi, M.S. y Peralta, S.H.(Eds.), Ordovician from the Andes, Serie Correlación Geológica, 17: 23-28. [ Links ]
Álvarez, L.A., Dalenz Farjat, A., Hernández, R.M. y Albariño, L.M. 2003. Integración de facies y biofacies en un análisis secuencial en plataformas clásticas devónicas del sur de Bolivia y noroeste Argentino. Revista de la Asociación Argentina de Sedimentología, 10 (2): 103-121. [ Links ]
Aráoz, L. 2002. [Palinología de los niveles inferiores de la Formación Santa Rosita, en el Abra de Zenta, Provincias de Salta y Jujuy, República Argentina. Facultad de Ciencias Naturales e I.M.L., Tucumán, Argentina, 189 pp. Seminario Inédito]. [ Links ]
Aráoz, L. 2009 a. [Estudios microforísticos del Ordovícico de la Sierra de Zenta, Cordillera Oriental Argentina. Tesis Doctoral Inédita. Facultad de Ciencias Naturales e I.M.L., Tucumán, Argentina, 364 pp. Inédito]. [ Links ]
Aráoz, L. 2009 b. Microforas ordovícicas en Sierra de Zenta, Cordillera Oriental Argentina. Temas de Paleontología I, Serie Correlación Geológica, 25: 37-94. [ Links ]
Aráoz, L. y Aceñolaza, G.F. 2004. Primeros registros palinológicos para el Tremadociano Superior de la Sierra de Zenta, Cordillera Oriental Argentina. XI Reuniao de Paleobotanicos e Palinólogos. (Gramado, Brasil), Boletim de Resumos: 9. [ Links ]
Araóz, L. y Vergel, M.d.M. 2001. Acritarcos del Ordovícico Inferior (Formación Santa Rosita) en el Abra de Zenta, Cordillera Oriental, Argentina. V Jornadas de Comunicaciones. Facultad de Ciencias Naturales e Instituto Miguel Lillo, Serie Monográfica y Didáctica, 41: 55.
Aráoz, L., Vergel, M.M. y Aceñolaza, G. 2015. Palynomorphs associated fauna and correlation of the Ordovician sequence of Zenta Range (Cordillera Oriental of Argentina). XVI Simposio de Paleobotánica y Palinología, (La Plata), Resúmenes: 30.
Aráoz, L., Aceñolaza, G., Vergel, M.d.M., Heredia, S., Tortello, F. y Milana, J.P. 2008. El ordovícico del sector central de la Sierra de Zenta (Sistema Interandino de las Provincias de Jujuy y Salta): cronoestratigrafía y correlación. XVII Congreso Geológico Argentino. (San Salvador de Jujuy, Argentina), Actas: 339.
Astini, R.A. 2003. The Ordovician Proto-Andean basins. En: Benedetto, J.L. (Ed.), Ordovician fossils of Argentina. Secretaría de Ciencias y Tecnología, Universidad Nacional de Córdoba, pp. 1-74.
Astini, R.A. 2008. Sedimentación, facies, discordancias y evolución paleoambiental durante el Cambro - Ordovícico. En: Coira, B., Zappettini, E.O. (Eds.), 17° Congreso Geológico Argentino: Geología y Recursos Naturales de Jujuy, Relatorio, pp. 50-73.
Azcuy, C.L. y di Pasquo, M.M. 2000. Carbonífero y Pérmico de las Sierras Subandinas, Cordillera Oriental y Puna. En: R. Caminos (Ed.), Geología Argentina. Instituto de Geología y Recursos Minerales, Anales 29: 239-260.
Bagnoli, G. y Stouge, S. 1991. Paleogeographic distribution of Arenigian (Lower Ordovician) conodonts. Anais Academia Brasileira de Ciências, 63 (2): 171-183.
Bergström, S.M. 1990. Relations between conodont provincialism and de changing palaeogeography during the Early Paleozoic. En: McKerrow, W.S. y Scotese, C.R. (eds.): Paleozoic Palaeogeography and Biogeographic. Geological So-ciety of America Memoir, 12: 105- 121.
