










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.11 no.1 Córdoba Jan./June 2001
TRABAJOS ORIGINALES
La diversidad del zoobentos en ríos de montaña del noroeste de Argentina: comparación entre seis ríos
Hugo R Fernández1,2, Fátima Romero3,*, Marcela Peralta3 & Luis Grosso3
1 Facultad de Ciencias Naturales e Instituto Miguel Lillo, San Miguel de Tucumán, Argentina
2 Flathead Lake Biological Station, Montana, EEUU
3 Fundación Miguel Lillo, San Miguel de Tucumán, Argentina
* Fundación Miguel Lillo; Miguel Lillo 251; 4000, San Miguel de Tucumán, Argentina. Email: fatiromero@yahoo.com.ar
Recibido: 9 octubre 2000;
Aceptado: 14 marzo 2001
RESUMEN. Se analiza la estructura de la comunidad bentónica de seis ríos pertenecientes a tres cuencas subtropicales del noroeste argentino. Todas las estaciones de muestreo se ubicaron en la ecorregión de las Yungas. Se registraron 78 taxones con una gran diferencia en la densidad total de individuos entre los ríos considerados. Por la similitud taxonómica (50%) se agrupan tres estaciones: ríos Medina, Yala, y Candelaria (con más de 35 taxones), mientras que el resto (ríos Pueblo Viejo, Piedras y Conchas) no superan los 27 taxones. La riqueza, según el análisis de rarefacción para una submuestra de 250 individuos, separa netamente los ríos Yala y Conchas del resto. Según el Análisis de Correspondencia las estaciones se ordenan según los gradientes de magnitud del río y la altura. Los grupos formados por similitud taxonómica, rarefacción y análisis de correspondencia serían producidos más por particularidades locales que por una impronta de cuencas.
ABSTRACT. Zoobenthic diversity in mountain rivers of northwestern Argentina: a comparison among six rivers: The structure of the benthic community of six rivers belonging to three subtropical basins from northwestern Argentina is analyzed. All the sampling stations were located in the mountain rain forest (Yungas) ecoregion. Seventy-eight taxa were recorded, although with a great difference in the total individual density among the considered rivers. Based on their taxonomic similarity (50%), three stations grouped together: Medina, Yala and Candelaria rivers (with more than 35 taxa), while the remaining rivers (Pueblo Viejo, Piedras and Conchas) formed a second group (with 27 taxa or fewer). According to rarefaction analysis of subsamples of 250 individuals, species richness clearly separates Yala and Conchas rivers from the others. According to the Correspondence Analysis, stations were ordered following the gradients of magnitude and altitude of the rivers. The groups formed by taxonomic similarity, rarefaction and Correspondence Analysis were due more to local peculiarities rather than to basin characteristics.
INTRODUCCIÓN
Es ampliamente aceptado que en la región Neotropical hay un mayor conocimiento de los grandes ríos que de los pequeños (Covich 1988). En Argentina solo recientemente se han iniciado estudios detallados sobre el bentos de ríos de distintas regiones (Corigliano & Cosarini 1984; Wais & Bonetto 1988; Marchese & Drago 1992; Corigliano & Freytes 1994; Miserendino 1995; Peso 1995).
En la extensa y heterogénea área que corresponde a la subregión de las Sierras en el noroeste argentino (Bianchi & Yañez 1992), se extienden las selvas subtropicales de montaña. Estas selvas, conocidas como Yungas (Figura 1), están constituidas por tres pisos de vegetación con un claro gradiente de riqueza norte–sur, que son cruzados por numerosos ríos que nacen en las serranías del oeste (Morales 1996). En la provincia de Tucumán, algunos de estos ríos presentan una alta diversidad de macroinvertebrados (Fernández & Palacios 1989; Fernández et al. 1995; Romero & Fernández 2001); sin embargo, la estructura de la comunidad bentónica y los factores que la determinan son desconocidos.

Figura 1. Mapa del noroeste argentino con la ubicación de las estaciones de muestreo y la ubicación de la región de las Yungas (área punteada).
Figure 1. Map of northwestern Argentina showing sampling stations and the distribution of Yungas (dotted area).
