










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.11 n.1 Córdoba ene./jun. 2001
TRABAJOS ORIGINALES
Body reserves and pregnancy rates of introduced red deer in Patagonia (Argentina) after a period of drought
Werner T Flueck*
Centro de Ecología Aplicada del Neuquén, Junín de los Andes, Neuquén, Argentina
* DeerLab; C.C. 176; 8400 Bariloche, Argentina. Email: deerlab@infovia.com.ar
Received: 7 September 2000;
Revised: 9 February 2001
Accepted: 7 May 2001;
Final version: 14 May 2001
ABSTRACT. Red deer (Cervus elaphus) were introduced to Patagonia in the 1920´s and have reached high densities in certain areas of the forest–steppe ecotone. Severe drought conditions during summer/autumn of 1999 were suspected to have an impact on reproduction of red deer. Necropsies of 50 adult females revealed a pregnancy rate of only 56% as compared to 98% in 1992–1998 or 100% in 1996–1997 in other population. The age distribution of pregnant females reflected the importance of not having been pregnant during the previous cycle. The primiparous 2 year-old females had a 77% pregnancy rate, whereas none of the lactating females were pregnant. Normally, a higher reproductive success occurs for middle age classes, thereby reducing the chances of becoming pregnant during stress periods which was reflected in this study in the age distribution of non-pregnant females. However, chest girths (as an index of body size) were not different for pregnant and non-pregnant females. Body fat reserves were severely depleted in most animals irrespective of reproductive status. The calf-to-adult female ratio in the following autumn was 30% as compared to 58% in 1991–1992. The ratio in the other population was 28% in the following autumn, indicating that the effect of the drought was regional. The results suggest that these red deer populations occur at densities where they can easily become food-limited through a singular environmental phenomenon such as a drought period. Although the recruitment rate was drastically reduced through such food shortage, the effect might be of short duration as the drought terminates, and red deer may continue to exert intensive pressure on the flora causing subsequent damage if population densities are not lowered through continuous hunting or other control means.
RESUMEN. Reservas corporales y tasa de preñez del ciervo colorado introducido en Patagonia (Argentina) después de un período de sequía: El ciervo rojo (Cervus elaphus) fue introducido en Patagonia en la década de 1920 y ha alcanzado densidades altas en ciertas áreas del ecotono entre el bosque y la estepa. Se especuló que las condiciones de severa sequía del verano-otoño de 1999 podían tener un efecto negativo sobre la reproducción del ciervo rojo. Las necropsias de 50 hembras adultas revelaron una tasa de preñez de solo 56%, comparado con un 98% en 1992–1998 o un 100% en 1996–1997 en otra población similar. La distribución de edades de hembras preñadas reflejó la importancia de no haber gestado durante el ciclo anterior. Las hembras primíparas de dos años tuvieron una tasa de preñez de 77%, mientras que ninguna de las hembras con crías lactantes estaba preñada. Normalmente, el mayor éxito reproductivo se observa en las clases de mediana edad, lo que reduce la probabilidad de preñez durante periodos de stress, lo cual se reflejó en este estudio en la distribución de edades de las hembras no-gestantes. Sin embargo, la circunferencia a la altura del pecho (como índice del tamaño corporal) no fue diferente entre las hembras gestantes y las no gestantes. Las reservas de grasa corporal estuvieron severamente agotadas en la mayoría de los animales, independientemente de su condición reproductiva. La relación entre crías y hembras adultas en el otoño siguiente fue de 30%, en comparación con 58% en 1991–1992. En la otra población, la relación fue de 28% en el otoño siguiente, indicando que el efecto de la sequía tuvo carácter regional. Los resultados sugieren que estas poblaciones de ciervo rojo tienen densidades a las cuales pueden experimentar fácilmente limitaciones de tipo nutricional debido a un fenómeno ambiental singular como un período de sequía. Aunque la tasa de reclutamiento se redujo drásticamente a través de la escasez de comida, el efecto podría ser temporario, hasta terminar la sequía, y los ciervos rojos podrían continuar ejerciendo una presión intensiva sobre la vegetación si no disminuyen las densidades de la población a través de la caza regular u otra forma de control.