Carlorosi, J. y Heredia, S. 2013. The ordovician conodont Trapezognathus Lindström, 1955 in the Andean Basin, Argentina. N. Jb. Geol. Paläont., Abh, 267: 309-321.
De la Puente, S. y Rubinstein, C.V. 2013. Ordovician chitinozoans and marine phytoplanckton of Central Andean Basin, northwestern Argentina: A biostratigraphic and paleobiogeographic approach. Review of Palaeobotany and Palynology, 198: 1-162.
del Papa, C. y di Pasquo, M.M. 2007. Palaeoenvironmental interpretation and palynology of outcrop and subsurface sections of the Tarija Formation (Upper Carboniferous), Northwestern Argentina. Journal of South American Earth Sciences, 23: 99-119.
di Pasquo, M.M. 2003. Avances sobre palinología, bioestratigrafía y correlación de las asociaciones presentes en los Grupos Macharetí y Mandiyutí, Neopaleozoico de laCuenca Tarija, provincia de Salta, Argentina. Ameghinia-na, 40 (1): 3-32.
di Pasquo, M.M. 2007 a. Asociaciones palinológicas presentes en las Formaciones Los Monos (Devónico) e Itacua (Carbonífero Inferior) en el perfil de Balapuca, sur de Bolivia. Parte 1. Formación Los Monos. Revista Geológica de Chile, 34 (1): 98-137.
di Pasquo, M.M. 2007 b. Update and importance of the Carboniferous and Permian paleontological records of the Tarija Basin. En: E. Díaz-Martínez, I. Rábano (eds.), 4º European Meeting on Paleontology and Stratigraphy of Latin American (Madrid, http://www.igme.es/4empsla), Instituto Geológico y Minero de España, Serie Cuadernos del Museo Geominero (Madrid) N° 8: 107-112.
di Pasquo, M.M. 2009. Primer registro de megaforas y palinología en estratos de la Formación Tarija (Pennsylvaniano), Arroyo Aguas Blancas, Provincia de Salta, Argentina. Descripción de dos especies nuevas. Andean Geology, 36 (1): 95-123.
di Pasquo, M.M. y Noetinger, S. 2008 a. First record of Early Devonian (Lochkovian) fora from the Santa Rosa For-mation - Alarache, Southern Bolivia. Geologica Acta, 6 (2): 1-21.
di Pasquo, M.M. y Noetinger, S. 2008 b. Resultados preliminares del análisis palinológico del Pozo San Antonio X-1 (Salta) entre 2544 y 1293 m de profundidad. XVII Congreso Geológico Argentino (Jujuy), Actas 1: 347-348.
di Pasquo, M. y Vergel, M. 2008. Primer registro palinológico del Pennsylvaniano del Norte de la Sierra de Zenta, provincia de Jujuy, Argentina. 12º Simposio Brasileiro de Paleobotánica y Palinología (filorianópolis, Brasil), Boletín de resúmenes: 51.
di Pasquo, M., Noetinger, S., Isaacson, P., Grader, G., Starck, D., Morel, E. y Anderson Folnagy, H. 2015. Mid - Late Devonian assemblages of herbaceous lycophytes from northern Argentina and Bolivia: age assessment with palynomorphs and invertebrates and paleobiogeographic importance. Journal of South American Earth Sciences, 63: 70-83.
di Pasquo, M., Vergel, M.d.M., Aceñolaza, G.F., Noetinger, S. y Aráoz, L. 2010. Nueva información palinológica de la Formación Lipeón en Abra Límite, Sierra de Zenta, Jujuy. XIII Simpósio Brasileiro de Paleobotânica y palinología (Salvador de Bahía, Brasil), Anais: 134-135.
di Pasquo, M., Vergel, M.d.M., Noetinger, S., Aráoz, L. y Aceñolaza, G.F. 2011. Estudios palinoestratigráficos del Paleozoico en Abra Límite, Sierra de Zenta, Provincia de Jujuy, Argentina. XVIII Congreso Geológico Argentino, Temas Generales: Paleontología Cambios Climáticos (Neuquén), Actas: 1470-1471.
