










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.11 no.2 Córdoba July/Dec. 2001
Alimentación de la carpa (Cyprinus carpio Linnaeus 1758) en la laguna de Lobos, Provincia de Buenos Aires, Argentina
Darío C Colautti* & Mauricio Remes Lenicov
Instituto de Limnología "Dr. Raúl A. Ringuelet", UNLP-CONICET, La Plata, Buenos Aires, Argentina
* Inst. de Limnología "Dr. Raúl A. Ringuelet" UNLP-CONICET; C.C. 712; 1900 La Plata; Buenos Aires, Argentina. Email: dariocolautti@infovia.com.ar
Recibido: 6 junio 2001;
Revisado: 17 julio 2001;
Versión final: 7 septiembre 2001;
Aceptado: 7 septiembre 2001
RESUMEN. La carpa (Cyprinus carpio) es un pez exótico que actualmente habita la mayoría de las lagunas pampásicas. El conocimiento del espectro trófico y de la importancia relativa de los componentes de su dieta representa información muy útil para la comprensión de su biología y del nicho ecológico ocupado por sus poblaciones en estos ambientes. En este trabajo se estudió la alimentación de la carpa en la laguna de Lobos, a partir del análisis de 75 tubos digestivos de peces con una longitud estándar promedio de 409.7 mm, capturados entre marzo y agosto de 1994. Los ítems alimentarios fueron contados y clasificados taxonómicamente hasta el nivel de familia. Con la información obtenida se calcularon índices alimentarios y se analizó el patrón de variación estacional de los contenidos digestivos empleando un Análisis de Correspondencia. La especie presentó un espectro trófico amplio. Los principales ítems alimentarios, en orden de importancia, fueron restos vegetales, Ciclopoidea, Chidoridae, Chironomidae, Bosminidae, Harpacticoidae y semillas. El segundo eje del ordenamiento indicó la existencia de estacionalidad en la dieta, que estaría relacionada con la disponibilidad de los organismos en el ambiente. La abundancia de organismos zooplanctónicos en la dieta coincide con lo hallado en ambientes de escasa profundidad de Asia y Europa, y estableció la mayor diferencia con respecto a varios lagos profundos del mundo donde la carpa es netamente bentófaga. Esto se debería al carácter oportunista y a la plasticidad trófica de la especie, la cual aprovecharía la oferta de un recurso alimentario particularmente abundante en las lagunas pampásicas.
ABSTRACT. Feeding of carp (Cyprinus carpio Linnaeus 1758) in Lobos shallow lake, Buenos Aires Province, Argentina: The carp (Cyprinus carpio) is an exotic fish that currently inhabits most of the Pampasic shallow lakes. A better knowledge of its trophic spectrum and of the relative composition of its diet may represent a very useful information to understand its biology, as well as the ecological niche occupied by its populations in these environments. In this work the food habits of the carp were studied in Lobos shallow lake, from the analysis of 75 digestive tracts of fish with a mean standard length of 409.7 mm, captured between March and August of 1994. Food items were counted and classified taxonomically up to the family level. Values for several indexes were calculated with this information, and the seasonal pattern of variation of the digestive contents was analyzed using Correspondence Analysis. The species showed a wide trophic spectrum. The main food items, in order of importance were vegetable fragments, Ciclopoidea, Chidoridae, Chironomidae, Bosminidae, Harpacticoidae and seeds. The second axis of the ordination showed the existence of seasonality in the diet, which could be related to the availability of organisms in the environment. The abundance of zooplanktonic organisms in the diet coincides with that found in fishes from Asian and European shallow lakes, and it established the major difference with several deep lakes of the world, where the carp is exclusively bottom-feeder. This could be due to the opportunistic character and to the trophic plasticity of this species, which would take advantage of the offer of a particularly abundant food resource in the Pampasic shallow lakes.
INTRODUCCIÓN
La carpa, Cyprinus carpio, es un pez de origen asiático que actualmente posee distribución cosmopolita (Crivelli 1981; Karam et al. 1986). Su introducción en Argentina ocurrió hacia fines del siglo XIX, aunque recién en la década de 1940 se registró en estado salvaje en el Río de la Plata (Mac Donagh 1945). Fue catalogada como potencial invasora de los ambientes continentales pampásicos (Ringuelet et al. 1967), pero su presencia en aguas interiores de la provincia de Buenos Aires no fue comunicada hasta 1987 (Barla & Iriart 1987). Durante esa década, debido a inundaciones extraordinarias, se propagó colonizando diversos limnótopos, incrementando rápidamente su área de distribución (Candia 1991; López et al. 1991, 1996; Fabiano et al. 1992; Miquelarena & López 1995; Almiron et al. 1997) y la biomasa de sus poblaciones (Colautti 1997).
Los efectos de la carpa sobre los ambientes acuáticos que ha invadido fueron circunscriptos a las relaciones observadas entre su abundancia y la destrucción de la vegetación acuática (Robel 1962; McCrimmon 1968; McCrimmon & Skobe 1970; Crivelli 1983; Fletcher et al. 1985), el aumento de la biomasa de algas (Brönmark & Weisner 1992) y el incremento de la turbidez (Skubinna et al. 1995; Sidorkewicj et al. 1996). Otros efectos de las carpas sobre el ambiente son especulativos (Powles et al. 1983) y hasta contradictorios. Si bien se le atribuye un impacto sobre cambios en la comunidad zooplanctónica (Kirk & Gilbert 1990; Kirk 1991), los estudios al respecto no han sido concluyentes (Lougheed et al. 1998).