INTRODUCTION
Red deer (Cervus elaphus) were first introduced from Europe to central Argentina in 1906, and from there to the Andean cordillera by 1924. The population (and the occupied range) in this mountainous region of Patagonia has increased since that time. Even though red deer have invaded several National Parks and have reached high densities in several locations (Flueck et al., pers. obs.), there have been few studies to date. Although the history of initial introductions of red deer in Patagonia is fairly well known (reviewed in Flueck & Smith-Flueck 1993), little information is available on subsequent developments. In addition to some local introductions, deer have also invaded Chile from several Argentine populations (Ortiz 1992) and thus red deer represent continuous populations across the Andes (Flueck et al. 1995). In the southern cone of South America, red deer are able to survive in dense rain forests, in ecotones, but also in the dry Patagonian steppe (Flueck et al. 1995). The negative impact of red deer on plant communities has been described within some habitats along this precipitation gradient (Veblen et al. 1989, 1992).
Although there are diverse views about natural principles of population regulation in cervids (Flueck 2000), the history of this red deer invasion points towards a lack of effective factors resulting in a top–down regulation. Rather, several indices, including results from this study, provide evidence that red deer experience a bottom–up regulation through food deprivation in the Patagonian environment. Undernourishment has been shown previously to affect reproductive performance of red deer (Staines 1978; Albon et al. 1986) and the likely underlying principles have been described (Flueck 1994). Thus, lacking effective extrinsic population regulation, red deer in Patagonia may be subjected to density-depended food deprivation and its sequel. I hypothesized that the Nahuel Huapi red deer population was close to being affected by density–dependent processes and the episode of an intense drought permitted to evaluate if red deer were subsequently responding in a density–dependent manner. Prior to this study there was no ecological information available on reproductive parameters of red deer to evaluate their invasive potential in this environment, particularly in relation to unusual environmental conditions.
STUDY AREA
The study area is in the Nahuel Huapi National Park (40°58'S; 71°12'W), province of Neuquén, Argentina. The topography is primarily mountainous with most features formed by glacial processes. The majority of soils originated from volcanic processes and are young. Towards the east, the volcanic material has been modified by fluvial and cofluvial transportation of material. The dominant climate is temperate with main precipitation occurring between April and September. There is an abrupt precipitation gradient from west to east due to the rain shadow effect of the Andes, which results in a strongly defined vegetation structure and floristic composition. The study site is between 900 and 1200 m elevation and represents the ecotone between forests and steppe (Mermoz & Martin 1986). Patches of forests are characterized by "ñire" (Nothofagus antarctica) and "ciprés" (Austrocedrus chilensis) at lower elevations, and are replaced by "lenga" (Nothofagus pumilio) at higher elevations. Forest patches at lower elevation alternate with wet grasslands ("mallines"), with abundant growth of herbaceous plants. At high elevation, they are replaced by grass-dominated steppe of "coirón amargo" (Stipa speciosa var. major) and "coirón dulce" (Festuca pallescens), with variable occurrence of shrub species like "neneo" (Mulinum spinosum), "calafate" (Berberis spp.), and "espino negro" (Colletia spinosissima). Riparian areas also contain galleries of tree like "radal" (Lomatia hirsuta), "maitén" (Maytenus boaria), and "laura" (Schinus patagonicus). Another population studied (10 km further east and 50 km further north) occupied a site broadly similar with respect to topography, altitude and habitat structure. Both sites have summer range habitat dominated by N. pumilio, whereas A. chilensis and extensive grasslands dominate at lower elevations.
