










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.11 n.1 Córdoba ene./jun. 2001
TRABAJOS ORIGINALES
Tendencias espaciales de la regeneración de Austrocedrus chilensis en relación con el dosel arbóreo
Marcelo F Arturi*, Juan J Ferrando, Juan F Goya, Pablo F Yapura & Jorge L Frangi
Laboratorio de Investigación de Sistemas Ecológicos y Ambientales, Facultad de Ciencias Agrarias y Forestales y Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Buenos Aires, Argentina
* Lab. Invest. de Sistemas Ecológicos y Ambientales, Univ. Nac. La Plata; CC 31; 1900 La Plata, Buenos Aires, Argentina. Email: talares@ceres.agro.unlp.edu.ar
Recibido: 29 noviembre 2000;
Aceptado: 30 abril 2001;
Versión final: 15 junio 2001
RESUMEN. Se analizó la relación entre el tipo de dosel de bosques de Austrocedrus chilensis, la densidad de renovales (individuos menores de 5 cm de DAP) de dicha especie y sus tendencias espaciales. Se determinó la densidad de renovales de Austrocedrus chilensis, agrupados en tres clases de altura (0.5–1 m, 1–2 m y >2 m), en dos sitios ubicados en áreas protegidas (El Guadal y Loma del Medio) del SO de la Provincia de Río Negro. Se registró el número de individuos de cada clase, así como la presencia-ausencia de las especies arbóreas del dosel. Las unidades muestrales se ordenaron mediante Análisis de Coordenadas Principales a partir de los datos de presencia–ausencia de especies. Los ejes definidos por dicho análisis se consideraron como una nueva variable que definió el tipo de dosel. Se analizó la correlación entre la densidad de los renovales y un índice de competencia que cuantifica el grado de ocupación de las parcelas por individuos adultos (>5 cm DAP). Se analizó la tendencia espacial de la densidad de cada clase de altura y de la variable tipo de dosel. Se utilizaron regresiones polinomiales sobre las coordenadas espaciales x e y definidas como los lados de un cuadro de 50 x 50 m en cada sitio. La mayor densidad de renovales de la clase de mayor altura tendió a ubicarse en sitios sin cobertura de adultos de la propia especie. Los individuos de la categoría menor presentaron una distribución independiente del tipo de dosel, pero se asociaron positivamente a los sitios con mayor ocupación por ejemplares adultos en ambos sitios. El crecimiento de los renovales presentó una fuerte asociación positiva con su altura. Los renovales más altos presentaron tendencias espaciales bien definidas. En cambio, no se detectaron tendencias espaciales en el tipo de dosel. Los renovales de mayor crecimiento se establecieron hace más de 40 años, y el tipo de dosel de los sitios probablemente ha cambiado desde entonces hasta el momento del muestreo. Las condiciones actuales de cobertura podrían no ser favorables a la regeneración, por lo que no resulta posible predecir el éxito de la regeneración a partir de esa variable.
ABSTRACT. Spatial trends and tree canopy incidence in Austrocedrus chilensis regeneration: We analyzed the relationships among the type of canopy in Austrocedrus chilensis forests, the density of saplings (individuals <5 cm DBH) and their spatial trends. We classified saplings into three height classes (0.5–1 m, 1–2 m, and >2 m), and we measured its densities in two sites, each one corresponding to a protected area (El Guadal and Loma del Medio) located in SW Río Negro Province, Argentina. We recorded the number of Austrocedrus chilensis individuals in each class. Also, we annotated the presence–absence of tree species in the canopy, defining canopy types, in each sample unit. Canopy type data was employed for sample units–ordination through Principal Coordinates Analysis. We performed a correlation analysis between sapling density and a competition index that quantifies the degree of site occupation by adult individuals (>5 cm DBH). Additionally, we analyzed the spatial trend of the density of each sapling height class and of the type of canopy variable obtained by multivariate analysis. We used polynomial regressions on x and y spatial coordinates defined as the sides of a 50 x 50 m quadrat in each site. The highest density of saplings of the tallest class preferred sites without canopy of the same species. Sapling individuals of the lowest class were distributed independently of the canopy type, but were positively associated to sites with a larger occupation by adults in both sites. Sapling height growth was strongly and positively associated to its height. Taller saplings showed well–defined spatial trends. However, we did not detected spatial trends in the canopy type. Saplings with greater growth established more than 40 years ago, and the canopy type of the sites probably changed from that time until our sampling date. The canopy type observed would be not favorable to regeneration and, then, it is not reliable to predict the recruitment success taking into account the actual canopy condition.