Las introducciones exitosas de especies producen un importante impacto, alterando la dinámica ecológica a través de modificaciones en la estructura trófica (Granado & García Novo 1981). El estudio e identificación de cuáles son los recursos y cómo son explotados proveen importantes herramientas para la comprensión, conservación, manejo o gestión de los ecosistemas (Ringuelet 1972). El estudio de la dieta de los peces provee aportes importantes a la comprensión de su autoecología y del rol ecológico de sus poblaciones. La identificación de los componentes de la dieta y su importancia relativa permite esclarecer el origen de los recursos que las sostienen (Windell & Bowen 1978). La dieta de la carpa ha sido ampliamente estudiada en diversos países del mundo (e.g., Uribe Zamora 1975; Guziur 1976; Matlak & Matlak 1976; Crivelli 1981). En este trabajo se analiza cualitativamente y cuantitativamente la dieta de la carpa en una laguna oligohalina típica de la cuenca del río Salado, con el fin de conocer el nicho trófico que ha ocupado esta especie invasora.
ÁREA DE ESTUDIO
La laguna de Lobos (35°16'S; 59°07'O) es un cuerpo de agua de llanura que forma parte del conjunto de lagunas oligohalinas típicas de la cuenca media y baja del río Salado de la provincia de Buenos Aires (Ringuelet 1972). Se encuentra a una altura de 21.12 m.s.n.m., tiene una superficie promedio de 750 ha y su principal afluente es el arroyo Las Garzas, que drena un área de 1720 km2 (Dangaus et al. 1990). Posee un único efluente, el arroyo Saladillo, en el cual se ha construido un dique regulador. Al nivel del vertedero, la laguna tiene una profundidad promedio de 1.15 m y máxima de 1.48 m. Hidrológicamente es inestable, debido a que el volumen de agua contenido está sujeto al manejo de la compuerta y a los eventos hídricos de la cuenca. Las precipitaciones son un factor de suma importancia, ya que producen los cambios ecológicos más notables (Boltovskoy et al. 1990) y determinan que se conecte con el resto de la red de aguas superficiales. La fauna íctica de la laguna está compuesta por 22 especies (López 1987), siendo mayor la abundancia relativa de la fracción de peces asociados al bentos y a zonas vegetadas (Freyre et al. 1997). La dinámica anual de la comunidad planctónica y algunos parámetros limnológicos fueron descriptos por Boltovskoy et al. (1990), mientras que Rodrigues Capítulo et al. (1995) hicieron lo propio con el bentos. El 16% de la superficie de la laguna se encuentra cubierta por juncos (Schaenoplectus californicus), distribuidos en parches a lo largo de sus orillas; durante este estudio no se registró la presencia de macrófitas sumergidas ni flotantes.
MÉTODOS
En el otoño y el invierno de 1994 se efectuaron muestreos mensuales entre marzo y agosto. Durante este periodo fueron capturadas 75 carpas con red de tiro costero. Los ejemplares se midieron, se pesaron y se les extrajo el tracto digestivo completo, que fue fijado en formol al 10%. Mediante Análisis de Varianza se evaluó la existencia de diferencias entre las tallas de las carpas capturadas en cada fecha.
En el laboratorio se vació el contenido del tercio anterior del tracto digestivo, que se lavó en un tamiz de 30 µ. El material retenido fue diluido en un volumen de 50 ml de agua y fue sometido a agitación magnética para extracción de submuestras (Starostka & Applegate 1970; Engel 1976), obteniéndose tres de 5 ml cada una. Once muestras de las 75 obtenidas fueron descartadas debido a que la parte anterior de los tubos digestivos sólo contenía mucus. La identificación y conteo de los componentes se realizó bajo lupa binocular; losítems de origen animal fueron clasificados taxonómicamente hasta el nivel de familia. El volumen de los ítems alimentarios que se presentaron como unidades discretas fue estimado de acuerdo a las ecuaciones sugeridas por Ringuelet et al. (1980) y Escalante (1982). La representación en cada submuestra de los restos vegetales (fragmentos vegetales de origen acuático y terrestre en diferente grado de descomposición) fue registrada con un valor porcentual y su volumen se estimó en forma indirecta por diferencia con el resto de los componentes. Finalmente, se expresó en forma de porcentaje el volumen ocupado por cada ítem para el total de las muestras (%Vol).
Tomando en consideración los estudios sobre los componentes de las comunidades de los ambientes pampásicos y su relación con la dieta de los peces (Destefanis & Freyre 1972; Boltovskoy et al. 1990; Rodrigues Capítulo et al. 1995; Grosman et al. 1996), se elaboró una tabla en la cual se asignó cada ítem alimentario de la carpa en esta laguna a una comunidad.
Para caracterizar el espectro trófico y la constancia de aparición de los ítems entre los componentes de la dieta se aplicaron el índice de presencia (IO), que es el cociente (multiplicado por 100) entre el número de contenidos en el que apareció determinado ítem y el número total de contenidos analizados (Hyslop 1980), y el índice de composición porcentual por número (CPN), que es el cociente (multiplicado por 100) entre el número de presencias de cierto ítem y el número total de presencias registrado (Crisp et al. 1978; Hyslop 1980). Con el objeto de comprobar si existía relación entre ambos índices, se transformaron los valores de IO y CPN a sus respectivos logaritmos y se estimó el Coeficiente de Correlación de Pearson. Finalmente, se obtuvo el índice de importancia relativa (IRI) (Pinkas et al. 1971), que relaciona los valores de losíndices calculados previamente entre sí y diagnostica la importancia de cada ítem en la dieta. Los valores se obtuvieron utilizando la siguiente fórmula:
IRI = IO (CPN + %Vol) .