CLIMATIC CONDITIONS
Precipitation of 415 mm between September 1996 and May 1997 was below the recorded average of 454 mm (1981–1996), and was followed by the extremely dry and hot spring-summer of 1998–1999, having been referred to as the drought of the century in much of Patagonia, including Chile. The relative effect of the drought is presumed to have been similar within the extent of the study site. Therefore, the weather station of INTA (41°07'S; 71°15'W; elevation 775 m) is considered representative of the climatic conditions for the area (C. Bustos, INTA EEA Bariloche, pers. comm.), and data collected regularly since 1981 have been analyzed.
The period of summer through winter 1998 (January–August) was substantially dry, with only 319.5 mm of precipitation as compared to the averaged 663.2 mm received during 1981–1997. This drought continued through the following spring 1998 and summer–autumn of 1999, producing only 145 mm of precipitation as compared to the average of 454 mm for 1981–1996 (Figures 1a and 1b). Furthermore, unusually low humidity (September 1998–April 1999 below 95% CI of previous 16 years) and very hot temperatures (Figures 1c and 1d) accompanied the dry period from spring 1998 through the following autumn. Hence, the pattern of the temperatureprecipitation relationship was very different as compared to averages of the climatic conditions for the past two decades (Figure 2). The substantially higher temperature in 1998–1999 resulted in an increased potential evapotranspiration rate (Ricklefs 1979), and the water deficit was effectively higher than would be indicated by the amounts of precipitation. Thus, after correcting for these temperature differences, the period of October 1998 to April 1999 received only 17% of precipitation as compared to the average of 1982–1997. Water stress was so intense that there was a high mortality rate among evergreen conifers and broad-leafed trees of all age classes, including old growth trees like Nothofagus dombeyi (pers. obs.).
Figure 1. Precipitation and temperature data for the studied area in Nahuel Huapi National Park. In all cases, mean values (and the 95% confidence interval) for the period indicated (open circles), and values for 1998–1999 (black triangles) are shown. (a) Monthly precipitation. (b) Accumulated precipitation. (c) Monthly average temperature. (d) Soil temperature at 10 cm depth.
Figura 1. Datos de precipitaciones y temperatura para el área de estudio en el Parque Nacional Nahuel Huapi. En todos los casos, se muestran tanto los valores promedio (y su IC del 95%) para el periodo señalado (círculos blancos) como los valores correspondientes a 1998–1999 (triángulos negros). (a) Precipitaciones mensuales. (b) Precipitaciones acumuladas. (c) Temperaturas promedio mensuales. (d) Temperaturas del suelo a 10 cm de profundidad.

Figure 2. Climatic diagram showing monthly average temperature and monthly total precipitation for October through April (spring–autumn) in the studied area in Nahuel Huapi National Park. Data for 1982–1997 (open circles) and for 1998–1999 (black triangles) are shown. The tail of the arrow represents October, whereas the head represents April.
Figura 2. Diagrama climático mostrando las temperaturas promedio mensuales y las precipitaciones totales mensuales desde octubre hasta abril (primavera–otoño) en el área de estudio en el Parque Nacional Nahuel Huapi. Se muestran los datos de 1982–1997 (círculos blancos) y los de 1998–1999 (triángulos negros). La cola de la flecha representa a octubre y la punta a abril.
METHODS
Female red deer were examined during the last trimester of gestation, mainly during the months of October and November. Adult females (2 years or older) were approached by stalking and collected at first sight using a rifle without regard for age or size (n = 50). All females were part of social groups, the smallest groups consisting of a female with a calf of the previous year and a yearling female. The time and location were noted, several morphometric measurements (Mitchell et al. 1976) were taken and the animal was examined for lactation status. Chest girth was used as an index to body size (Millspaugh & Brundige 1996). Subsequently, a necropsy was performed to measure fat reserves and determine the reproductive status. A female was considered primiparous according to the physiognomy of the uterus and udder. Rump fat was measured as the deepest layer along an incision placed at a 45-degree angle, beginning at the base of the tail (Kistner et al. 1980). Brisket fat was measured as the deepest layer along a mid-central incision over the sternum, beginning at the xyphoid process (Austin 1984). Omental fat was estimated as a percentage coverage by fat of the membrane (Dauphine 1976), and the kidney fat index was calculated as fat mass divided by kidney mass times 100 (Anderson et al. 1990). Females without fetus were examined carefully to determine if it was due to not being pregnant or due to parturition, and the pregnancy rate was compared to rates from previous years in the same study area. Pregnancy rates were evaluated with regard to age and lactation status of yearling and older females. Reproductive performance of another population in a similar site was contrasted to the present results. For females collected in November 1996 (n = 19) and not pregnant, lactation was indicative of previous pregnancy, whereas females collected in May 1997 (n = 23) provided information on pregnancy rates as well as breeding history of the preceding season by the state of lactation and physiognomy of the udder. Herd composition counts were conducted during autumn using 20–40x spotting scopes from various vantage points to classify animals as either adult female, calf or male.