INTRODUCCIÓN
Diversos trabajos indicaron que el grado de cobertura y la composición del dosel forestal condicionan a la regeneración de las especies arbóreas (Bray 1956; Augspurger 1984; Brokaw 1985; Marquis et al. 1986; Sork 1988; Lieberman et al. 1989; Whitmore 1989; Ellison et al. 1993). Esa relación entre las características del dosel y la regeneración podría conducir a la existencia de patrones espaciales semejantes en ambas variables.
En zonas mésicas, la regeneración de Austrocedrus chilensis (D. Don) Florin et Boutleje se produce masivamente después de incendios (Veblen & Lorentz 1987). Este proceso conduce al desarrollo de rodales coetáneos monoespecíficos o mixtos (Gobbi & Sancholuz 1992; Veblen et al. 1995). Los bosques menores de 150 años presentan escasa regeneración debido al efecto de la cobertura del dosel (Veblen et al. 1992). Debido a que Austrocedrus chilensis se establece en etapas tempranas de la sucesión, es probable que su regeneración se vea afectada por la presencia de un dosel arbóreo tanto de la propia especie como de otras. El estudio de la relación entre el tipo de dosel y la abundancia y distribución espacial de los renovales permitiría comprender la expresión espacial del proceso de regeneración y sería aplicable a tratamientos silvícolas para manejar la regeneración natural. El objetivo de este trabajo es poner a prueba la hipótesis de que las variaciones en el tipo de dosel arbóreo inciden sobre la densidad y distribución espacial de los renovales y que esa incidencia es diferente para distintas clases de tamaño de los renovales.
MATERIALES Y MÉTODOS
Área de estudio
El trabajo se llevó a cabo en la Reserva El Guadal y en la Reserva Provincial Loma del Medio, ubicadas en las cercanías de la localidad de El Bolsón (41°56'S; 71°33'O). La temperatura media anual del área es de 9.3 °C y la precipitación de 904 mm anuales, con una ocurrencia marcadamente invernal, registrándose períodos de déficit hídrico durante el verano. Los dos sitios estudiados se encuentran libres de pastoreo por ganado doméstico. La estructura del bosque fue estudiada en ambos sitios (Goya, obs. pers.) y existen dos parcelas permanentes de 50 x 50 m subdivididas en cuadros de 10 x 10 m. Ambos constituyen rodales maduros post–incendio con edades promedio de 80 y 75 años, respectivamente. Sólo en Loma del Medio se realizaron intervenciones forestales. Se realizó una corta de saneamiento aproximadamente 10 años antes del muestreo de este trabajo. Considerando individuos mayores de 5 cm de diámetro a la altura del pecho, El Guadal presenta una densidad de 1428 ind/ha y un área basal de 40.8 m2/ha; en Loma del Medio esos valores son de 620 ind/ha y 23.6 m2/ha, respectivamente. Ambos sitios difieren marcadamente en las características de los suelos. El Guadal presenta suelos con mayor proporción de arena que Loma del Medio y, consecuentemente, mayor drenaje y menor capacidad de retención de agua. Esto determina diferencias ambientales relevantes considerando que la estación seca ocurre durante el verano.