Heredia, S., Carlorosi, J., Aceñolaza, G. y Aceñolaza, F. 2008. El Género Trapezognathus (Conodonta) en el Ordovícico de la Cordillera Oriental: Implicancias Estratigráficas y Bioestratigráficas. XVII Congreso Geológico Argentino (Jujuy, Argentina), Actas, I: 357.
Manca, N., Heredia, S., Hunicken, M. y Rubinstein, C. 1995. Macrofauna, conodontes y acritarcos de la Formación Santa Rosita (Tremadociano), Nazareno, provincia de Salta, Argentina. Boletín de la Academia Nacional de Ciencias, 60 (3-4): 267-275.
Martin, F. 1982. Some aspects of late Cambrian and early Ordovician acritarchs. En: Basset, M.G. and Dean, W.T. (Eds.), The Cambrian - Ordovician boundary: sections, fossil distributions and correlations (Cardiff), National Museum of Wales, Geological Series 3: 29-40.
Moczydłowska M., Popov, L. y Stockfors, M. 2004. Upper Cambrian-Ordovician successions overlying Timanian com-plexes: new evidence of acritarchs and brachiopods from Kolguev Island, Arctic Russia. Geobios, 37: 239-251.
Noetinger, S. 2011. [Estudios de asociaciones micro y megaforísticas del devónico en la porción sur de la Cuenca Tarija, noroeste argentino y sur de Bolivia: edad, correlación y ambiente de sedimentación. Facultad de Ciencias Naturales y Exactas, Universidad de Buenos Aires, 319 pp. Tesis Doctoral Inédita].
Noetinger, S. y di Pasquo, M. 2008. First record of a Devonian palynological assemblage from the Zenta Range, Eastern Cordillera, Northwestern Argentina. XII Simpósio de Paleobotanicos e Palinólogos (filorianópolis, Brasil), Boletin de resumos: 158.
Noetinger, S. y di Pasquo, M. 2010. Palynomorphs from the Abra Límite, Zenta Range, Eastern Cordillera, Northwes-tern Argentina. Revista Brasilera Paleontologia, 13 (1): 13-20.
Noetinger, S. y di Pasquo M.M. 2011. Devonian palynoforas of the San Antonio x-1 borehole from the Tarija Basin, northwestern Argentina. Geologica Acta, 9 (2): 199-216.
Noetinger, S., di Pasquo, M., Isaacson, P., Aceñolaza, G. y Vergel, M.M. 2016. Integrated study of fauna and microfora from the Early Devonian (Pragian - Emsian) of northwestern Argentina. Historical Biology, 28 (7): 913-929.
Ottone, J.C. 1999. Bioestratigrafía - Palinología. En: González Bonorino, G., Omarini, R. y Viramonte, J. (Eds.): Geología del Noroeste Argentino, XIV Congreso Geológico Argentino (Salta), Relatorio, tomo I: 223-224.
Ottone, J., Toro, B. y Waisfeld, B. 1992. Lower ordovician palynomorphs from the Acoite formation, Northwestern Argentina. Palynology, 16: 93-116.
Ottone, E.G., Waisfeld, B.G. y Astini, R.A. 1995. Acritarcas del Ordovícico Temprano de la Quebrada de Chalala, Noroeste de argentina. Ameghiniana, 32 (3): 237-242.
Ramos, V. 1999. Las provincias geológicas del territorio argentino. En: Caminos, R. (Ed.), Geología Argentina. Instituto de Geología y Recursos Minerales (Buenos Aires), Anales, 29(3): 41-96.
Rubinstein, C.V. 1997. Tremadocian acritarchs from northwestern Argentina. Review of Palaeobotany and Palynology, 98: 41-45.