Age estimation was done by comparing tooth eruption and tooth wear patterns to a reference set of known-aged jaws from 1 to 18 years obtained from deer from the same area. The set was prepared using cementum annuli analysis performed in Matson´s laboratory (n = 99). This method has been shown to determine ages correctly (>97%) in Cervus elaphus aged 2–15 years (Keiss 1969, Hamlin et al. 2000). Age estimation of deer jaws by eruption-wear criteria without using a reference set can be very misleading. Hamlin et al. (2000) concluded that possible reasons for the degree of error observed among supposedly experienced personnel included less than ideal conditions during examination at hunter check stations (like working in inclement weather), that most personnel were volunteering their time, and training of personnel was based on example "jawboards" leaving the worker with a predominantly mental visual model for subsequent work. Under these conditions, each jaw was thus examined individually, dissociated from the rest, under field conditions, and only compared to the mental visual model. Age determination under field conditions without a known-age reference set has been noted to result in high error rates elsewhere (Keiss 1969, Gilbert & Stolt 1970). To circumvent these potential sources of errors in this study, age estimation of jaws was performed in the laboratory after all jaws had been collected, using the known-aged set as a reference. Jaws were first grouped according to determined age and subsequently each group was re-evaluated comparatively. Donnelly (1997) found that tooth wear is similar among red deer of the same age. Furthermore, the age classes up to 4.5 years old can readily be distinguished without error, particularly using comparative material.
Comparisons of means were done using the two-sample t test, and frequencies were analyzed using the Fisher's Exact test (at α = 0.05). Curve fitting was done using least squares estimation.
RESULTS
Several biological parameters were similar in pregnant and non-pregnant females. Chest girths as an index to body size ranged from 108 to 136 cm (P = 0.31), and the median ages of 5.6 (SE = 0.6) years in pregnant (n = 28) and 7.6 (SE = 1.0) years in non-pregnant females (n = 22) were also similar (P = 0.08). However, the frequencies in age classes were different among the two groups, following an exponential distribution in pregnant females (Chi-square goodness of fit, χ2 = 24.6; d.f. = 2; P < 0.001) and a gamma distribution in non-pregnant females (α = 2.6; β = 0.34; Kolmogorov = 0.17).
Body condition measurements indicated that the females were severely undernourished in 1999. Of several fat reserves evaluated, they were very low for all females, reaching on average only 0.1 mm of rump fat, 0.44 mm of brisket fax, and 0.28% of omental fat, irrespective of reproductive status. Thus, when comparing females (n = 19) collected right around parturition in 1996 in the other population (95% lactating, 5% still pregnant; Flueck & Smith-Flueck, pers. obs.) with females collected 4–6 weeks before parturition in 1999, the latter had comparatively much less fat reserves: 17% of brisket, 7% of omentum, 4% of rump, and 1% of kidney fat, respectively (Figure 3). These differences would have been more pronounced if indices in 1999 were measured right around parturition, as was done in 1996. Lactation in 1999 was significantly more prevalent in non-pregnant (32%) than pregnant females (0%) (Fisher´s Exact test, P = 0.005).