MÉTODOS
Se determinó la densidad de individuos de Austrocedrus chilensis menores de 5 cm de DAP (diámetro a 1.3 m de altura) separados en tres categorías de tamaño: 0.5–1.0 m de altura (I), 1–2 m de altura (II) y >2 m de altura y <5 cm de DAP (III). En ambos sitios de estudio se utilizaron parcelas cuadradas de 16 m2 ubicadas en los vértices y en el centro de los cuadros de 10 x 10 m de cada una de las parcelas permanentes. El total de parcelas fue de 61 en cada sitio. En cada parcela se registró el número y la altura de los individuos de cada categoría y las especies arbóreas presentes en el dosel. Los sitios fueron comparados mediante los datos de presencia–ausencia de especies en el dosel arbóreo, utilizando un coeficiente para datos cualitativos (porcentaje de desacuerdo, Digby & Kempton 1987). A partir de la matriz de similitud se ordenaron los sitios mediante un Análisis de Coordenadas Principales (Digby & Kempton 1987). Los ejes calculados representaron una variable teórica que caracterizó el tipo de dosel en cada parcela. Los ejes fueron correlacionados con la densidad de cada una de las categorías de individuos mediante el Coeficiente de Correlación por Rangos de Spearman (Steel & Torrie 1992).
Las densidades de los renovales de las tres categorías fueron comparadas con un índice de competencia calculado para cada una de las 61 parcelas. Dicho índice es la suma de los cocientes área basal/distancia de todos los árboles ubicados en un radio de 5 m en torno del centro de cada parcela. Este índice refleja la ocupación por ejemplares adultos en torno a cada parcela mediante una medida de la cantidad y tamaño de los individuos y la distancia al centro de la parcela. Los datos para los cálculos de los índices provinieron de los estudios previos sobre estructura (Goya, obs. pers.). Se calcularon correlaciones por rangos entre el índice de competencia y la densidad de renovales de cada categoría.
Se colectaron 75 y 110 renovales en El Guadal y Loma del Medio, respectivamente, a los que se determinó la edad y se calculó el incremento promedio anual en altura. Se calcularon regresiones simples de la edad y el crecimiento en altura sobre la altura en cada sitio.
Se utilizaron regresiones polinomiales con las coordenadas espaciales (Gittins 1968; Legendre & Fortin 1989; Cressie 1991) para describir tendencias en la distribución espacial de la densidad y el tipo de dosel. Se evaluaron mediante el ajuste de polinomios de segundo orden utilizando las coordenadas espaciales x e y como variables independientes. El orden de los polinomios se estableció en función de la posibilidad de interpretar los resultados como tendencias lineales y cuadráticas. Si bien los polinomios de orden mayor pueden lograr mejores ajustes, la interpretación de los términos es más dificultosa. Se adoptaron valores conservativos debido a que el tipo de datos utilizado suele presentar dependencia espacial (Cressie 1991). Además, los residuos de los polinomios ajustados a datos espaciales suelen presentar autocorrelación, especialmente cuando se utilizan polinomios de bajo orden (Cressie 1991). Ambos problemas aumentan la tasa de error Tipo I, por lo que se justifica la utilización de valores bajos de a para evaluar la existencia de tendencias.
RESULTADOS
En El Guadal se registró dosel de Austrocedrus chilensis en la mayor parte de las parcelas (46 sobre 61), mientras que 11 no tuvieron cobertura de dosel. Ninguna de las otras especies presentó un valor de frecuencia mayor de 8. Sólo cuatro parcelas presentaron un dosel arbóreo en el que Austrocedrus chilensis no estuvo presente. El primer eje del ordenamiento separó los sitios sin presencia de cobertura arbórea de aquellos con dosel de Austrocedrus chilensis y otras especies (Tabla 1). La densidad de los renovales de la categoría mayor se correlacionó negativamente con el primer eje del Análisis de Coordenadas Principales, indicando una asociación positiva con los sitios en claros. Los renovales de las categorías I y II presentaron una distribución independiente del tipo de dosel.