Para los restos vegetales detríticos y los sedimentos no fue posible obtener valores de CPN e IRI, debido a que no se presentaron en forma discreta. Por lo tanto, la importancia de estos ítems en la dieta solo pudo ser evaluada a partir del IO y del volumen porcentual.
La composición de la dieta y su posible relación con la estacionalidad fue explorada mediante un Análisis de Correspondencia (Pielou 1984) sobre la matriz de 64 individuos por los 15 ítems de mayor importancia relativa de acuerdo al IRI. Como datos básicos se utilizaron los logaritmos de las abundancias relativas de los ítems (log (x + 1)). Esta transformación se efectuó con el objeto de suavizar las diferencias entre los promedios de los distintos ítems, de manera que el ordenamiento estuviera más influido por cambios en la composición de la dieta. Los posibles cambios alimentarios en relación a las fechas de muestreo fueron analizados mediante la estimación del Coeficiente de Correlación por Rangos de Spearman (rs) entre los días julianos correspondientes a la toma de muestra de cada contenido y el valor de los mismos en cada una de las dos primeras dimensiones del ordenamiento.
RESULTADOS
Los estadísticos referentes a las tallas de los individuos utilizados en el estudio, discriminados por fechas, se presentan en la Tabla 1. El Análisis de Varianza no detectó diferencias significativas entre las longitudes de los grupos de ejemplares correspondientes a cada fecha (F = 0.14, P = 0.97).
Tabla 1. Cantidad y tamaño de las carpas capturadas en la laguna de Lobos en 1994. Se muestran la fecha de muestreo, el número de ejemplares analizados (n), la longitud estándar promedio (± DE, con el rango entre paréntesis) y el número de ejemplares involucrados en el análisis de la dieta (nd). Todas las medidas están en mm.
Table 1. Number and size of carps captured in Lobos shallow lake during 1994. Sampling dates, number of captured individuals (n), mean standard length (± SD, with range between brackets), and number of individuals included in diet analysis (nd) are shown. All measures are in mm.

Los elementos hallados en los tractos digestivos fueron agrupados en 27 ítems, de los cuales la mayoría corresponde a la comunidad bentónica (16) y planctónica (6). También se hallaron elementos procedentes de la comunidad terrestre y del perifiton (Tabla 2). La presencia de alimentos de origen animal y vegetal indicaron que la especie posee una dieta omnívora y un espectro trófico amplio. Los resultados del IO, del CPN, del volumen porcentual y del IRI para cada uno de los ítems se consignan en la Tabla 2. Los copépodos Ciclopoidea y los restos vegetales detríticos fueron los alimentos más recurrentes, habiéndolos hallado en casi la totalidad de los contenidos. No obstante, los Chironomidae, los cladóceros Chidoridae y Bosminidae, los copépodos Harpacticoidea y las semillas fueron explotados con frecuencia similar y se encontraron en más del 50% de los individuos. Los logaritmos del IO y del CPN mostraron una correlación positiva entre sí (r = 0.76, P < 0.05). En el análisis volumétrico se destacó la importancia de los ítems restos vegetales, Chironomidae, Ciclopoidea, Palaemonidae, huevo de pez y semillas (Tabla 2). Sin embargo, algunos de éstos ocuparon gran volumen por ser los ítems de mayor tamaño y no resultaron relevantes al evaluar su importancia numérica o su presencia (e.g., Palaemonidae y huevo de pez).
Tabla 2. Ítems alimentarios encontrados en los contenidos digestivos de las carpas capturadas en la laguna de Lobos en 1994. Se muestran los valores del índice de presencia (IO), del índice de composición porcentual por número (CPN), del volumen porcentual (%Vol) y del índice de importancia relativa (IRI) para cada ítem alimentario. La comunidad a la que se asignó cada uno de los ítems está basada en la bibliografía consultada.
Table 2. Food items found in digestive tracts of carps captured in Lobos shallow lake during 1994. Values for the occurrence index (IO), for the percent composition per number index (CPN), for percent volume (%Vol), and for the relative importance index (IRI) are shown for each item. Each item was assigned to a community on the basis of literature data.
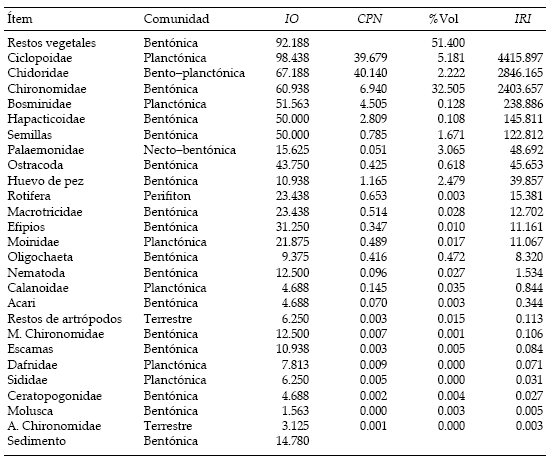
El IRI permitió jerarquizar en orden de importancia relativa los ítems alimentarios de la carpa en la laguna de Lobos, y su distribución permitió discriminar tres grupos. El primero, comprendido por Ciclopoidea, Chidoridae y Chironomidae, presentó la mayor importancia relativa debido a que sus valores de IRI se ubicaron uno o dos órdenes de magnitud por encima del resto de los ítems. No obstante, los restos vegetales deben considerarse parte de este grupo, porque presentaron el máximo valor de volumen (más del 50%) y el segundo de IO; y, teniendo en cuenta la correlación entre IO y CPN, es de esperar que también hubiesen alcanzado un valor elevado para este último índice y, consecuentemente, también el mayor IRI. El segundo grupo quedó formado por los ítems Bosminidae, Harparticoidae, semillas, Palaemonidae y Ostracoda, con relevancia secundaria en la dieta. Un último conjunto estaría conformado por el resto de los ítems, los cuales serían utilizados ocasionalmente. Entre ellos merecen destacarse huevos de pez, Rotifera, Macroticidae, Efipios, Moinidae y Oligocheta.