Figure 3. Body fat reserves of adult red deer females around parturition in Nahuel Huapi National Park during 1996 (open bars) and 1999 (black bars). Values shown are means (+ 95% CI) for rump and brisket fat (both in mm), and for omentum and kidney fat (both in percentage).
Figura 3. Reservas de grasa corporal de hembras adultas de ciervo rojo en el Parque Nacional Nahuel Huapi durante 1996 (barras blancas) y 1999 (barras negras). Se muestran los promedios (+ IC del 95%) de grasa en ancas y en pecho (ambos en mm) y de grasa en omentum y en riñón (ambos en porcentaje).
The nutritional stress as evidenced by the depleted fat reserves affected the pregnancy rate of females in 1999. For instance, no yearlings were found pregnant and only 77% (n = 13) of 2 year-olds conceived. In contrast, pregnancy rates in 1996–1997 in the other population were 9% (n = 11) for yearlings, 100% (n = 8) for 2 year-olds and 100% (n = 39) for older females (Table 1; Flueck & Smith-Flueck, pers. obs.). Females older than 3 years conceived at a rate of merely 51% (n = 35) in 1999, as opposed to 100% (n = 31) in 1996–1997. All lactating adult females in 1997 (n = 12) were pregnant, whereas no lactating adult females were pregnant in 1999 (n = 7). However, in 1999 even non-lactating adult females (n = 43) conceived at a low rate of 65% as opposed to 100% (n = 9) in 1997. There were no indications of reabsorption or abortion of fetuses in non-pregnant females.
Table 1. Breeding history of adult red deer females in Nahuel Huapi National Park. Values are number of individuals (with percentage in brackets).
Tabla 1. Estado reproductivo de hembras adultas de ciervo rojo en el Parque Nacional Nahuel Huapi. Los valores corresponden al número de individuos (con el porcentaje entre paréntesis).
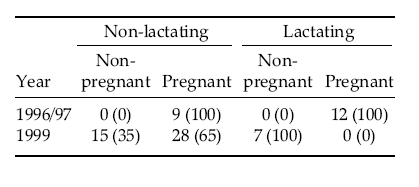
Pregnancy rates from 1999 could be compared to data obtained on the same population between 1992–1998. Thus, the pregnancy rate of 56% among adult deer in 1999 contrasted with the 98% (n = 88) obtained before (Flueck, pers. obs.). Furthermore, whereas 29% of yearlings were pregnant in previous years, none were found pregnant in 1999.
Herd composition data from the Nahuel Huapi red deer population further corroborate the unusual circumstances of 1998–1999. The calf-to-adult female ratio in autumn of 1991–1992 combined was 58% (n = 1242), whereas in autumn of 2000 it was merely 30% (n = 472). In comparison, herd counts in autumn of 2000 in the other population revealed a ratio of 28% (n = 364) calves.