Los factores físicos ejercen su influencia sobre los macroinvertebrados de los ríos según una jerarquía de escalas espaciales en la que se reconocen diferentes matices (Brussock et al. 1985; Quinn & Hickey 1990; Wasson & Barrere 1999) que están directamente relacionados con el continuum en los ríos (Vannote et al. 1980). Esta teoría supone que la entrada de energía, el transporte de materia orgánica, su almacenamiento y su uso por los grupos funcionales tróficos de macroinvertebrados pueden ser regulados por procesos geomorfológicos fluviales. Otros autores (Palmer et al. 1996) proponen considerar los factores de control que operan a escalas regionales, como la dispersión, y que no son normalmente tenidos en cuenta. Bajo ciertas circunstancias, factores como las crecientes pueden ser definitorios en la determinación de los conjuntos de organismos bentónicos de un sitio.
En nuestro caso, al trabajar en una macroescala, se establecieron como factores fijos e invariables en toda el área a la vegetación de la cuenca y a la geología (Quinn & Hickey 1990). Estos factores definen a las Yungas como una ecorregión (Vides Almonacid et al. 1998). Dentro de esta extensa área considerada, las cuencas se comportan como unidades geomorfológicas en las cuales es de esperar que la estructura de las comunidades bentónicas de cada una de ellas sea más similar entre sí que con el resto. Sobre esta hipótesis, y buscando un mejor conocimiento de la estructura comunitaria de macroinvertebrados bentónicos en esta ecorregión, se muestrearon tramos de ríos de la zona superior de las tres grandes cuencas hidrográficas del noroeste argentino.
ÁREA DE ESTUDIO
Se seleccionaron estaciones de muestreo en seis ríos del noroeste argentino ubicados en las cabeceras de las tres cuencas más importantes de la ecorregión de las Yungas argentinas: Río Salí, Río Juramento y Río Bermejo (Figura 1). Esta área cumple con las condiciones de aparente homogeneidad en las características geográficas asociadas con factores climáticos, fisiográficos, geológicos, edafológicos y vegetacionales de una ecorregión (Barbour et al. 1996). Los ríos estudiados se eligieron sobre la base de sus similitudes (Tabla 1): Río Medinas (RM) y Río Pueblo Viejo (RPV) en Tucumán, Río de las Piedras (RP) y Río de las Conchas (RCh) en Salta, Río Yala (RY) y Río Candelaria (RC) en Jujuy. Las precipitaciones en esta área ocurren principalmente en verano, oscilando entre 785 y 1119 mm.
Tabla 1. Características físicas y biológicas de los ríos estudiados en el noroeste argentino.
Table 1. Physical and biological features of rivers sampled in northwestern Argentina.
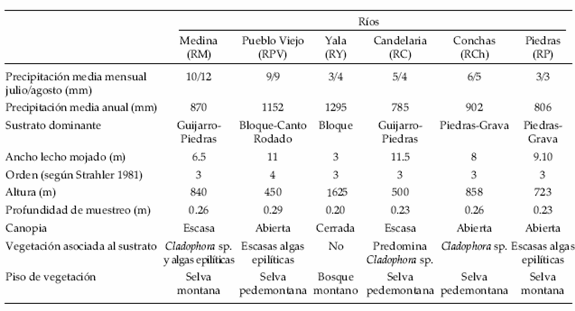
Si bien las estaciones de muestreo están ubicadas dentro de la Provincia Fitogeográfica de las Yungas, varían en cuanto a su ubicación en los diferentes pisos de vegetación. Las ubicadas en RPV, RC y RCh se encuentran en la selva pedemontana (400–600 m), RM y RP en la selva montana (600–1500 m) y RY en el bosque montano (1500–2500 m), según Morales (1996).