Tabla 1. Correlaciones por rangos de Spearman de los valores de los ejes 1 y 2 del Análisis de Coordenadas Principales con la presencia de diferentes especies en el dosel y con la densidad de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) en la Reserva El Guadal. *: P < 0.05; **: P < 0.01.
Table 1. Spearman rank correlations of scores from the first two Principal Coordinates Analysis axes with the presence of different canopy species and with the density of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII) in El Guadal Reserve. *: P < 0.05; **: P < 0.01.

En Loma del Medio, Austrocedrus chilensis estuvo presente en el dosel en 27 de las 61 parcelas, mientras que las que no presentaron dosel arbóreo fueron 23. La frecuencia de las restantes especies no fue en ningún caso mayor de 6, habiéndose observado 11 parcelas con dosel arbóreo en las que Austrocedrus chilensis estuvo ausente. El eje 2 del Análisis de Coordenadas Principales separó sitios con dosel de Austrocedrus chilensis y/o Nothofagus antarctica (hacia el extremo positivo) de aquellos en claros o con dosel de Lomatia hirsuta (hacia el extremo negativo) (Tabla 2). Este eje se correlacionó negativamente con la densidad de los renovales de las categorías II y III, mientras que los renovales de la categoría I presentaron una distribución independiente del tipo de dosel.
Tabla 2. Correlaciones por rangos de Spearman de los valores de los ejes 1 y 2 del Análisis de Coordenadas Principales con la presencia de diferentes especies en el dosel y con la densidad de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) en la Reserva Loma del Medio. *: P < 0.05; **: P < 0.01.
Table 2. Spearman rank correlations of scores from the first two Principal Coordinates Analysis axes with the presence of different canopy species and with the density of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII) in Loma del Medio Reserve. *: P < 0.05; **: P < 0.01.

En los dos sitios el índice de competencia se correlacionó positivamente con la densidad de renovales de la categoría I, pero no se hallaron correlaciones significativas para las restantes clases de tamaño (Tabla 3).
Tabla 3. Correlaciones por rangos de Spearman entre el índice de competencia y la densidad de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) en las reservas El Guadal y Loma del Medio. *: P < 0.05; **: P < 0.01.
Table 3. Spearman rank correlations between the competition index and the density of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII) in El Guadal and Loma del Medio reserves. *: P < 0.05; **: P < 0.01.

En ambos sitios, la altura de los renovales mostró una asociación más fuerte con el crecimiento (R2 = 0.90, P < 0.001 para El Guadal; R2 = 0.86, P < 0.001 para Loma del Medio) que con la edad (R2 = 0.18, P < 0.001 para El Guadal; R2 = 0.45, P < 0.001 para Loma del Medio). La mayor parte de los renovales fueron mayores de 30 años en ambos sitios (Figura 1).
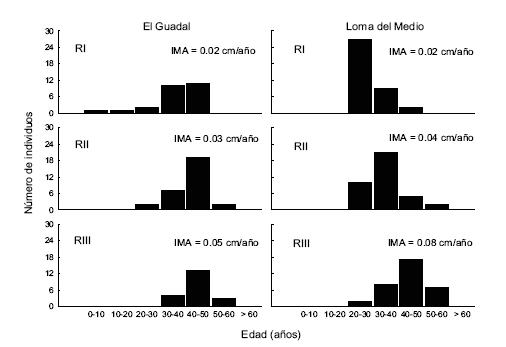
Figura 1. Distribución de frecuencias por clases de edades de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) en las reservas El Guadal y Loma del Medio. Se indica el promedio de los incrementos promedio anuales en altura (IMA) en cada caso.
Figure 1. Frequency distribution (per age classes) of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII) in El Guadal and Loma del Medio reserves. Mean annual height increment (IMA) is indicated.