El primer eje del Análisis de Correspondencia retuvo el 22.2% (autovalor = 0.32) de la inercia de la matriz de individuos por ítems alimentarios (Figura 1), indicando que la mayor diferencia en cuanto a composición del contenido digestivo estuvo dada entre las carpas que consumieron Oligochaeta, huevos de pez y Palaemonidae y las que presentaron Rotifera, Bosminidae, Efipios y Chidoridae. Esas diferencias no parecieron estar asociadas a la fecha de captura (rs= 0.24, P = 0.04). El segundo eje (autovalor = 0.22) representó el 15.2% de la variación, indicando diferencias entre los contenidos que tenían Rotifera, Palaemonidae, Oligochaeta y Bosminidae y los que tenían Harpacticoidae, Chidoridae, Macrotriciade y Efipios. Estas diferencias estuvieron asociada a la fecha (rs= 0.66, P < 0.001). Las carpas capturadas en abril y marzo tendieron a aparecer en el cuadrante inferior del ordenamiento, mientras que las capturadas en julio y agosto tendieron a estar en el cuadrante superior (Figura 1). Los restos vegetales, ciclopoideos, semillas y Ostracoda fueron comunes en la mayoría de las carpas y por eso se ubicaron en el centro del ordenamiento.

Figura 1. Ubicación de las muestras del contenido digestivo discriminadas por mes (símbolos) y de los ítems alimentarios (letras) en el plano definido por las dos primeras dimensiones del Análisis de Correspondencia realizados con los datos de dieta de las carpas capturadas en la laguna de Lobos en 1994. A: Chidoridae, B: Bosminidae, C: Moinidae, D: Macrotricidae, E: Ciclopoidae, F: Hapacticoidae, G: Chironomidae, H: Ostracoda, I: Palaemonidae, J: Efipios, K: Oligochaeta, L: Rotifera, M: semillas, N: huevos de pez, P: restos vegetales.
Figure 1. Distribution of samples of digestive tracts, according to month (symbols), and of food items (letters), in the plane defined by the first dimensions obtained in the Correspondence Analysis of diet data for carps captured in Lobos shallow lake during 1994. See letter codes in the legend above.
DISCUSIÓN
A partir de los resultados obtenidos pueden realizarse algunas generalizaciones acerca de los constituyentes de la dieta de la carpa y de la manera en que la especie los utiliza durante el otoño y el invierno en la laguna de Lobos. Los restos vegetales, Ciclopoidea, Chidoridae y Chironomidae aparecieron con alta frecuencia, en número y volumen elevados; por lo tanto, serían los recursos tróficos más importantes para la carpa. Los Bosminidae, Harpacticoidea, las semillas, Palaemonidae y Ostracoda resultarían alimentos secundarios, mientras que el resto de los ítems representaron alimentos ocasionales. La alta correlación existente entre el IO y el CPN indica que los individuos muestreados explotaron los recursos de manera similar y que los alimentos numéricamente importantes también fueron consumidos con mayor asiduidad. Teniendo en cuenta los componentes de la dieta y su ubicación en el ambiente, puede afirmarse que la carpa se alimenta básicamente del bentos, ya que la mayor parte de los elementos hallados pertenecen a dicha comunidad. Entre los 27 ítems identificados, las presas zooplanctónicas fueron siete; sin embargo, su importancia relativa en la dieta resultó elevada. Esto estaría vinculado a la abundancia de estos organismos y a la mezcla parcial existente entre los componentes de las comunidades bentónica y planctónica en las lagunas pampásicas (Rodrigues Capítulo et al. 1995).
El patrón de variación estacional de la composición cuali–cuantitativa de los contenidos se manifestó con un predominio de Bosminidae, Rotifera, Palaemonidae y Moinidae en los contenidos digestivos de marzo y abril, mientras que las semillas, Ostracoda y Macrotricidae estuvieron mejor representados en los contenidos de junio, al final del otoño. El consumo de retos vegetales y Ciclopoidae se mantuvo constante, mientras que la ingesta de Chironomidae experimentó un leve incremento hacia fines del invierno. Durante esta estación se acentuó la dominancia de los Chidoridae y Efipios en julio, y de Harparticoidae, Oligochaeta y huevos de pez hacia agosto. Estas variaciones de la dieta estarían influenciadas por la disponibilidad de los ítems en el ambiente. Boltovskoy et al. (1990) y Rodrigues Capítulo et al. (1995) resaltan las fluctuaciones espacio–temporales de la abundancia de los organismos planctónicos y bentónicos en la laguna de Lobos, y concuerdan en que los Ciclopoidae constituyen el ítem más constante en presencia y abundancia. Por su parte, los Bosminidae y los Rotifera dominan a principios del otoño, mientras que los Efipios dominan durante el invierno y, finalmente, los Harpacticoidae y los Chironomidae incrementan su abundancia hacia la primavera. Esto coincide con el patrón de cambio estacional en la dieta de la carpa, explica la variación intra-estacional y apoya la hipótesis del consumo basado en la disponibilidad, tal cual se plantea en la teoría del consumo óptimo (Pyke 1984). Dado que la laguna de Lobos es un limnótopo oligohalino típico de la cuenca media y baja del río Salado, es muy probable que durante otoño e invierno la alimentación de la especie no difiera demasiado con respecto a otros cuerpos de agua de dicha región.