MATERIALES Y MÉTODOS
Las muestras fueron tomadas entre el 7 de julio y el 19 de agosto de 1993, durante el período seco estacional que corresponde al invierno. Este período comprende la última fase de colonización del sustrato después del período de lluvias. Se tomaron, en cada caso, dos réplicas con red Surber de parcela cuadrada (900 cm2) que posteriormente, para el análisis, se integraron en una sola. Las muestras se tomaron en el centro de la corriente para disminuir la variabilidad que presenta la zona bentónica y fueron fijadas en el campo con formol al 4%. Los organismos fueron separados bajo lupa estereoscópica binocular y se conservaron en alcohol 75%. La identificación de los diferentes taxones se realizó hasta el nivel más bajo posible. El material fue depositado en la colección del Instituto Fundación Miguel Lillo, bajo la siguiente denominación: colección EB 1992 001 RM (Tuc.) 001, RPV (Tuc.) 001, RP (Sal.) 001, RCh (Sal.) 001, RY (Juj.) 001 y RC (Juj.) 001.
Se calcularon las medidas de riqueza numérica de cada río como una primera descripción de la comunidad (Krebs 1989). Los índices de riqueza y diversidad ofrecen un modo alternativo para describir relaciones de abundancia de especies en las comunidades, permitiendo además ser usados con fines comparativos (Fernández 1997). Ya que no existe un índice que no presente algún defecto en algunos de sus atributos como capacidad discriminatoria y sensibilidad al tamaño de la muestra, elegimos dos que se pueden usar complementariamente: el índice de rarefacción (Magurran 1989) y el índice de Shannon–Wiener.
El índice de riqueza a partir de la rarefacción es muy recomendado (Hurlbert 1971; Krebs 1989) y permite obtener el número de especies esperado en cada muestra si todas tuvieran igual tamaño, minimizando el problema del tamaño muestral (Green 1979). El índice de rarefacción se calculó mediante
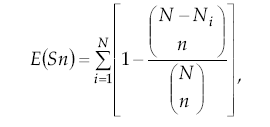
donde: E(Sn) = número de especies esperado, N = número total de individuos, Ni = número total de individuos del taxón i, y n = submuestra de N. La comparación de la riqueza entre estaciones se hizo sobre la base de una submuestra de 250 individuos, determinada por el menor valor entre los totales. Para este cálculo se usó la mínima determinación taxonómica alcanzada, para maximizar las propiedades del índice.
El índice de diversidad de Shannon–Wiener, como una medida de dominancia–uniformidad (Krebs 1989), fue seleccionado por su buena capacidad discriminatoria, su moderada sensibilidad al tamaño muestral y por su facilidad de cálculo (Fernández 1997). Se usó la fórmula

donde: pi = ni/ N y ni = número de individuos del taxón i y N es el número total de individuos. Ante los diferentes niveles alcanzados en las identificaciones, el índice se calculó sobre la base de familias de insectos que, se espera, reflejarán la riqueza de especies ante la dificultad de expresar consistentemente las identificaciones específicas (Bournaud et al. 1996; Jacobsen et al. 1997).
Con fines comparativos, se usó el Índice de Jaccard (Krebs 1989) basado en el grado de similitud entre dos muestras por la presenciaausencia de las especies. Este índice se calcula mediante la fórmula
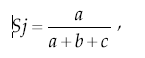
donde a es el número de especies que aparecen en ambas muestras, y b y c corresponden al número de especies en cada una de las dos muestras (Krebs 1989). En este caso también se usaron las familias de insectos. Los resultados se presentan en un diagrama de Trellis (Green 1979) que permite resumir los resultados de un modo gráfico simple.
Para detectar los patrones que pudieran existir en los datos se utilizó un Análisis de Correspondencia. Este análisis define un espacio dentro de un sistema ortogonal de ejes que expresan de un modo conciso un gran número de aspectos diferentes de los datos en un solo diagrama (Krebs 1989; Gauch 1994). Para este fin se usó el programa Canonical Correspondence Analysis 3.1, conocido como CANOCO (ter Braak 1990). Para reducir la influencia de los taxones muy abundantes se transformaron los valores mediante log (x + 1) (Gauch 1994). Dada la característica exploratoria del análisis, se usó la mínima determinación taxonómica alcanzada entre los artrópodos.