El análisis de regresión indicó, en ambos sitios, tendencias espaciales en la densidad de renovales de las categorías II y III (Tabla 4). No se hallaron tendencias en los renovales menores ni en el tipo de dosel. Para los renovales de la categoría III el modelo espacial explicó el 37% (El Guadal) y el 49% (Loma del Medio) de la varianza, indicando tendencias bien definidas en la distribución de su abundancia (Figura 2). En El Guadal se hallaron relaciones lineales con el eje x y cuadráticas con el eje y. En Loma del Medio se halló una relación cuadrática con el eje y. En ambos sitios, el porcentaje de varianza explicado por los modelos espaciales de los renovales de la categoría II fue menor de 20%. En ninguno de los sitios se detectaron tendencias espaciales en el tipo de dosel.
Tabla 4. Parámetros de las regresiones polinomiales sobre las coordenadas espaciales x e y para la densidad de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) y para el tipo de dosel en las reservas El Guadal y Loma del Medio. Se indican los valores de a y b para P < 0.05. *: P < 0.05; **: P < 0.01; ***: P < 0.001; ns: no significativo; n = 61.
Table 4. Polynomial regression parameters on spatial coordinates (x, y) of the density of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII), and of canopy type in El Guadal and Loma del Medio reserves. Values for a and b for P < 0.05 are indicated. *: P < 0.05; **: P < 0.01; ***: P < 0.001; ns: not significant; n = 61.


Figura 2. Parcelas con valores superiores al percentil 66 de densidad de renovales de Austrocedrus chilensis de las tres categorías de tamaño (RI, RII y RIII) y de cobertura de dosel en las reservas El Guadal y Loma del Medio.
Figure 2. Plots with values above percentile 66 in the density of Austrocedrus chilensis saplings belonging to three size classes (RI, RII y RIII), and in the cover of canopy in El Guadal and Loma del Medio reserves.
DISCUSIÓN
La incidencia del dosel arbóreo sobre la regeneración pudo detectarse mediante una variable teórica obtenida por Análisis de Coordenadas Principales. Ninguna de las variables que definieron el tipo de dosel pudo mostrar por sí sola esa relación. La tendencia encontrada indica que los individuos mayores se encuentran principalmente en áreas sin cubierta arbórea o con un dosel de especies diferentes de Austrocedrus chilensis. Esta asociación negativa con la cubierta de la propia especie es poco pronunciada pero se observó en los dos sitios estudiados. En El Guadal se observó una tendencia negativa entre los renovales mayores y la presencia de N. antarctica, y en Loma del Medio una tendencia positiva con L. hirsuta. En ambos casos, esas relaciones se infieren por la correlación con los ejes de ordenamiento del tipo de dosel, pero no quedan claramente documentadas debido a la baja frecuencia de esas especies. La asociación negativa débil entre los renovales mayores y los sitios con cobertura de la propia especie fue el resultado más evidente en ambos sitios.
La independencia de los renovales menores respecto del tipo de dosel indica que esta variable no incide sobre la probabilidad de establecimiento, mientras que la relación positiva con el índice de competencia indica su tendencia a aparecer en sitios con alta ocupación por ejemplares adultos. Los renovales mayores presentaron una tendencia negativa, aunque no significativa, con la ocupación por ejemplares adultos. En consecuencia, los sitios con dosel de Austrocedrus chilensis y alta ocupación determinarían menor probabilidad de paso hacia clases de mayor tamaño. Las diferencias de crecimiento constituyen la principal causa de las diferencias de altura de los renovales en ambos sitios. Esto indica que las diferencias de tamaño resultan de variaciones en las condiciones de crecimiento más que de diferencias de edad.