Según Sibbing (1982, 1988, 1991), Sibbing & Uribe (1985) y Sibbing et al. (1986), la carpa posee una serie de especializaciones vinculadas a mecanismos eficientes de extracción, purificación, retención selectiva, trituración, deglución y asimilación del alimento. De acuerdo a estos autores, estas particularidades restringen su acceso al fitoplancton y limitan la utilización eficiente de los restos vegetales y de organismos con mayor tamaño y movilidad (macroinvertebrados y peces). A pesar de lo mencionado, los restos vegetales conforman un ítem que aparece con alta frecuencia y máximo volumen en los contenidos revisados. La abundancia de los mismos concuerda con la revisión efectuada por Powles et al. (1983), quienes señalan que el volumen ocupado por este ítem varía entre 19 y 98%. Sin embargo, el valor alimenticio de los restos vegetales es incierto, debido a que la carpa es incapaz de romper mecánicamente las paredes celulares (Sibbing 1988) y no posee celulasas en el tubo digestivo (Summerfelt et al. 1970). Según Sibbing (1991) la tendencia al herbivorismo sería consecuencia de la escasez de presas animales. Por otra parte, Eder & Carlson (1977) sostienen que la ingestión de restos vegetales se relaciona con la búsqueda de invertebrados. En Lobos, el consumo de rotíferos de talla promedio de 80 µ (IO = 23.4) se debería a la estrecha relación de éstos con los restos vegetales, ya que la abertura mínima del tamiz branquial de una carpa adulta retiene partículas mayores a 250 µ. Por lo tanto, su presencia en los contenidos puede ser considerada accidental. El consumo de huevos de peces, principalmente de pejerrey (Odontesthes bonariensis), siempre estuvo vinculado a los restos vegetales. Su aparición en los contenidos (IO = 10.9) ocurrió hacia fines del invierno (25 agosto 1994). Una observación interesante es que en el 90% de los casos se trató de huevos vacíos que se encontraban fijos a la materia vegetal por sus filamentos. Estos resultados coinciden con los de Cole (1905) y Leach (1919), quienes concluyeron que los huevos de peces son constituyentes infrecuentes en la dieta de la carpa. No obstante, no es posible establecer conclusiones definitivas en este aspecto porque los muestreos no abarcaron completamente la época de desove de las especies autóctonas. Las semillas de macrófitas, principalmente de Schaenoplectus californicus, representaron un ítem de origen vegetal importante. La posibilidad de triturarlas con los dientes faríngeos le permite a la carpa aprovechar su alto valor calórico (Sibbing 1988). Los Palaemonidae presentaron un alto porcentaje en el análisis volumétrico, pero los bajos valores de IO y CPN indican que se trata de un recurso poco explotado. Según Drenner et al. (1982), el poder de succión que es capaz de generar la carpa le permite capturar, de una vez, grandes cantidades de crustáceos, incluso aquellos con mayor capacidad de escape. La dominancia de los microcrustáceos con respecto a los Palaemonidae estaría relacionada con la abundancia de los mismos en el ambiente. Esta hipótesis debería ser puesta a prueba con estudios experimentales en los cuales se considere la oferta alimentaria. El sedimento fue hallado en una baja proporción de ejemplares, pero fue abundante en aquellos contenidos con predominancia de organismos ligados al fondo, como Oligochaeta y Chironomidae; su ingestión puede ser considerada accidental.
La revisión bibliográfica realizada sobre la dieta de la carpa en ambientes naturales (McCrimmon 1968; Guziur 1976; Matlak & Matlak 1976; Crivelli 1981; Powles et al. 1983; Panek 1987; Sibbing 1988; Chapman & Fernando 1994) indica que es un pez omnívoro que se alimenta sobre el fondo, fundamentalmente de invertebrados bentónicos. No obstante, se observan diferencias entre el régimen alimentario de las carpas en lagos profundos con estratificación y el de ambientes de aguas someras. En la laguna de Lobos, donde el consumo de invertebrados zooplanctónicos fue elevado, la dieta resultó parecida a la adoptada por la especie en ambientes de escasa profundidad de otros sitios del planeta. Las diferencias cuali–cuantitativas mencionadas estarían relacionadas a las condiciones limnológicas y a la capacidad de la especie para aprovechar los recursos disponibles (Moen 1953; Nakamura 1955; Sigler 1958; Rehder 1959; Walberg & Nelson 1966; Summerfelt et al. 1970; Eder & Carlson 1977; Chapman & Fernando 1994).
Como ya se ha mencionado, la carpa es capaz de producir modificaciones sobre los ecosistemas luego de colonizarlos. Es poco lo que puede decirse con relación a los posibles efectos que podría causar la especie sobre la flora de la laguna, ya que el material vegetal hallado en los contenidos estaba compuesto por fragmentos de plantas acuáticas y terrestres en diferente grado de descomposición. En el largo y mediano plazo, es posible que se detecten efectos indirectos relacionados a la inclusión de esta especie como nuevo elemento en el ecosistema, aunque es difícil establecer pronósticos. Los mismos seguramente estarán vinculados a cambios en la dinámica de nutrientes, la turbidez y la abundancia relativa de las especies animales y vegetales, como ha ocurrido en otras partes del mundo (Lougheed et al. 1998).