RESULTADOS
Durante el trabajo se colectaron un total de 22660 individuos pertenecientes a 78 taxones (Tabla 2). En RM se destaca la dominancia de la especie A de Chironominae (Chironomidae: Diptera), de Tetrahygrobatella argentinensis (Hydrachnidia) y de Hydroptilidae (Trichoptera). Este río también presentó mayor densidad y riqueza (50 taxones), como puede observarse en la Tabla 3. La riqueza esperada para una submuestra de 250 individuos señala a RY como el más rico, seguido por RCh (Figura 2, Tabla 3); en un segundo grupo están RP, RPV, RM y RC, este último con el valor más bajo. El índice de diversidad (Tabla 3) muestra a RY como el más diverso y a RM en último lugar, con un amplio rango entre ambos.
Tabla 2. Composición taxonómica y densidad (individuos / m2) del bentos en los ríos estudiados en el noroeste argentino. Los códigos de los ríos son los mismos que en la Tabla 1.
Table 2. Density (individuals / m2) and taxonomic composition of benthic communities in the rivers sampled in northwestern Argentina. River codes are the same as in Table 1.
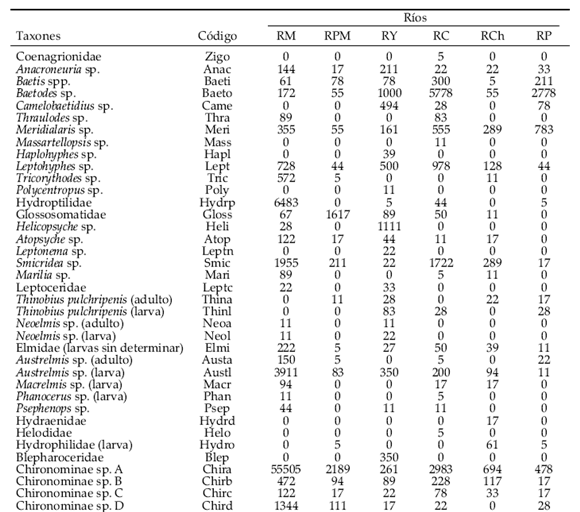

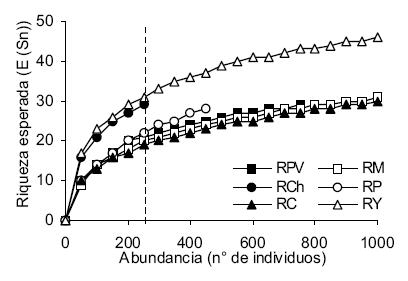
Figura 2. Curvas de rarefacción para los ríos estudiados en el noroeste argentino. Se señala la submuestra de 250 individuos sobre la que se realizó la comparación entre ríos.
Figure 2. Rarefaction curves for the rivers sampled in northwestern Argentina. The subsample of 250 individuals which allows the comparison among rivers is shown.
Tabla 3. Número de individuos y riqueza (de acuerdo a diferentes índices) de los ríos estudiados en el noroeste argentino. Los códigos de los ríos son los mismos que en la Tabla 1.
Table 3. Number of individuals and richness (from different indices) of rivers sampled in northwestern Argentina. River codes are the same as in Table 1.

La similitud de la composición taxonómica entre los ríos separa a RM, RY y RC, con riqueza de familias de insectos mayor a 35 taxones, del resto de las estaciones (Tabla 3, Figura 3). Se destacan los valores extremos con la mayor similitud entre RP y RPV y la menor entre RCh y RY (Figura 3).
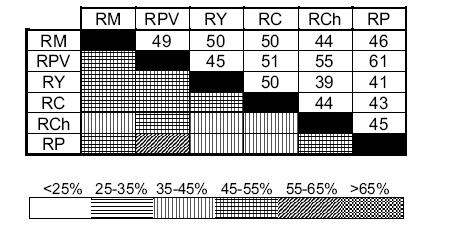
Figura 3. Diagrama de Trellis (Green 1979) mostrando la similitud de la composición taxonómica entre los ríos estudiados en el noroeste argentino. Los códigos de los ríos son los mismos que en la Tabla 1.
Figure 3. Trellis diagram (Green 1979) showing taxonomic composition similarity among the rivers sampled in northwestern Argentina. River codes are the same as in Table 1.