El análisis de la tendencia espacial permitió determinar la existencia de una distribución no uniforme de los renovales de mayores tamaños. Las tendencias observadas para los renovales de la categoría III en ambos sitios presentaron altos porcentajes de varianza explicada y alta significación de los R2. Estos resultados no fueron tan categóricos en los renovales de la categoría II, indicando que su distribución es aproximadamente uniforme. Debido a que no se detectaron tendencias en el tipo de dosel, no es posible atribuir a esta variable las tendencias de los renovales mayores. Además, la falta de tendencia espacial en los renovales menores reflejaría una distribución uniforme de los propágulos, por lo que esta variable tampoco podría constituir una explicación de aquella tendencia.
La falta de congruencia entre la tendencia de la distribución espacial de los renovales y la del tipo de dosel podría deberse a cambios en la cubierta ocurridos durante el desarrollo del rodal, que habrían debilitado la asociación entre ambas variables. El establecimiento de los renovales de la categoría III ocurrió hace más de 40 años. Estos sitios podrían haberse saturado por su establecimiento y el dosel haber cambiado desde entonces. Esta interpretación es coherente con la ausencia de renovales pequeños que presenten crecimientos semejantes a los de categoría III y que podría deberse a la falta de situaciones favorables de dosel desde aquel momento hasta la actualidad. Los renovales de categoría III podrían considerarse remanentes de un proceso de regeneración por el que se ocuparon los espacios disponibles hace más de 40 años. Esos espacios habrían presentado condiciones de cobertura diferentes de las que tienen actualmente y que hicieron posible el establecimiento de los individuos que los ocuparon. Esos cambios en el dosel y la ausencia de sitios semejantes en la actualidad serían la causa de la débil relación entre esa variable y la regeneración. La tolerancia de Austrocedrus chilensis a la sombra es otra de las características que favorece la existencia de una relación débil con el tipo de dosel. Esta especie es capaz de permanecer durante muchos años en situaciones poco favorables, manteniendo un bajo crecimiento.
La ausencia de renovales menores de 25 años no indica que el proceso de su establecimiento se haya interrumpido. La tasa de crecimiento en altura de los renovales menores es de 2 cm/año, por lo que necesitan 25 años para alcanzar la altura mínima muestreada de 50 cm. Esto coincide con las observaciones de quienes señalan que las plántulas de Austrocedrus chilensis tienden a establecerse bajo la planta madre o bajo cobertura de otras especies (Gobbi & Schlichter 1998; Kitzberger et al. 2000). Sin embargo, el desarrollo posterior requiere condiciones de menor cobertura. En bosques mésicos cercanos a los estudiados se encontró que la regeneración está asociada a claros y a cambios de cobertura debidos a procesos de mortalidad en bosques de más de 150 años (Veblen 1989; Veblen et al. 1995). Desde el punto de vista del manejo, debería considerarse la posibilidad de estimular el crecimiento de los renovales mediante una reducción de la cobertura del dosel. La distribución espacial de las clases menores indica que no habría restricciones espaciales en su disponibilidad. Las limitaciones podrían estar relacionadas con la posible disminución de la capacidad de respuesta a la liberación en los individuos de mayor edad, ya que pueden permanecer durante mucho tiempo en condiciones de bajo crecimiento. Existen experiencias locales de liberación de individuos equivalentes a los renovales de la categoría III (Marcelo Rey, com. pers.). Debido a la alta relación entre la altura y el diámetro, los mismos sufrieron daños irreversibles por efecto del viento y la acumulación de nieve. Por este motivo, el manejo debería tender a estimular el crecimiento de los individuos de menor edad, que actualmente tendrían menos de 50 cm de altura. Esto podría lograrse mediante cortas de aclareos sucesivos (Smith et al. 1997).
AGRADECIMIENTOS
Este trabajo contó con el financiamiento parcial de la Universidad Nacional de La Plata. Deseamos agradecer a Sabrina Vaccaro y Cecilia Díaz por la colaboración en el trabajo de campo. A Marcelo Rey, del Servicio Forestal Andino, Provincia de Río Negro y a Ernesto Andenmatten, del Campo Forestal General San Martín, INTA.