Los trabajos sobre alimentación de peces de la pampasia (Destefanis & Freyre 1972; Escalante 1982, 1983a, 1983b, 1984) permiten establecer que la carpa comparte gran parte de sus ítems alimentarios con la mayoría de las especies nativas y que, además, posee algunos exclusivos como las semillas. Aunque buena parte de las especies autóctonas son generalistas, ninguna parece tener un nicho trófico tan amplio como la carpa, que ante la escasez de recursos alimentarios tendría ventajas comparativas. El establecimiento de poblaciones de carpa en las lagunas pampásicas es solo un ejemplo más de la gran variedad de limnótopos que es capaz de colonizar esta especie, que habita aguas continentales desde las altas latitudes de Canadá (McCrimmon 1968) y Finlandia (Ahlfors et al. 1982) hasta el trópico (Pinilla et al. 1992; Ritter Ortiz et al. 1992). La eficiencia para explotar diferentes comunidades y aprovechar los recursos disponibles, recurriendo incluso a los niveles tróficos más bajos (Schroeder 1980), ha sido indudablemente una de las claves de su éxito y de su actual distribución cosmopolita.
AGRADECIMIENTOS
Los autores quieren expresar su agradecimiento a Fabián Grosman por el apoyo brindado en las primeras etapas del trabajo y a Juan Manuel Morales por su valiosa contribución en la elaboración de la versión definitiva del manuscrito. Esta es la contribución 713 del Instituto de Limnología "Dr. Raúl A. Ringuelet".
BIBLIOGRAFÍA
AHLFORS, P; P KUMMU & K WESTMAN. 1982. Introduction of carp Cyprinus carpio in Finland. EIFAC. Symposium on stock enhancement in the management of freshwater fisheries, Budapest. 19 pp. [ Links ]
ALMIRON, A; M AZPELICUETA; J CASCIOTTA & A LÓPEZ CAZORLA. 1997. Ichthyogeographic boundary between the Brazilian and Austral subregions in South America, Argentina. Biogeographica 73:23–30. [ Links ]
BARLA, MJ & R IRIART. 1987. La presencia de la carpa (Cyprinus carpio L.) en la laguna de Chascomús y su significado. Limnobios 2:685–686. [ Links ]
BOLTOVSKOY, A; A DIPPOLITO; M FOGETTA; N GÓMEZ & G ÁLVAREZ. 1990. La laguna de Lobos y su afluente: limnología descriptiva, con especial referencia al plancton. Biología Acuática 14:1–38. [ Links ]
BRÖNMARK, C & SEB WEISNER. 1992. Indirect effects of fish community structure on submerged vegetation in shallow, eutrophic lake: an alternative mechanism. Hydrobiologia 243/244:293–301. [ Links ]
CANDIA, C. 1991. Sobre la distribución de la Carpa Cyprinus carpio en el Río de la Plata y en cuerpos de agua de la Provincia de Buenos Aires. Biología Acuática 15(2):166–167. [ Links ]
CHAPMAN, G & CH FERNANDO. 1994. The diet and related aspects of feeding of Nile tilapia Oreochromis niloticus L. and common carp Cyprinus carpio L. in lowland rice fields in northeast Thailand. Aquaculture 123:281–307. [ Links ]
COLAUTTI, D. 1997. Ecología de la carpa Cyprinus carpio en la cuenca del río Salado, provincia de Buenos Aires. Tesis Doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. 215 pp. [ Links ]
COLE, LJ. 1905. The German Carp in the United States. Reports of the United States Bureau of Fisheries 1904:525–641. [ Links ]
CRISP, DT; RH MANN & JC MCCORMACK. 1978. The effects of impoundment and regulation upon the stomach contents of fish at Cow Gree, Upper Teesdale. J. Fish. Biol. 12:287–301. [ Links ]
CRIVELLI, AJ. 1981. The biology of the carp (Cyprinus carpio L.) in the Camargue, southern France. J. Fish. Biol. 18:271–290. [ Links ]
CRIVELLI, AJ. 1983. The destruction of aquatic vegetation by carp. Hidrobiologia. 106:37–41. [ Links ]
DANGAUS, N; A BLASI; M MORMENEO; M GAILLARD & R BURAKOWSKY. 1990. Estudio geológico de la Laguna de Lobos, Partido de Lobos, Provincia de Buenos Aires. Centro de investigación de suelos y aguas de uso agropecuario (CISAUA), Conv. UNLP-MAAP. [ Links ]
DESTEFANIS, SU & LR FREYRE. 1972. Relaciones tróficas de los peces de la Laguna de Chascomús con un intento de referenciación ecológica y tratamiento bioestadístico del espectro trófico. Acta Zoologica Lilloana 29:17–33. [ Links ]
DRENNER, RW; GL VINYARD; M GOPHEN & S MCCOMAS. 1982. Feeding behaviour of cichlid Sarotherodon galileaeum: selective predation and Lake Kinneret zooplankton. Hidrobiologia 37:17–20. [ Links ]
EDER, S & CA CARLSON. 1977. Food habits of carp and whitesuckers in the South Platte and St. Vrain rivers in Goosequill Pond, Weld County, Colorado. T. Am. Fish. Soc. 106:339–346. [ Links ]
ENGEL, S. 1976. Food habits and prey selection of Coho salmon (Onkorhyncus kisutch) and Cisco (Coregonus artedii) in relation to zooplankton dynamics in Pallette Lake, Wisconsin. T. Am. Fish. Soc. 105:607–608. [ Links ]