En el Análisis de Correspondencia, el eje I (λ = 0.23) explicó el 32.9% de la variación total, mientras que el eje II (λ = 0.17) explicó el 23.8%. Se observa que entre los ríos muestreados hay dos (RM y RY) que se separan netamente del resto y entre sí (Figura 4a). Estos ríos están caracterizados por un grupo de taxones (Figura 4b). En el extremo superior de la Figura 4a se reúnen el resto de los ríos, notándose que dos de ellos (Ch y RP) pertenecen a la misma cuenca (Río Juramento, Salta).

Figura 4. Análisis de Correspondencia de las muestras de los ríos estudiados en el noroeste argentino. (a) Distribución de las muestras en el plano definido por los componentes I y II. (b) Distribución de los taxones en el plano definido por los componentes I y II. Los códigos de los ríos y de los taxones son los mismos que en las Tablas 1 y 2.
Figure 4. Correspondence Analysis on the samples from the rivers sampled in northwestern Argentina. (a) Distribution of samples in the plane defined by components I and II. (b) Distribution of taxa in the plane defined by components I and II. River and taxon codes are the same as in Tables 1 and 2.
DISCUSIÓN
El río Medina aparece como el de mayor riqueza y abundancia, lo que estaría relacionado con la profusión de algas filamentosas que ofrecen un sustrato propicio para los macroinvertebrados (Shannon et al. 1994; Downes et al. 1998). Esta proliferación de algas es frecuente cuando existe enriquecimiento por materia orgánica que actúa como una perturbación intermedia, aumentando la riqueza esperada para el lugar (Allan 1995). Sin embargo, cuando se compararon muestras del mismo tamaño usando rarefacción, la secuencia de estaciones se modificó sustancialmente. La estación RY (Jujuy) aparece como la más rica, seguida por RCh (Salta), mientras que la otra estación de Jujuy (RC) queda en último lugar.
Los grupos formados a partir de la similitud (basados en presencias y ausencias) parecen ser producidos más por particularidades locales que por una impronta de cuenca. Nuestros resultados coinciden con las afirmaciones de Downes et al. (1998), quienes sostienen que los procesos locales son importantes reguladores de las comunidades de los ríos. Los factores de control local, restringidos en nuestro caso al período entre perturbaciones (invierno) como fue observado por Fernández & Palacios (1989) y Fernández (1997), son más importantes que los regionales en correspondencia con lo propuesto por Palmer et al. (1996). Hay que destacar la independencia del ordenamiento con respecto a las cuencas, como ya fue observado por Wasson & Barrere (1999) en Bolivia.
El índice de diversidad también presenta a RY y RCh como las estaciones más diversas. Es interesante notar esta similitud en los resultados entre estas dos estaciones a partir de diferentes índices y datos de abundancia (ver MATERIALES Y MÉTODOS).
El agrupamiento obtenido en el Análisis de Correspondencia estaría relacionado, con respecto al eje I, con variables como la altitud y el estrato de vegetación asociado a cada estación. A pesar de que este análisis no discrimina dentro de un gradiente de altura de 400 m (entre los 450–850 m), sí lo hace en el rango de 800 m que existe entre RY (1625 m.s.n.m.) y RCh (800 m.s.n.m.), que es el más alto de los otros ríos. Estas variables se imponen sobre otras como las precipitaciones (factores locales) y las cuencas o la fragmentación del área de Yungas (factores regionales). Sobre el eje II, en cambio, responden a otro factor de control local como el ancho del río en la estación muestreada. Este factor es un buen descriptor de la magnitud del río y, por lo tanto, de las características del canal, cuya importancia para la comunidad bentónica ya fuera discutida, entre otros, por Vannote et al. (1980) y particularmente por Brussock et al. (1985). La magnitud de un río en un punto determinado responde en última instancia al caudal, que es el verdadero determinante de la estructura comunitaria (Allan 1995). Las cuatro estaciones reunidas en el extremo superior del gráfico (Figura 4a) comparten una serie de características que podemos resumir aquí en dos: la vegetación marginal alejada del canal (canopia abierta) y el ancho superior a 7 m.