BIBLIOGRAFÍA
AUGSPURGER, CK. 1984. Seedling survival of tropical trees species: interactions of dispersal distances, light–gaps, and pathogens. Ecology 65:1705–1712. [ Links ]
BRAY, JR. 1956. Gap phase replacement in a maple–basswood forest. Ecology 37:598–600. [ Links ]
BROKAW, NBL. 1985. Treefalls, regrowth and community structure in tropical forests. Pp. 53–69 en: STA Pickett & PS White (eds). The ecology of natural disturbance and patch dynamics. Academic Press. Nueva York, EEUU. [ Links ]
CRESSIE, NA. 1991. Statistic for spatial data. Wiley. Nueva York, EEUU. 900 pp. [ Links ]
DIGBY, PGN & RA KEMPTON. 1987. Multivariate analysis of ecological communities. Chapman & Hall. Londres, Reino Unido. 203 pp. [ Links ]
ELLISON, AM; JS DENSLOW; BA LOISELLE & D BRENÉS. 1993. Seed and seedling ecology of Neotropical melastomataceae. Ecology 74:1733–1749. [ Links ]
GITTINS, R. 1968. Trend surface analysis of ecological data. J. Ecol. 56: 845–869. [ Links ]
GOBBI, M & L SANCHOLUZ. 1992. Regeneración postincendio del ciprés de la cordillera (Austrocedrus chilensis) en los primeros años. Bosque 13:25–32. [ Links ]
GOBBI, M & T SCHLICHTER. 1998. Survival of Austrocedrus chilensis seedlings in relation to microsite conditions and forest thinning. For. Ecol. Manage. 111:137–146. [ Links ]
KITZBERGER, T; DF STEINAKER & TT VEBLEN. 2000. Effects of climatic variability on facilitation of tree establishment in northern Patagonia. Ecology 81:1914–1924. [ Links ]
LEGENDRE, P & M FORTIN. 1989. Spatial pattern and ecological analysis. Vegetatio 73:107-138. [ Links ]
LIEBERMAN, M; D LIEBERMAN & R PERALTA. 1989. Forests are not just swiss cheese: canopy stereogeometry of non-gaps in tropical forests. Ecology 70:550–552. [ Links ]
MARQUIS, RJ; HJ YOUNG & HE BRAKER. 1986. The influence of understory vegetation cover on germination and seedling establishment in a tropical lowland wet forest. Biotropica 18:273–278. [ Links ]
SMITH, DM; BC LARSON; MJ KELTY & PMS ASHTON. 1997. The practice of silviculture: applied ecology. 9th edn. John Wiley & Sons. 537 pp. [ Links ]
SORK, VL. 1988. Effects of predation and light on seedling establishment in Gustavia superva. Ecology 68:1341–1350. [ Links ]
STEEL, RG & JH TORRIE. 1992. Bioestadística: principios y procedimientos. 2da edn. McGraw–Hill. 633 pp. [ Links ]
VEBLEN, TT. 1989. Nothofagus regeneration in treefall gaps in Northern Patagonia. Can. J. For. Res. 19:365–371. [ Links ]
VEBLEN, T; B BURNS; T KITZBERGER; A LARA & R VILLALBA. 1995. The ecology of conifers of Southern South America. Pp. 120–155 en: NJ Enright & RS Hill (eds). Ecology of the Southern conifers. Melbourne University Press. Victoria, Australia. [ Links ]
VEBLEN, T; T KITZBERGER & A LARA. 1992. Disturbance and forest dynamics along a transect from Andean rain forest to Patagonian shrubland. J. Veg. Sci. 3:507–520. [ Links ]
VEBLEN, TT & DC LORENTZ. 1987. Post-fire stand development of Austrocedrus–Nothofagus forests in Patagonia. Vegetatio 71:93–111. [ Links ]
WHITMORE, TC. 1989. Canopy and the two major tree forests groups. Ecology 70:536–538. [ Links ]