DISCUSSION
Red deer have been shown earlier to occur at high densities in this region, reaching 100 deer/km2 in ecotonal habitat and 50 deer/km2 in open steppe (Flueck et al., pers. obs.). The prevailing climatic conditions of 1998–1999 were responsible for the observed response by the red deer population by reducing the pregnancy rate of adult deer to a low of 56% as compared to previous years. Pregnancy rates, particularly of yearlings and 2 year-olds, were also strongly affected. The relatively high pregnancy rate of 2 year-olds in 1999 compared to older females is likely explained by the fact that they were primiparous and, therefore, they were in better condition as compared to females that had been pregnant in the previous season. Conceivably, for the same reasons, none of the females that were still lactating became pregnant. Other studies found that lactating females in high density red deer populations (i.e., limited by nutrition) will conceive at rates as low as 40% (Clutton-Brock et al. 1982:264). On the other hand, the age distribution of non-pregnant females can be interpreted as an indication of the normally higher reproductive success of middle age classes, and likely most of these females had produced a calf during the previous year. Chest girth, as an index of body size, did not differ among pregnant and non-pregnant females, and the most likely factor determining pregnancy was body condition with respect to fat reserves during the rut (Flueck 1994). There were no cases of resorption of fetuses or abortions, in agreement with observations by Mitchell et al. (1986). Of 1471 females examined, they found no signs of resorption or abortion, although only 773 females were pregnant due to undernourishment, concluding that females either conceived and had a calf or did not conceive at all. Observation during the rut in the autumn of 1999 corroborated the subsequent low pregnancy rates in the present study. Comparing use of rutting places with previous years, there were fewer females involved in the rut of 1999. For instance, while a certain place had on average 10 stags in all years, the number of females dropped from a usual 25 to 3 in 1999, and there was more than usual fighting activities (Flueck, pers. obs.). There were also two marked peaks indicating that a substantial number of females conceived with a delay. A further coincidental observation was a male calf collected in spring with absence of antler pedicle growth. This has been related to undernourishment (Raesfeld 1988:92), as may have been caused by the drought of 1998–1999. The population structure in autumn of 2000 revealed an unusually low ratio of calf to adult females compared to 1991–1992 (30% vs. 58%), corroborating the low pregnancy rates of 1999. Furthermore, a similarly low calf ratio of 28% in the other population indicated that the phenomenon likely was regional in extent. The results indicate that these red deer populations occur at densities where they can become easily food-limited through singular climatic phenomenon such as a drought period (see Anthony 1976; Frank & McNaughton 1992). As the drastic reduction of the recruitment rate through food shortage was temporary, due to the drought, red deer numbers will likely increase again under more favourable environmental conditions. Therefore, they might continue to exert intensive pressure on the flora, with associated damage to plant, unless population densities are lowered through continuous hunting or other control means.
In conclusion, the drought of the century in Patagonia occurring in 1998–1999 resulted in practically an absence of fat reserves in autumn in most red deer females irrespective of reproductive status. Consequently, the pregnancy rate of adult females dropped by nearly half and no yearlings conceived as compared to previous years. The age distribution of pregnant females reflected the importance of not having been pregnant during the previous cycle. The red deer population occurred at a density where it became food-limited through a singular environmental phenomenon such as a drought period. The study revealed that red deer in Patagonia lacked effective intrinsic population regulation and were subjected to density-depended processes working on the reproductive rate through food deprivation.
ACKNOWLEDGMENTS