Como ya se ha discutido en numerosas ocasiones (Covich 1988; Allan 1995), la fauna de macroinvertebrados bentónicos parece contradecir el paradigma de la tendencia al enriquecimiento específico hacia el Ecuador (Schluter & Ricklefs 1994). Si comparamos la riqueza de familias de Insecta de nuestros resultados (20 a 40) con las 36 familias obtenidas por Miserendino (1995) en un río templado de la Patagonia y con las 37 colectadas por Salinas et al. (1999) en las Yungas de Bolivia, observamos que en un rango de 26 grados de latitud no hay diferencias significativas que permitan argumentar a favor o en contra del paradigma antes mencionado.
AGRADECIMIENTOS
Agradecemos a los árbitros que revisaron este manuscrito aportando valiosas sugerencias. A Santiago Cohen por su valioso apoyo en la colecta y procesamiento de material. HR Fernández pertenece al Consejo de Investig. Cient. y Técn. de la Argentina (CONICET).
BIBLIOGRAFÍA
ALLAN, JD. 1995. Stream ecology: structure and function of running waters. Chapman & Hall. Londres, Reino Unido. 388 pp. [ Links ]
BARBOUR, MT; J GERRITSEN; GE GRIFFITH; R FRYDENBORG; E MCCARRON ET AL. 1996. A framework for biological criteria for Florida streams using benthic macroinvertebrates. J. N. Am. Benthol. Soc. 15(2):185–211. [ Links ]
BIANCHI, A & C YAÑEZ. 1992. Las precipitaciones en el Noroeste Argentino. 2da edn. INTA, EEA Salta. Salta, Argentina. 298 pp. [ Links ]
BOURNAUD, M; B CELLOT; P RICHOUX & A BERRAHOU. 1996. Macroinvertebrate community structure and environmental characteristics along a large river: congruity of patterns for identification to species or family. J. N. Am. Benthol. Soc. 15(2):232–253. [ Links ]
TER BRAAK, CJF. 1990. Update notes CANOCO version 3.10. Agricultural Mathematic Group. Wageningen, Países Bajos. 35 pp. [ Links ]
BRUSSOCK, PP; AV BROWN & JC DIXON. 1985. Channel form and stream ecosystem models. Wat. Res. Bull. 21(5):859–866. [ Links ]
CORIGLIANO, MC & AG FREYTES. 1994. Efecto de la confluencia de dos arroyos de diferente orden sobre el bentos. Ecología Austral 4:87–94. [ Links ]
CORIGLIANO, MC & GM COSSARINI. 1984. Zoobentos de arroyos inmisarios del embalse Río Tercero con especial referencia a la zonación del Río Grande (Calamuchita, Prov. Córdoba, Argentina). Ecosur 11(21/22):85–93. [ Links ]
COVICH, AP. 1988. Geographical and historical comparison of Neotropical stream: biotic diversity and detrital procesing in highly variable habitat. J. N. Am. Benthol. Soc. 7(4):361–386. [ Links ]
DOWNES, BJ; PS LAKE; ESG SCHREIBER & A GLAISER. 1998. Habitat structure and regulation of local species diversity in a stony, upland stream. Ecol. Monogr. 68(2):237–257. [ Links ]
FERNÁNDEZ, HR. 1997. La taxocenosis de hidrácaros bentónicos de los ríos de las Sierras Centrales y del Noreste de la Provincia de Tucumán, Argentina. Tesis Doctoral, Universidad Nacional de Tucumán. 152 pp. [ Links ]
FERNÁNDEZ, HR & AN PALACIOS. 1989. La fauna intersticial hiporreica de dos ríos de montaña del Noroeste de Argentina. Rev. Idrobiol. 28(3): 231–246. [ Links ]
FERNÁNDEZ, HR; F ROMERO; L GROSSO; ML DE GROSSO; M PERALTA & MC RUEDA. 1995. La diversidad del zoobentos en ríos de montaña del NOA I: el río Zerda, Provincia de Tucumán, República Argentina. Acta Zool. Lilloana 43(1):215–219. [ Links ]
GAUCH, HG. 1994. Multivariate analysis in community ecology. Cambridge University Press. 298 pp. [ Links ]