ESCALANTE, AH. 1982. Contribución al conocimiento de las relaciones tróficas de los peces de agua dulce del área platense. I. Astianax eigenmanniorum (Osteichthyes Tetragonopteridae). Limnobios 2:311–322. [ Links ]
ESCALANTE, AH. 1983a. Contribución al conocimiento de las relaciones tróficas de los peces de agua dulce del área platense. II. Otros Tetragonopteridae. Limnobios 2:379–402. [ Links ]
ESCALANTE, AH. 1983b. Contribución al conocimiento de las relaciones tróficas de los peces de agua dulce del área platense. III. Otras especies. Limnobios 2:453–463. [ Links ]
ESCALANTE, AH. 1984. Contribución al conocimiento de las relaciones tróficas de peces de agua dulce delárea platense. IV. Dos especies de Cichlidae y miscelanea. Limnobios 2:562–578. [ Links ]
FABIANO, G; F AMESTOY; C GARCÍA & L ARES. 1992. Estudio de las variaciones en la abundancia, la estructura y la distribución espacio–temporal de los efectivos de carpa común (Cyprinus carpio L.) en el Río de la Plata medio e inferior y en el Río Uruguay inferior. Publicaciones de la Comisión Administradora del Río Uruguay. Serie Técnico–Científica 1:13–24. [ Links ]
FLETCHER AR; AK MORISON & DJ HUME. 1985. Effects of Carp (Cyprinus carpio L.) on communities of aquatic vegetation and turbidity of waterbodies in the lower Goulburn Rever Basin. Aust. J. Mar. Fresh. Res. 36:311–327. [ Links ]
FREYRE, LR; ME MAROÑAS & ED SENDRA. 1997. Demografía del pejerrey Odontesthes bonariensis bonariensis de la laguna de Lobos (Provincia de Buenos Aires). Natura Neotropicalis 28:47–59. [ Links ]
GRANADO, C & F GARCÍA NOVO. 1981. Cambios ictiológicos durante las primeras etapas de sucesión en el embalse de Arrocampo (Cuenca del río Tajo, Cáceres). Volumen homenaje al Dr. Lozano Cabo. Bol. Inst. Esp. Oceano. 319(6):224–243. [ Links ]
GROSMAN, FM; JR GONZÁLEZ & EJ USUNOFF. 1996. Trophic niches in an Argentine pond as a way to assess functional relationships between fishes and other communities. Water SA 22(4):345–350. [ Links ]
GUZIUR, J. 1976. The feeding of two years old carp, Cyprinus carpio L., in a vendance Lake Klawoj. Ekologia Polska 24(2):211–235. [ Links ]
HYSLOP, EJ. 1980. Stomach contents analysis, a review of methods and their application. J. Fish. Biol. 17:411–429. [ Links ]
KARAM, M; M TALAAT & J OLAH. 1986. Fishery studies on Cyprinus carpio L. in Hungarian inland waters. Aquacultura Hungarica (Szarvas) 5:241–250. [ Links ]
KIRK, KL. 1991. Inorganic particle alter competition in grazing plankton: the role of selective feeding. Ecology 72:915–923. [ Links ]
KIRK, KL & JJ GILBERT. 1990. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 71:1741–1755. [ Links ]
LEACH, GC. 1919. The artificial propagation of Carp. United States Boreau of Fisheries, Economic Circular 39:1–19. [ Links ]
LÓPEZ, H. 1987. Apuntes ictiológicos de la laguna de Lobos (Pcia. Bs. As.). Bol. Asoc. Arg. Limnol. 5:15–16. [ Links ]
LÓPEZ, H; A RODRIGUEZ CAPÍTULO; J CASCIOTA & J IWASZKIW. 1991. Caracterización limnológica preliminar de la Laguna "El Hinojo", Trenque Lauquen. Situación Ambiental de la Provincia de Buenos Aires 1(1):1–23. [ Links ]
LÓPEZ, HL; LC PROTOGINO & AE AQUINO. 1996. Ictiología continental de la Argentina: Santiago del Estero, Catamarca, Córdoba, San Luis, La Pampa y Buenos Aires. Aquatec. 3:1–14. [ Links ]
LOUGHEED, VL; B CROSBIE & P CHOW-FRASER. 1998. Predictions of the effect of common carp (Cyprinus carpio) exclusion on water quality, zooplankton, and submergent macrophites in Great Lakes wetland. Can. J. Fish. Aquat. Sci. 55:1189–1197. [ Links ]
MAC DONAGH, EJ. 1945. Pesca de una "Carpa de espejuelos" en el Río de la Plata. Not. Mus. La Plata. 89:315–324. [ Links ]
MATLAK, J & O MATLAK. 1976. The natural food of carp fry, Cyprinus carpio L. Acta Hydrobiologia 18:203–228. [ Links ]
MCCRIMMON, HR & E SKOBE. 1970. The fisheries of Lake Simcoe. Ontario Department of Lands and Forests, Toronto. 140 pp. [ Links ]
MCCRIMMON, R. 1968. Carp in Canada. Bull. Fish. Res. Bd. Can. 165:1–93. [ Links ]
MIQUELARENA, AM & HL LÓPEZ. 1995. Fishes of the lagunas Encadenadas (Province of Buenos Aires, Argentina), a wetland of international importance. Freshwater Forum 5(1):48–53. [ Links ]
MOEN, T. 1953. Food habits of the Carp in northwest Iowa lakes. Iowa Academy of Sciences Proceedings 60:655–686. [ Links ]
NAKAMURA, N. 1955. Comparison of the Carp and the Crucian Carp from the piscicultural point of view. Bull. Jap. Soc. Sci. Fish. 21(2):77–81. [ Links ]
PANEK, FM. 1987. Biology and ecology of Carp. Pp. 1–15 en: EL Cooper (ed.). Carp in North America. American Fisheries Society, Bethesda, EEUU. [ Links ]