I thank several landowners for their interest and support of this study, as well as the Administración de Parques Nacionales from Argentina for supporting the study within park boundaries. Also thanks to G. D. Elliott and R. Pribish for assistance with fieldwork, and three anonymous reviewers for especially constructive advice. This work was done as member of the CIC of the Consejo de Investigación Científica y Técnica de la Argentina.
REFERENCES
ALBON, SD; B MITCHELL; BJ HUBY & D BROWN. 1986. Fertility in female red deer (Cervus elaphus): the effects of body composition, age and reproductive status. J. Zool. 209:447–460. [ Links ]
ANDERSON, AE; DC BOWDEN & DE MEDIN. 1990. Indexing the annual fat cycle in a mule deer population. J. Wildl. Manage. 54(4):550–556. [ Links ]
ANTHONY, RG. 1976. Influence of drought on diets and numbers of desert deer. J. Wildl. Manage. 40(1):140–144. [ Links ]
AUSTIN, DD. 1984. Fat depth at the xiphoid process— a rapid index to deer condition. Great Basin Nat. 44(1):178–181. [ Links ]
CLUTTON-BROCK, TH; FE GUINNESS & SD ALBON. 1982. Red Deer: behavior and ecology of two sexes. The University of Chicago Press. Chicago, USA. 378 pp. [ Links ]
DAUPHINE, T. 1976. Biology of the Kaminuriak population of Barren Ground Caribou. Part 4. Growth, reproduction and energy reserves. Can. Wildl. Serv. Rep. Series 38:1–71. [ Links ]
DONNELLY, PJ. 1997. Age determination from the jaws of adult deer. Deer 10(4):233–235. [ Links ]
FLUECK, WT. 1994. Relationship between body weight, lipid reserves, and ovulation rate in non-equatorial cervids: a hypothesis. Europ. J. Wildl. Res. 40:12–21. [ Links ]
FLUECK, WT. 2000. Population regulation in large northern herbivores: evolution, thermodynamics, and large predators. Europ. J. Wildl. Res. 46:139–166. [ Links ]
FLUECK, WT & JM SMITH-FLUECK. 1993. Über das in Argentinien angesiedelte Rotwild (Cervus elaphus, L., 1758): verbreitung und tendenzen. Europ. J. Wildl. Res. 39:153–160. [ Links ]
FLUECK, WT; JM SMITH-FLUECK & KA RÜEGG. 1995. Management of introduced red deer in Patagonia. Pp. 525–528 in: JA Bissonette & PR Krausman (eds). Integrating people and wildlife for a sustainable future. The Wildlife Society. Bethesda, USA. [ Links ]
FRANK, DA & SJ MCNAUGHTON. 1992. The ecology of plants, large mammalian herbivores, and drought in Yellowstone National Park. Ecology 73(6):2043–2058. [ Links ]
GILBERT, FF & SL STOLT. 1970. Variability in aging Maine white-tailed deer by tooth-wear characteristics. J. Wildl. Manage. 34(3):532–535. [ Links ]
HAMLIN, KL; DF PAC; CA SIME; RM DESIMONE & GL DUSEK. 2000. Evaluating the accuracy of ages obtained by two methods for montane ungulates. J. Wildl. Manage. 64(2):441–449. [ Links ]
KEISS, RE. 1969. Comparison of eruption-wear patterns and cementum annuli as age criteria in elk. J. Wildl. Manage. 33(1):175–180. [ Links ]
KISTNER, TP; CE TRAINER & NA HARTMANN. 1980. A field technique for evaluating physical condition of deer. Wildl. Soc. Bull. 8(1):11-17. [ Links ]
MERMOZ, M & C MARTIN. 1986. Mapa de vegetación del parque y la reserva Nacional Nahuel Huapi. Administración de Parques Nacionales, Delegación Regional Patagonia. Bariloche, Argentina. [ Links ]
MILLSPAUGH, JJ & GC BRUNDIGE. 1996. Estimating elk weight from chest girth. Wildl. Soc. Bull. 24(1):58– 61. [ Links ]
MITCHELL, B; D MCCOWAN & IA NICHOLSON. 1976. Annual cycles of body weight and condition in Scottish red deer, Cervus elaphus. J. Zool. 180:107–127. [ Links ]
MITCHELL, B; D MCCOWAN & T PARISH. 1986. Performance and population dynamics in relation to management of red deer, Cervus elaphus, at Glenfeshie, Inverness-Shire, Scotland. Biol. Conserv. 37:237–267. [ Links ]
ORTIZ, CR. 1992. Current status of introduced red and fallow deer populations in Chile: the need of management. Pp. 30–36 in: RD Brown (ed.). The biology of deer. Springer Verlag. New York, USA. [ Links ]
RAESFELD, FV. 1988. Das rotwild. 9th edn. Verlag Paul Parey. Hamburg, Germany. 416 pp. [ Links ]
RICKLEFS, RE. 1979. Ecology. Chiron Press. New York, USA. 966 pp. [ Links ]
STAINES, BW. 1978. The dynamics and performance of a declining population of red deer (Cervus elaphus). J. Zool. 184:403–419. [ Links ]
VEBLEN, TT; M MERMOZ; C MARTIN & E RAMILO. 1989. Effects of exotic deer on forest regeneration and composition in northern Patagonia. J. Appl. Ecol. 26:711–724. [ Links ]
VEBLEN, TT; M MERMOZ; C MARTIN & T KITZBERGER. 1992. Ecological impact of introduced animals in Nahuel Huapi National Park, Argentina. Conserv. Biol. 6:71–83. [ Links ]