GREEN, RH. 1979. Sampling design and statistical methods for environmental biologist. J. Wiley & Sons. New York, EEUU. 257 pp. [ Links ]
HURLBERT, SH. 1971. The nonconcept of species diversity: a critique and alternative parameters. Ecology 52:577–586. [ Links ]
JACOBSEN, D; R SCHULTZ & A ENCALADA. 1997. Structure and diversity of stream invertebrate assemblage: the influence of temperature with altitude and latitude. Freshwat. Biol. 38:247–261. [ Links ]
KREBS, CJ. 1989. Ecological methodology. Harper & Row. New York, EEUU. 470 pp. [ Links ]
MAGURRAN, AE. 1989. Diversidad ecológica y su medición. Ediciones Vedrà. Barcelona, España. 200 pp. [ Links ]
MARCHESE, M & IE DE DRAGO. 1992. Benthos of the lotic environments in the middle Paraná River system: transverse zonation. Hydrobiologia 237:1–13. [ Links ]
MISERENDINO, L. 1995. Composición y distribución de macrozoobentos de un sistema lótico andinopatagónico. Ecología Austral 5:133–142. [ Links ]
MORALES, J. 1996. Comunidades arbóreas no saturadas en las Yungas de Argentina. Ecología Austral 6:17–22. [ Links ]
PALMER, MA; JD ALLAN & CA BUTMAN. 1996. Dispersal as a regional process affecting the local dynamics of marine and stream benthic invertebrates. Trends Ecol. Evol. 11(8):322–326. [ Links ]
PESO, JG. 1995. Zoobentos del arroyo Urugua-i. Estudio preliminar (Misiones, Argentina). Rev. Asoc. Cienc. Nat. Litoral 26(2):21–37. [ Links ]
QUINN, JM & CW HICKEY. 1990. Characterization and classification of benthic invertebrate communities in 88 New Zealand rivers in relation to environmental factors. N. Zealand J. Mar. Freshwat. Res. 24:387–409. [ Links ]
ROMERO, F & HR FERNÁNDEZ. 2001. Abundance and Diversity of a mayfly taxocene in a South American mountain stream. Pp. 173–178 en: E Domínguez (ed.). Trends in research in Ephemeroptera and Plecoptera. Kluwer/Plenum Press. New York, EEUU. 489 pp. [ Links ]
SALINAS, G; R MARÍN; C HENRY; O FORSATI & JG WASSON. 1999. Efecto de la materia en suspensión sobre los invertebrados bénticos de los ríos de aguas claras en las Yungas de Bolivia. Rev. Bol. Ecol. 6:183–186. [ Links ]
SCHLUTER, D & RE RICKLEFS. 1994. Species diversity. An introduction to the problem. Pp. 1–10 en: RE Ricklefs & D Schluter (eds). Species diversity in ecological communities. Historical and geographic perspectives. University of Chicago Press. Chicago, EEUU. [ Links ]
SHANNON, JP; DW BLINN; LE STEVENS & JP CARDER. 1994. Trophic interactions and benthic animal community structure in a Colorado River, Arizona, USA. Freshwater Biol. 31:213–220. [ Links ]
STRAHLER, AN. 1981. Geografía física. Omega. Barcelona, España. 767 pp. [ Links ]
VANNOTE, RL; GW MINSHALL; RW CUMMINS; JR SEDELL & CE CUSHING. 1980. The river continuum concept. Can. J. Fish. Aquat. Sci. 37:130–137. [ Links ]
VIDES ALMONACID, R; HR AYARDE; GJ SCROCCHI; F ROMERO; C BOERO & JM CHANI. 1998. Biodiversidad de Tucumán y el noroeste argentino. Opera Lilloana 43:1–89. [ Links ]
WAIS, I & AA BONETTO. 1988. Analysis of the allochtonous organic matter and associated macroinvertebrates in some streams of Patagonia (Argentina). Verh. Internat. Verein. Limnol. 23(3): 1455–1459. [ Links ]
WASSON, JG & B BARRERE. 1999. Regionalización de la cuenca amazónica boliviana: las hidroecoregiones de la zona andina. Rev. Bol. Ecol. 6:111–120. [ Links ]