PIELOU, EC. 1984. The interpretation of ecological data: a primer on classification and ordination. Wiley, New York, EEUU. [ Links ]
PINILLA, GA; P VARGAS & E PATINO. 1992. Aspectos poblacionales de la carpa (Cyprinus carpio L.) en la Laguna Fuquene. Boletin Eutropica 25:28–41. [ Links ]
PINKAS, L; M OLIPHANT & I IVERSON. 1971. Food habits of albacore, bluefin tuna and bonito in Californian waters. Calif. Fish Game 152:1–105. [ Links ]
POWLES, PM; HR MCCRIMMON & DD MACRAE. 1983. Seasonal feeding of Carp, Cyprinus carpio, in the Bay of Quinte watershed, Ontario. Can. Field Nat. 97(3):293–298. [ Links ]
PYKE, GH. 1984. Optimal foranging theory: a critical review. Annu. Rev. Ecol. Syst. 15:523–575. [ Links ]
REHDER, DD. 1959. Some aspects of the life history of the Carp, Cyprinus carpio in the Des Moines River, Boone County. Iowa State Journal of Science 34:11–26. [ Links ]
RINGUELET, RA. 1972. Ecología y bioecología del habitat lagunar o lago de tercer orden de la región neotropical templada (Pampasia sudoriental de la Argentina). Physis 31(81):55–76. [ Links ]
RINGUELET, RA; RH ARAMBURÚ & A ALONSO DE ARAMBURÚ. 1967. Los peces argentinos de agua dulce. Comisión de Investigaciones Científicas, La Plata. 602 pp. [ Links ]
RINGUELET, RA; R IRIART & AH ESCALANTE. 1980. Alimentación del pejerrey (Basilichthys bonariensis bonariensis, Atherinidae) en Laguna de Chascomús, Bs. As., Arg. Relaciones ecológicas de complementación y eficiencia trófica del plancton. Limnobios 1(10):447–460. [ Links ]
RITTER ORTIZ, W; JS SÁNCHEZ & RR RODRÍGUEZ MALDONADO. 1992. Crecimiento, supervivencia y optimización de la carpa (Cyprinus carpio L.) en la presa de Atlangatepec, Tlaxcala. An. Inst. Mar. Limnol. Univ. Auton. México 19(1):43–56. [ Links ]
ROBEL, RJ. 1962. The relationship of carp to waterfowl food plants on a western marsh. Utah Dep. Fish Game Inf. Bull. 62:1–4. [ Links ]
RODRIGUES CAPÍTULO, A; A PAGGI & I CESAR. 1995. Composición del zoobentos de la laguna de Lobos, (Provincia de Buenos Aires Argentina). Limnética 11(1):29–37. [ Links ]
SCHROEDER, GL. 1980. The breakdown of feeding niches in fish ponds under natural conditions of severe competition. Bamidgeh. 32(1):20–24. [ Links ]
SIBBING, FA. 1982. Pharyngeal mastication and food transport in the carp (Cyprinus carpio L.): a cineradiographic and electromyographic study. J. Morph. 172:223–258. [ Links ]
SIBBING, FA. 1988. Specializations and limitations in the utilization of food resources by the carp, Cyprinus carpio: a study of oral food processing. Environ. Biol. Fishes 22(3):161–178. [ Links ]
SIBBING, FA. 1991. Food capture and oral processing. Pp. 377–412 en: IJ Winfield & JS Nelson (eds). Cyprinid fishes: systematics, biology and exploitation. Chapman & Hall, London, Reino Unido. [ Links ]
SIBBING, FA; JW MOSSE & A TERLOUW. 1986. Food handling in the carp (Cyprinus carpio L.), its movement pattern, mechanisms and limitations. J. Zool. (London) 210:160–203. [ Links ]
SIBBING, FA. & R URIBE. 1985. Regional specializations of the oropharyngeal wall and food processing in the carp (Cyprinus carpio L.). Neth. J. Zool. 35:377–422. [ Links ]
SIDORKEWICJ, NS; AC LÓPEZ CAZORLA & OA FERNÁNDEZ. 1996. The interaction between Cyprinus carpio L. and Potamogeton pectinatus L. under aquarium conditions. Hydrobiologia 340:271–275. [ Links ]
SIGLER, WF. 1958. The ecology and use of Carp in Utah. Utah University Bulletin 405:1–63. [ Links ]
SKUBINNA, JP; TG COON & TR BATTERSON. 1995. Increased abundance and depth of submersed macrophytes in response to decreased turbidity in Saginaw Bay, Lake Huron. J. Great Lakes Res. 21:476–488. [ Links ]
STAROSTKA, VJ & R APPLEGATE. 1970. Food selectivity of bigmouth buffalo Ictiobus cyprinellus in Lake Poinsett, South Dakota. T. Am. Fish. Soc. 99:571–573. [ Links ]
SUMMERFELT, RC; PE MAUCK & G MENSINGER. 1970. Food habits of the carp Cyprinus carpio L. in five Oklahoma reservoirs. Proceedings of the Annual Conference, Southeast Association of the Fish and Game Commission 24:352–377. [ Links ]
URIBE ZAMORA, M. 1975. Selection des proies par le filtre branchial de la carpe miroir, Cyprinus carpio. Tesis doctoral. Universite Claude Bernard, Lyon. 127 pp. [ Links ]
WALBERG, CH & WR NELSON. 1966. Carp, river carpsucker, smallmouth buffalo, and bigmouth buffalo in Lewis and Clark Lake, Missouri River. United States Fish and Wildlife Service Research Report 69:1–30. [ Links ]
WINDELL, JT & SH BOWEN. 1978. Methods for study of fish diets based on analysis of stomach contents. Pp. 219–226 en: T Bagenal (ed.). Methods for assessment of fish production in fresh waters. 3ra edn. Blackwell Scientific Publications, Oxford. [ Links ]