Rubinstein, C.V. y Toro, B.A. 1999: Acritarch and graptolite biostratigraphy in the Lower Arenig of the peri-Gondwana related Eastern Cordillera, Argentina. Acta Universitatis Carolinae - Geológica, 43 (1/2): 255-258.
Rubinstein, C.V., Toro, B.A. y Waisfeld, B.G. 1999: Acritarch biostratigraphy of the upper Tremadoc - Arenig of the Eastern Cordillera, northwestern Argentina: relationships with graptolite and trilobite faunas. Bollettino della Societa Paleontologica Italiana, 38 (2-3): 267-286.
Rubinstein, C.V., de la Puente, G.S., Toro, B.A. y Servais, T. 2007. The presence of the messaoudensis -trifdum acritarch assemblage (Upper Tremadocian- filoian) in the Central Andean Basin, north-western Argentina: calibration with chitinozoans and graptolite zonation. Acta Palaeontologica Sinica, 46 (suppl.): 422-428.
Schulz, A., Santiago, M., Hernández, R., Galli, C., Alvarez, L. y del Papa, C. 1999. Modelo estratigráfico del Carbonífero en el sector sur de la cuenca de Tarija. 4° Congreso de Exploración de Hidrocarburos (Mar del Plata), Actas II: 695-704.
Starck, D. 1996. Facies continentales en el siluro - devónico de la cuenca del Noroeste. Provincia de Salta, Argentina. XII Congreso Geológico de Bolivia (Tarija), Memorias: 231-238.
Starck, D. 1999. Facies continentales en el Siluro-Devónico de la cuenca del Noroeste. Provincia de Salta, Argentina. Boletín de Informaciones Petroleras, 16: 99-107.
Starck, D. y del Papa, C. 2006. The northwestern Argentina Tarija Basin - Stratigraphy, depositional systems and con-trolling factors in a glaciated basin. Journal of South American Earth Sciences, 22: 169-184.
Tongiorgi, M., Yin, L. y Di Milia, A. 2003. Lower Yushanian to lower Zhejiangian palynology of the Yangtze Gorges area (Daping and Huanghuachang sections), Hubei province, south China. Palaeontographica, Abteilung B, 266: 1-16.
Tortello, F. y Aceñolaza, G. 2010. Trilobites tremadocianos de Abra de Zenta (Cordillera Oriental, Provincias de Jujuy y Salta). Revista de la Asociación Geológica Argentina, 66 (1): 156-163.
Vecoli, M. 2000. Palaeoenvironmental interpretation of microphytoplankton diversity trends in the Cambrian - Or-dovician of the northern Sahara platform. Palaeogeography, Palaeoclimatology, Palaeoecology, 160 (3-4): 329-346.
Vecoli, M. 2004. Stratigraphic and palaeoenvironmental distribution of organic-walled microfossils in Cambrian - Or-dovician transitional strata of borehole Bir Ben Tartar-1 (Tt-1; Ghadamis Basin, southern Tunisia). Memoirs of the Association of Australasian Palaeontologists, 29: 13-30.
Vergel, M.d.M., di Pasquo, M., Aráoz, L. y Noetinger, S. 2008 a. Asociaciones palinológicas del Paleozoico en la Sierra de Zenta, provincia de Jujuy, Argentina. XII Simposio de Paleobotanicos e Palinologos (filorianópolis, Brasil), p. 227.
Vergel, M.d.M., di Pasquo, M., Noetinger, S., Aráoz, L. y Aceñolaza, G.F. 2008 b. Estudios palinoestratigráficos del Paleozoico en la Sierra de Zenta, Provincia de Jujuy, Argentina. II Jornadas Geológicas Fundación Miguel Lillo (Tucumán), Acta geológica Lilloana, Suplemento del volumen, 21 (1): 152-154.
Volkheimer, W. , Melendi, D.L. y Aceñolaza, F.G. 1980. Una microfora ordovícica de la Formación Mojotoro, provincia de Salta. Revista de la Asociación Geológica Argentina, 35 (3): 401-416.
Recibido: 30 de Setiembre del 2016 Aceptado: 10 de Diciembre del 2016

